


Abstract
Background/Aim: This study investigated a novel combined therapy of rosmarinic acid (RA)/blue light on head and neck squamous cell carcinoma (HNSCC) cell proliferation in vitro. Materials and Methods: HNSCC cells were exposed to BL (500 mW/cm2) for 90 s, and incubated with 80 μg/ml RA for 1 hour. Cell viability was determined after 24 h using WST-1 assay. Western blot was used to detect treatment-induced changes in epidermal growth factor receptor (EGFR) activation. Hydrogen peroxide (H2O2) and nitric oxide levels were quantified using CM-H2DCFH-DA assays. Apoptosis was assessed using Annexin V/PI staining and flow cytometry. Results: RA/blue light treatment resulted in a significant reduction in cell viability, EGFR activation and H2O2 levels in all HNSCC cell lines. However, no significant changes in NO production or apoptosis induction were found. Conclusion: RA/blue light effectively decreased HNSCC cell proliferation through reduction in EGFR activation and H2O2 production, and not via induction of apoptosis.
Head and neck squamous cell carcinoma (HNSCC) is the sixth most common type of cancer worldwide, which constitutes approximately 90% of all head and neck cancers (1). HNSCC tumors tend to have an aggressive nature, leading to a 20 times higher risk of metastasis (2). Historically, the treatment of HNSCC has been surgery and radiotherapy, and in the last several years the integration of surgery, radiotherapy and chemotherapy has become the standard of care (3). Many of the drug treatments required at late stages are intensely toxic, and can cause permanently debilitating side effects or even death (4). For instance, administration of cisplatin can cause permanent damage to renal, gastrointestinal, and nervous systems (4, 5). Current research has begun to shift towards naturally occurring compounds in order to combat these tremendous side effects of cancer medications (6). Many of these compounds have been used in traditional medicines for centuries with very few adverse reactions (7). Furthermore, several studies have shown that there is a lower risk of various cancers developing when plant-derived foods are incorporated into the diet, suggesting that such natural products may have chemopreventative properties (6).
Rosmarinic acid (RA) is a naturally occurring polyphenol compound found in plant species Lamiaceae, which includes the common herbs basil, rosemary, sage, and mint (6). There are multiple biological activities associated with RA, which include anti-inflammatory and antioxidant properties as well as neuro- and hepatoprotective activities (8, 9). Such biological activities may render RA to also have an anti-tumorigenic as well as a chemopreventative role. Multiple studies have been shown that RA has the ability to hinder cancer growth within organs throughout the body (10). One study has revealed that rosemary extract, administered through enriched foods, decreased skin tumor numbers in patients with advanced breast cancer (10, 11), suggesting that it reduces metastatic potential. Osakabe et al. have previously shown that pre-treatment with RA reduces reactive oxygen species (ROS) production in mice treated with 12-tetradecanoylphorbol 13-acetate (TPA), which may contribute to its anticarcinogenic property via scavenging of reactive oxygen radicals (6).
It has long been reported that cancers thrive in an oxidative environment (12), making redox status a target for chemotherapeutic agents. Chemotherapeutic drugs have been shown to either increase oxidative stress to toxic levels or to inhibit ROS altogether, which may lead to a decrease in cancer cell proliferation (13). The relationship between hydrogen peroxide and pro-proliferative signaling pathways is well established and largely is a consequence of EGFR-Nox signaling (14). Epidermal growth factor receptor (EGFR) is a highly expressed receptor tyrosine kinase found in a wide variety of tumor cell types, and often is overexpressed in HNSCC (15). Binding of the EGF ligand to the surface receptor leads to downstream activation of proteins that regulate cell proliferation and metastasis (16). Thus, EGFR is considered a key regulatory protein involved in cancer cell survival. Tumur et al. have shown that RA pre-treatment significantly reduces EGFR activation in UM-SCC-6 cells (17), providing a rationale for RA's anti-proliferative effects. Furthermore, RA treatment significantly reduced cell migration, suggesting that RA may reduce tumor metastasis (17).
Blue light (BL) (400-500 nm) is a commonly used tool in dentistry to catalyze the polymerization of resin composites (18). Most light research has focused on the effects of UV light on tissue damage. However, the effects of quartz-tungsten-halogen (QTH) BL on biological systems have not been extensively studied. Lewis et al. have previously shown that BL (400-500 nm) exhibits a level of selective toxicity, causing significant reduction in oral cancer cell viability, while having no adverse effects on normal human epidermal keratinocytes (NHEK) (19). BL treatment was also effective in significantly slowing tumor growth in vivo (20). Thus, BL may be beneficial for the treatment of peripheral cancers with little or no effect on normal healthy cells.
