


Abstract
Background/Aim: Cancer stem cells (CSCs) are a subpopulation of cells that retain self-renewal and pluripotency capabilities, are resistant to chemotherapy, and are thought to facilitate metastasis. Target cell expression of proliferating cell nuclear antigen (PCNA) or lectin-like transcript 1 (LLT1) inhibits natural killer (NK) cell functions. The purpose of this study was to characterize the expression of LLT1 or PCNA as NK cell evasion strategies of HCT 116, a colorectal cancer cell line. Materials and Methods: Protein expression was determined by flow cytometry and/or confocal microscopy. Stem-like cells were sorted and characterized, and NK cell effector functions measured by interferon-γ secretion and cytotoxicity assay. Results: PCNA expressing cells are potential CSCs, blocking PCNA alters interferon-γ secretion, and blocking PCNA or LLT1 increases cytotoxicity. Conclusion: PCNA is a potential biomarker of stem-like colon cancer cells. Based on the results of this study, PCNA and LLT1 should be further explored as in vivo immunotherapeutic targets for NK cell-mediated killing.
Natural killer (NK) cells form the first line of defense against cancer and infections. NK cell function is dependent on a delicate balance of activating and inhibitory receptors (1). Cancer cells may evade NK-mediated killing by expressing ligands for NK cell inhibitory receptors or down-regulating the expression of ligands for activating receptors. Recent evidence has demonstrated that NKp44, a member of the natural cytotoxicity receptor (NCR) family, can function as an activating or inhibitory receptor depending on ligand interaction (2). Briefly, an isoform of mixed-lineage leukemia-5 (MLL5) is an activating ligand for NKp44 with the immunoreceptor tyrosine-based activation motif (ITAM) in DNAX activation protein of 12 kDa (DAP12) facilitating activating signals, whereas proliferating cell nuclear antigen (PCNA) is an inhibitory ligand with a putative immunoreceptor tyrosine-based inhibition motif (ITIM) on the cytoplasmic tail of NKp44, facilitating inhibitory signals (3-6). Additionally, it has been recently shown that platelet-derived growth factor (PDGF)-DD engagement of NKp44 triggers IFN-γ secretion (7).
PCNA, a homotrimeric nuclear protein, which was first identified as a sliding clamp for DNA polymerase δ, is also involved in other vital cellular functions such as chromatin remodeling, DNA repair, and cell-cycle regulation. PCNA’s critical role in cell proliferation has led to the use of PCNA as a diagnostic and prognostic marker (8). PCNA levels in cancer cell lines have been shown to be 5-6 times higher than PCNA levels in either primary or transformed cell lines (9). Additionally, our lab identified that PCNA associates with Major Histocompatibility Complex (MHC) I on the surface of the cell and forms the inhibitory ligand for NKp44, resulting in the inhibition of NK function (4, 10). Therefore, surface PCNA may be used by cancer cells as an immune evasion strategy. Recently, Shemesh et al. identified an NKp44-derived peptide, which binds to PCNA, and when fused with a cell-penetrating peptide it was able to reduce cell viability and promote death in human and murine cancers (11). This further highlights PCNA as an ideal candidate for immunotherapy, as surface expression of PCNA has not been identified in non-transformed cells, and intracellular targeting of PCNA is possible.
An additional immune evasion strategy used by cancer cells is surface expression of lectin-like transcript 1 (LLT1/CLEC2D). LLT1 binds to NKRP1A (CD161) and functions as an NK cell inhibitory ligand (12-15). LLT1 expression has been described by others as an immune escape in glioma cell lines (16). Recently, our lab also demonstrated the role of LLT1 in immune evasion in breast and prostate cancer (17, 18).
In 2020, it is estimated that approximately 9% of all cancer-related deaths will be due to colorectal cancers. Colorectal cancer has metastasized in 21% of patients at diagnosis leading to a 5-year relative survival rate of 14% (19). Although these data seem bleak, the developments of cancer screening, treatment, and prevention has increased the 5-year relative survival rate of colorectal cancer from 50% during 1975-1977 to 64% during 2009-2015 (19). The increased survival rate can be attributed to early screening and detection as well as better treatments. These statistics highlight the importance of research on colorectal cancer metastasis and metastasis specific treatments.
Metastasis occurs when tumor cells undergo epithelial-mesenchymal transition (EMT), a process in which differentiated epithelial cells dedifferentiate into mesenchymal cells with stem cell properties, which include enhanced migratory capacity, invasiveness, and resistance to apoptosis and chemotherapy (20, 21). EMT has been documented in many in vitro studies, but the acceptance and significance of EMT in human disease has been greatly debated until recent years. Evidence for EMT in vivo has been documented in mouse models of breast cancer and human colon carcinoma. Additionally, direct in vivo imaging has yielded evidence to support EMT in cancer progression and metastasis (22). Our lab has previously demonstrated that cells with surface PCNA have increased Matrigel invasion compared to cells lacking surface PCNA. This demonstrates that cells with surface PCNA are more invasive and may be the CSC population (data unpublished).
Recently, NANOG, SOX2, and Oct-4, transcription factors associated with self-renewal and pluripotency of embryonic stem cells, along with other markers such as CD44 and CD133, have been used to identify CSC populations (23-26). CD44 is a cell surface glycoprotein used primarily for cell-cell and cell-matrix interactions by binding to hyaluronan (HA), but also has many other functions depending on the extracellular structure, which can vary due to alternative splicing and post-translational modifications (27). Recently, CD44 has been identified as regulating TLR2 activation of macrophages and inhibition of proinflammatory cytokine production when CD44 interacts with HA or a CD44-specific antibody (28). CD133 (prominin-1) is a member of the pentaspan transmembrane glycoproteins, and is localized to membrane protrusions, such as microvilli. As such, it is hypothesized that CD133 plays a role in organizing membrane topology (26). While the exact function of CD133 remains unknown, it has been implicated in many types of cancer as a cancer stem cell marker. A recent meta-analysis also demonstrated that CD133 expression is positively correlated with metastasis, vascular invasion, and lymphatic invasion in colorectal cancer patients. Additionally, patients with higher CD133 expression had poorer survival and a lower 5-year overall survival rate (29).
Materials and Methods
Cell culture. HCT 116 (ATCC CCL-247) cells were maintained in McCoy’s 5A media, supplemented with 10% fetal bovine serum (FBS; Gemini Bio-Products, West Sacramento, CA, USA), and antibiotic-antimycotic solution containing penicillin G, streptomycin sulfate, and amphotericin B (Life Technologies, Carlsbad, CA, USA). DB (ATCC CRL-2289) cells were maintained in RPMI 1640 supplemented with 10% FBS (Gemini Bio-Products), 10 mM HEPES (Life Technologies), 10 mM Sodium Pyruvate (Life Technologies), 10 mM non-essential amino acids (Life Technologies), and antibiotic-antimycotic solution. NK-92MI (ATCC CRL-2408), a human NK cell line constitutively expressing IL-2, was maintained in Minimum Essential Medium Alpha Medium (Life Technologies) supplemented with 12.5% fetal bovine serum (Gemini Bio-Products), 12.5% horse serum (Gemini Bio-Products), 0.2 mM inositol (Sigma-Aldrich, St. Louis, MO, USA), 0.1 mM 2-mercaptoethanol (Sigma-Aldrich), and 0.02 mM folic acid (Sigma-Aldrich). All cells were cultured at 37°C in a humidified 5% CO2/95% air environment.
Flow cytometry. Cells were first incubated with 1 μg Human IgG Fc fragment (Rockland, Gilbertsville, PA, USA) to block nonspecific interactions. For flow cytometry, cells were then incubated with 1 μg Rabbit-anti-Human-CD133 (Proteintech, Rosemont, IL, USA) followed by 0.2 μg Donkey-anti-Rabbit-IgG-PE (BioLegend, San Diego, CA, USA), 1 μg anti-PCNA-Alexa Fluor 488 (BioLegend), or 0.2 μg anti-CD44 (BioLegend). The following isotype controls were used: Rabbit IgG (Rockland), mIgG2a-AF488 (BioLegend), and rIgG2b-APC (BioLegend). Flow cytometry was performed using a Cytomics FC500 Flow Cytometer (Beckman Coulter, Brea, CA, USA) and results were analyzed using FlowJo Software v10.1 (FlowJo, Ashland, OR, USA). The median florescence intensity (MFI) was also quantified.
Cell sorting. Cells were first incubated with Human IgG Fc fragment (Rockland) to block non-specific interactions. Cells were then incubated with 1 μg Rabbit-anti-Human-CD133 (Proteintech, Rosemont, IL, USA), washed twice, incubated with 0.2 μg Donkey-anti-Rabbit-IgG-PE (BioLegend), 1 μg anti-PCNA-Alexa Fluor 488 (BioLegend), and 0.2 μg anti-CD44 (BioLegend), and washed twice. Sorting was performed using a SH800 Cell Sorter (Sony, Biotechnology, San Jose, CA, USA). After sorting, cells were washed in 5 ml complete media five times to remove bound antibodies. Removal of bound antibodies was confirmed by flow cytometry.