To the best of our knowledge, for the first time, this study investigates whether a combination therapy treatment of RA/BL would have a synergistic inhibitory effect on HNSCC cell proliferation. Furthermore, EGFR, a key regulatory protein in head and neck squamous carcinoma cell proliferation was studied to determine if EGFR activation can be influenced in response to these therapies. Finally, oxidative and nitrosative stress (H2O2/NO) was assessed, as well as treatment-induced apoptosis.
Materials and Methods
Reagents. RA was purchased from Sigma-Aldrich (St. Louis, MO, USA). A concentration of 80 μg/ml was used for all experiments. BL was delivered from a QTH dental curing light (500 mW/cm2; VIP, Bisco, Schaumburg, IL, USA). Fiberoptic tips (6.4 mm diameter) were used to uniformly treat cell cultures in a 24- or 96-well format at a distance of 7.5 mm from the medium surface. All antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA).
Cell culture. HNSCC cell lines (UM-SCC-1, UM-SCC-6 and OSC-2) were grown in Dulbecco's modified Eagle's medium/F-12 (DMEM/F-12, Thermo Scientific, Rockford, IL, USA) containing 2.5 mM L- glutamine, 15 mM HEPES buffer, 10% fetal bovine serum, 100 μg/ml penicillin, and 100 μg/ml streptomycin. Human oral keratinocytes (HOK) cells were grown in OKM Growth Medium containing OKGS Supplement (Sciencell, Carlsbad, CA, USA). Cells were trypsinized at 70% confluency and seeded into 96- or 24-well plates at densities of 5×103 or 5×104 cells/well, respectively. After seeding, the cells were allowed to attach overnight at 37°C with 5% CO2. Cells were then starved for 2 h with serum-free DMEM/F12 plus 100 μg/ml penicillin and 100 μg/ml streptomycin prior to specified wells being treated with BL (400-500 nm) (60 J/cm2, 2 min) and incubated at 37°C with 5% CO2 for 1 h. RA (80 μg/ml in serum-free DMEM/F-12) was added to specified wells (RA and combination treatment) and incubated for the specified time period. For EGFR activation experiments, EGF (100 μg/ml; Sigma-Aldrich) was added 1 h after BL or combination treatment and cells were harvested 5 min later for protein extraction and western blot analysis.
Cell proliferation. Cell proliferation was assessed using the WST-1 Cell Proliferation assay kit (Roche, Indianapolis, IN, USA). UM-SCC-6 cells were treated as described above. At 24 h, WST-1 solution (10 μl/well) was added and cells were incubated at 37°C. Absorbance was measured using a microplate reader at 460 nm at 1, 2, and 3 h time points.
Western blot analysis. Cells were washed twice with ice cold PBS, then scraped into RIPA buffer (Thermo Scientific) containing phosphatase and protease inhibitor cocktails (Halt™; Thermo Scientific), transferred to microfuge tubes, and incubated on ice for 5 min. Cell membranes were removed by centrifugation for 20 min at 4oC at 12000 rpm and protein concentration of the supernatant was determined using the Coomassie Bradford protein assay (Thermo Scientific). Proteins (25 μg) were separated by electrophoresis on 8% SDS-PAGE gels and transferred to PVDF membranes (BioRad, Hercules, CA, USA). Total EGFR and activated EGFR were detected with primary antibodies specific for EGFR (D38B1; 1:500) and phospho-EGFR (Y992; 1:500). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a loading control and detected with anti-GAPDH (14C10; 1:2500). Secondary antibodies conjugated to IRDye® (1:10000; LI-COR, Lincoln, NE, USA) were used to detect protein-primary antibody complexes. Membranes were scanned on the LI-COR Odyssey imaging system and band densities were calculated using ImageJ software.
Hydrogen peroxide assay. Hydrogen peroxide was measured at 24 h post-treatment in untreated control, BL-treated, RA-treated, and combination treated UM-SCC-6 cells using the Amplex® Red Hydrogen Peroxide Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) as described by the manufacturer's instructions. The results were read on a microplate reader with an emission of 590 nm and excitation of 560 nm.
Nitric oxide assay. Nitric oxide levels were assessed in UM-SCC-6 cells immediately after treatment using 4-amino-5-methylamino-2’,7’-difluorofluorescein diacetate (DAF-FM diacetate) reagent (5 μM; Molecular Probes Life Technologies, Eugene, OR, USA). The DAF-FM diacetate was added immediately following treatment and the cells were incubated for 20 min at 37°C. The cells were washed twice with 1X PBS, incubated in 1X Dulbecco's PBS for 15 min, and then fluorescence was read on a microplate reader with excitation at 495 nm and emission at 515 nm.
Flow cytometry. UM-SCC-6 cells were seeded at a density of 50,000 cells in 1 ml DMEM/F-12 in 24-well plates and allowed to attach overnight. The cells were then treated with BL, RA and combination, and incubated again for a 12-h period. For positive control, 1 μM staurosporine (STS; ApexBio, Taiwan) was used to induce apoptosis. At 12 h, the cells were harvested and prepared for flow cytometry using the Annexin V Apoptosis Detection Kit (Thermo Fisher Scientific) following the manufacturer's instructions. In the Annexin V vs. propidium iodide (PI) plot, the FL4-A filter was used for Annexin V and FL2-A filter was used for PI.

WST-1 cell proliferation assay of various HNSCC cell lines: UM-SCC-6, OSC-2, UM-SCC-1 and normal HOK cells 24 h post-treatment with RA, BL, and RA+BL. Data are expressed as % control (set at 100%) ±SD.
Statistical analysis. Data were expressed as mean±SD. One-way ANOVA test was performed to compare the values between the groups. Results were considered statistically significant when p-values were less than 0.05.
Results
RA, BL and combination treatments reduced the growth rate of HNSCC cell lines. The effects of RA and BL alone and in combination were tested on three HNSCC cell lines - UM-SCC-1, UM-SCC-6, and OSC-2 – and compared to their effects on normal human oral keratinocytes (HOK). All HNSCC cell lines tested showed significant reductions in cell proliferation 24 h following each single treatment of RA (47-64% reduction) or BL (61-74% reduction) when compared to untreated controls (Figure 1). In addition, in each case the combination treatment was significantly more effective (82-89% reduction) than either single therapy alone. However, there was a much less dramatic effect on HOK, with RA or BL single treatment reducing proliferation by 34% and 16%, respectively. In addition, there was no additive or synergistic effect of the dual therapy, suggesting a reduced sensitivity to these treatments in normal non-cancerous cells.

Western blot of total and activated (P-EGFR) levels in UM-SCC-6 cells. Control (no treatment or EGF stimulation), EGF-stimulated, and EGF-stimulated following pre-treatment with BL or combination (BL+RA). *p<0.05 compared to EGF-stimulated control; n=3.
All therapies significantly decreased EGFR activation. Previous studies in our laboratory had reported the significant reduction in EGFR activation resulting from pre-treatment with RA (17). We used the same procedure to test the effects of BL and combination treatment on EGFR activation. As shown in Figure 2, both BL and RA +BL treatments reduced EGFR activation by ~50%.

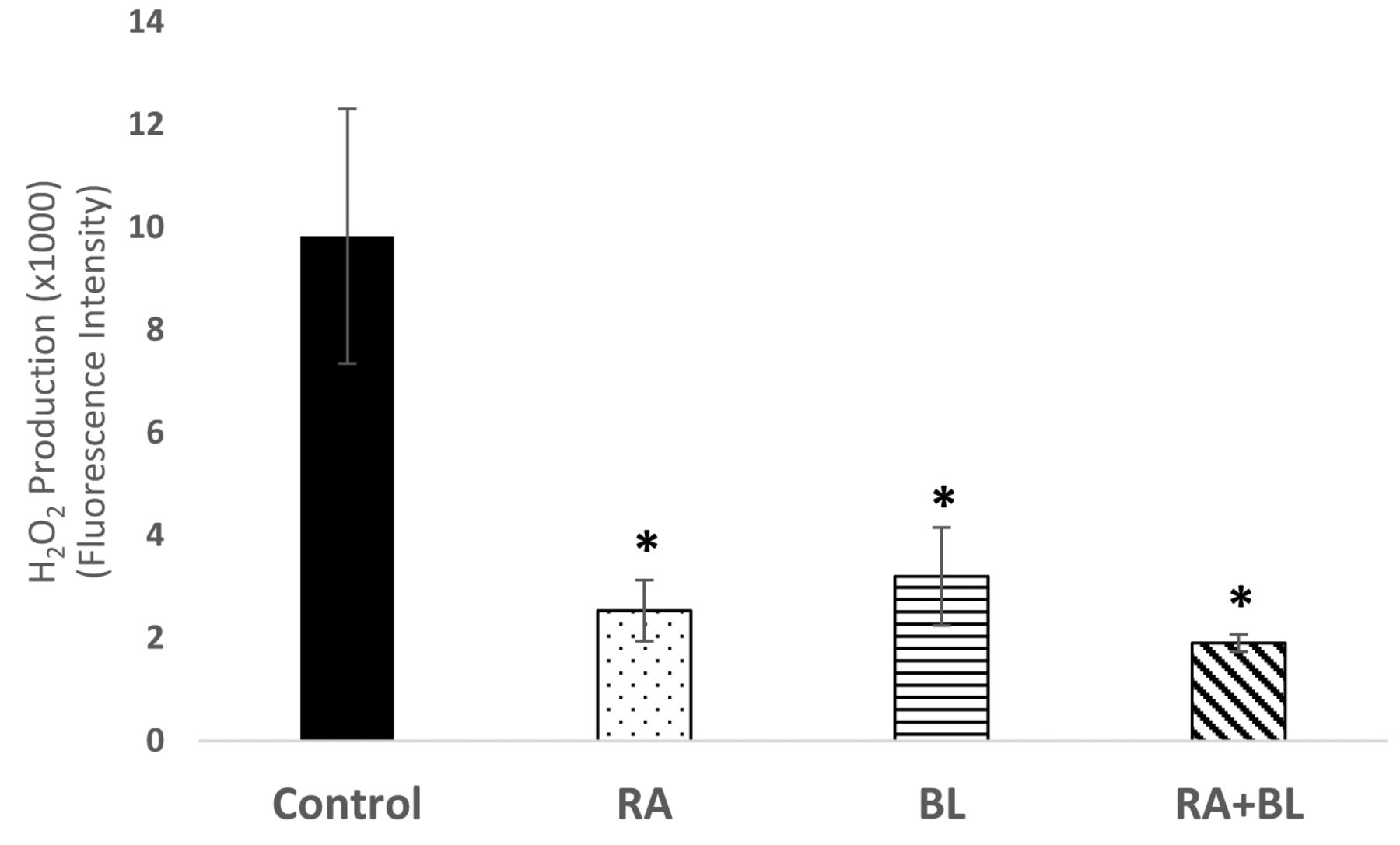
Hydrogen peroxide levels were assessed in UM-SCC-6 cells at 24 h post-treatment using Amplex Red. All treatments significantly reduced hydrogen peroxide levels; n=6 (*p<0.05).

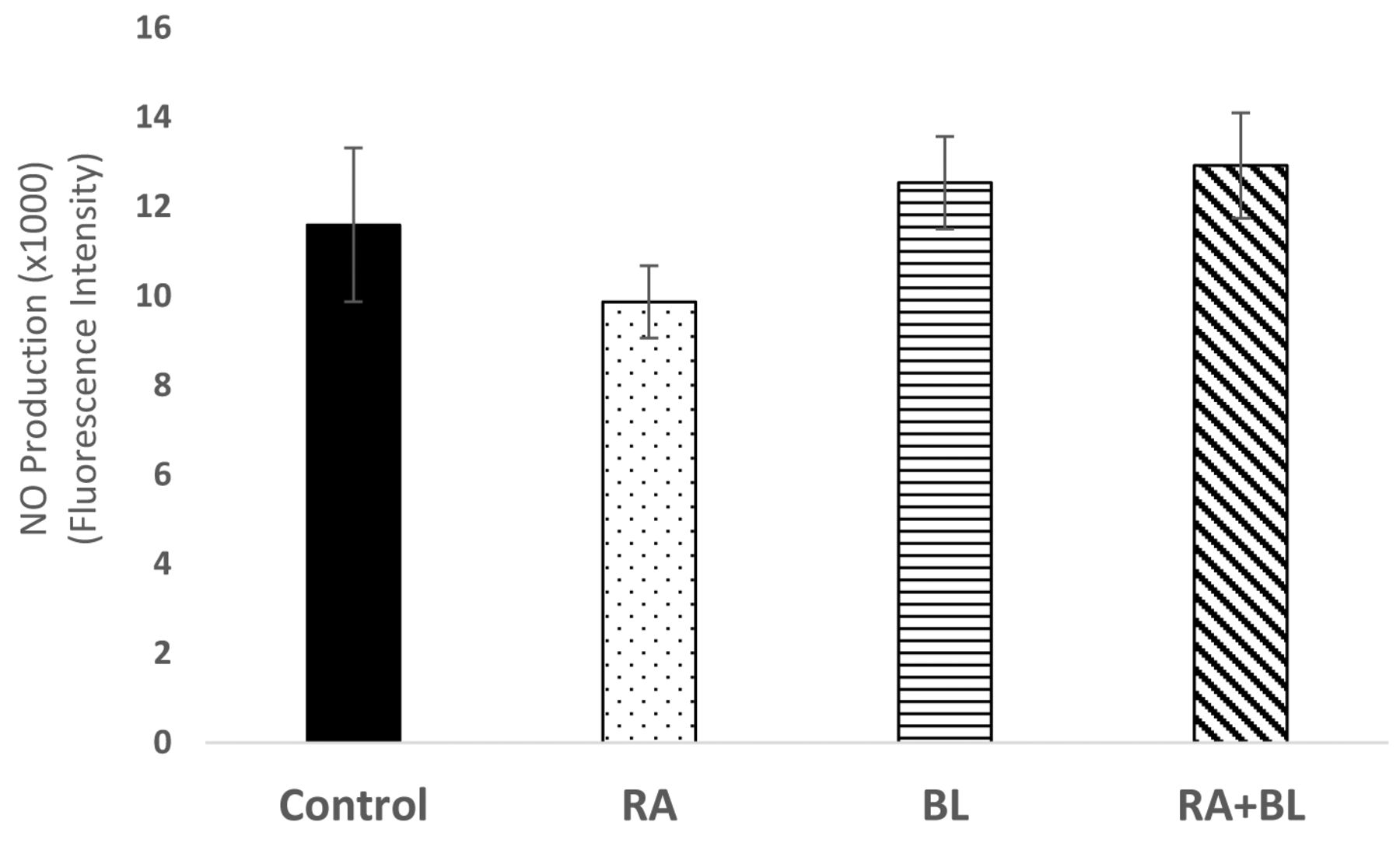
Nitric oxide levels were assessed immediately after treatment by addition of DAF-FM diacetate reagent and fluorescence quantitation (495 nm excitation/515 nm emission); n=6.
All therapies significantly decreased hydrogen peroxide levels. Hydrogen peroxide (H2O2) can be found constitutively activated in cancer cells leading to increased cell proliferation and survival (21). This constant activation can be a direct outcome of EGFR signaling due to increases in NAD(P)H oxidase (NOX) signaling, which leads to the production of superoxide ions (O2−) that can then be converted to H2O2 through superoxide dismutase (SOD) (22). At 24 h after treatment, RA and BL reduced H2O2 production by ~4- and 3-fold, respectively, while the combination treatment reduced H2O2 levels by ~5-fold (Figure 3). BL and RA may be blocking growth factor signaling and over-activating catalase, peroxiredoxins and glutathione peroxidases that would result in the observed H2O2-scavenging effect (23).

Flow cytometric analysis of AnnexinV/PI-stained UM-SCC-6 cells treated with RA, BL or combination compared to untreated negative control and staurosporine-treated (STS) positive control profiles at 12 h after treatment.
BL enhanced nitric oxide production, while RA is a nitric oxide scavenger. As shown in Figure 4, BL slightly enhanced production of nitric oxide when compared to untreated control cells, although the effect was not statistically significant. This effect was evident in both single and combination treatment groups. Conversely, RA decreased nitric oxide production, though not significantly.
Neither treatment induced apoptosis or necrosis in UM-SCC-6 cells. Flow cytometric analysis of Annexin V/propidium iodide stained cells showed no evidence of increased levels of apoptosis or necrosis in cells treated with RA, BL, or combination treatment (Figure 5). Profiles for all treatment groups were nearly identical to that of the untreated controls, while staurosporine treated cells showed significantly increased levels of apoptotic and necrotic cells.
Discussion
The aim of this study was to provide a scientific foundation for the use of BL and RA as an effective adjunctive treatment for HNSCC. The mechanisms underlying the effects of these treatments on cell proliferation of oral carcinoma were investigated by exploring EGFR activation, oxidative/nitrosative stress (H2O2/NO), and induction of apoptosis.
The last truly established therapy for HNSCC, EGFR-targeting monoclonal antibody cetuximab, appeared in 2006 (15). Epidermal Growth Factor Receptor is a cell-surface protein of the erbB family of receptor tyrosine kinases that regulates cell growth and differentiation (24). EGFR has been noted to contribute to proliferation and invasion of cancer cells and has been indicated to be overexpressed in 90% of HNSCC (25). Both RA and BL treatments significantly reduced EGFR activation and therefore could be important non-toxic components in HNSCC therapies.
Cancer cells are generally known for a marked increase in ROS levels, both as a result of an increase in metabolism and as a signaling system for cell proliferation (26). ROS is primarily compartmentalized within mitochondria since it is generated as a by-product of the electron transport system. This process releases 0.4-4.0% of consumed oxygen back into the mitochondria as hydrogen peroxide (H2O2), superoxide (O2−), singlet oxygen (O−), or the hydroxyl radical (27, 28). Anticancer therapies can be designed to increase the overall ROS to toxic levels and create debilitating cell damage, leading to the induction of apoptosis or senescence (26). Several studies have shown that increases in intracellular ROS create an H2O2 dependent induction of apoptosis (29). For instance, a non-steroidal anti-inflammatory, sulindac, has been studied as a potential treatment for colon and lung cancers due to its ability to increase intracellular ROS, resulting in a H2O2 dependent induction of apoptosis (30). Conversely, reduction of H2O2 diminishes the proliferative signal associated with growth factor activation and could reduce proliferation or induce cell senescence. RA has been shown to induce a nominal increase in antioxidative enzymes, which may contribute to RA being anti-tumorigenic (6). A similar antioxidant, genistein, has also been shown to have anticarcinogenic properties through its ability to decrease H2O2 formation in an in vivo and in vitro study (31). Similar studies using antioxidant compounds, such as caffeic acid phenethyl ester, have resulted in a decrease in hydrogen peroxide levels through a scavenging mechanism that reduced cell growth within human leukemic HL-60 cells (9, 32). Likewise, BL has been shown to induce the production of peroxiredoxins in human THP-1 monocytic cells, thus suggesting an antioxidant effect of this treatment (19). Although neither treatment significantly altered NO levels, both behaved as has been previously reported. BL has reportedly increased intracellular NO levels through direct, non-enzymatic release of NO from photolabile nitrosylated proteins (33), while RA has been identified as a NO scavenger (34, 35). Alterations in NO signaling can have far-reaching effects. For example, there is evidence to support that NO has inhibitory effects on NFĸB DNA binding (36). Downregulation of NFĸB downstream targets may serve to decrease cancer cell proliferation and metastasis.
Our results showed that both RA and BL significantly decreased cell proliferation in all HNSCC cell lines tested (Figure 1). In addition, the combination therapy was significantly more effective than either single treatment alone. It is important to note that normal human oral keratinocyte cells (HOK) were much less affected by the treatments than the tumor cell lines (Figure 1), suggesting that this combination treatment has the potential to inhibit cancer cell proliferation and growth, while resulting in minimal toxicity to normal healthy cells. Not surprisingly, the decrease in cell proliferation of UM-SCC-6 cells illustrated in Figure 1 correlated with the downregulation in EGFR activation (Figure 2) and the reduction in H2O2 levels (Figure 3) resulting from RA and BL treatment. No evidence for these treatments causing an induction in apoptosis was found (Figure 5). Taken together, these data suggest that the combination treatments are synergistically working to inhibit growth factor signaling and possibly inducing cell cycle arrest or cell senescence (19, 37).
In conclusion, to the best of our knowledge, this is the first study that confirms that treatments of BL in conjunction with RA inhibited cell proliferation in HNSCC cell lines. The underlying mechanisms have yet to be fully explored, but our findings to date suggest that these treatments show promise. Future studies will examine whether this combination treatment may provide means to reduce therapeutic doses of traditional chemotherapeutics, such as cisplatin, and thus diminish the debilitating side effects of these drugs.
Acknowledgements
The Authors thank Dr. Stephen Hsu for providing the OSC-2 cell line, the Western University of Health Sciences Intramural Research Grant Award IMR12342D, the Western University of Health Sciences Summer Student Fellowship Grant and the College of Dental Medicine for support of this research.
Footnotes
Authors' Contributions
MSBS graduate students Christi Waer and Preet Kaur performed most of the experiments. Dental students DeeDee Hui and Bonnie Le received the Western University Summer Research Fellowship and performed the initial cell proliferation analyses. Carlos Guerra contributed to organization and performance of experiments as laboratory manager/research assistant. Drs. Lewis, Henson and Tumur contributed equally to experimental design and data analysis. Dr. Dalia Saleem contributed to statistical analysis and preparation of the manuscript.
Conflicts of Interest
There are no conflicts of interest related to this study.
- Received November 15, 2019.
- Revision received January 14, 2020.
- Accepted January 15, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.