Confocal microscopy. HCT 116 cells were cultured on gelatin coated coverslips for 48 h, fixed with 2% paraformaldehyde, incubated with 1 μg anti-PCNA-AF488 (BioLegend) or 0.125 μg anti-CD44-PE (BioLegend) and followed by staining with Hoechst 33342 (Thermo Fisher Scientific, Waltham, MA, USA) following the manufacturers’ protocol, and imaged on the LSM 510 Confocal Laser Microscope (Zeiss, Dublin, CA, USA) using the 40×, 1.2 NA, 0.28 WD (water).
Quantitative real-time (RT) PCR. RNA was isolated from sorted cells using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s protocol. cDNA was synthesized using SuperScript IV VILO Master Mix with ezDNase Enzyme (Invitrogen) following the manufacturer’s protocol. RT-PCR was performed using TaqMan Gene Expression Master Mix (Applied Biosystems, Foster City, CA, USA) and TaqMan primers and probes for SOX2 (Hs01053049_s1), POU5F1 (Hs00999632_g1), and NANOG (Hs04399610_g1). GAPDH (Hs02758991_g1) was used as a housekeeping control and B lymphoblastoid cells were used for normalization. Fold change was calculated using the delta-delta Ct method.
51Cr release cytotoxicity assay. Target cells and NK92-MI cells were incubated with Human IgG Fc fragment to block non-specific interactions prior to use. Target cells were labeled with 51Cr for 1 h at 37°C and then, incubated with either 1 μg anti-PCNA (BioLegend) or mIgG2a isotype control (BioLegend). Target cells were then incubated with NK cells at E:T ratios of 25:1, 5:1, and 1:1 for 4 h at 37°C. NK cells were previously confirmed to express NKp44. Percent specific lysis was compared to target cells incubated with isotype antibody or with no antibody as a positive control (no blocking) of cell lysis under unblocked conditions. Alternatively, NKp44 was blocked on NK cells with 1 μg anti-NKp44 antibody (BioLegend) or 1 μg mIgG1 isotype control (BioLegend) antibody prior to incubation with target cells. Supernatants were collected and percent specific lysis was calculated. Experiments were performed in triplicate.
Interferon-γ release assay. Target cells were sorted as previously stated. Sorted cells were then washed 5 times in 10 ml warmed complete media to remove bound antibodies. PCNA, NKp44, or isotype control blocking antibodies were used as previously stated. NK cells were added at an E:T ratio of 1:3 and co-incubated for 18 h. After 18 h, supernatant was collected and an interferon-γ ELISA was conducted per the manufacturers’ protocol (ELISA MAX, BioLegend, San Diego, CA, USA).
Statistical methods. Statistical analysis was performed through Prism 6.0 (GraphPad Software, San Diego, CA, USA). Specific statistical tests used are stated in figure captions. Differences were considered significant at p-value <0.05.
Results
HCT 116 cells express ligands for NK cell receptors. Flow cytometry was performed to verify that HCT 116 cells express ligands for NK cell receptors. HCT 116 cells were incubated with conjugated antibodies against PCNA and LLT1 (Figure 1A). A Human Diffuse B cell lymphoma (DB) cell line was used as a control (Figure 1C and D), as we have previously characterized PCNA and LLT1 expression (10, 30). HCT 116 cells consistently express the cell surface ligands PCNA and LLT1 (Figure 1B). On average, 8% of HCT 116 cells express surface PCNA (data not shown). In an effort to evaluate the relative expression of surface PCNA on this small population, we analyzed the MFI ratio of PCNA+ cells (Figure 1B). As suspected, the MFI ratio of PCNA+ cells was significantly higher in both HCT 116 and DB cells when compared to the PCNA expression of the total cell population; illustrating cells with surface PCNA express a significantly higher amount of surface PCNA. Additionally, surface expression of PCNA was confirmed through confocal microscopy (Figure 1E). Cells were cultured on a gelatin-coated cover slip and incubated with conjugated antibodies. Cells were not permeabilized to prevent anti-PCNA antibodies binding to cytoplasmic or nuclear PCNA. CD44 and Hoechst 33342 were used as a surface marker and nuclear marker, respectively.

HCT 116 cells express PCNA and LLT1. Cells were first blocked with Human IgG Fc fragment to prevent reverse binding of conjugated antibodies. Cells were then incubated with 1 μg PCNA-AF488 or LLT1-PE (filled histograms). Appropriate isotype controls are shown in empty histograms (A) HCT 116 cells express PCNA and LLT1. (B) Cumulative data from analysis shown in (A). PCNA refers to the total cell population, and PCNA+ refers to cells expressing PCNA. (C) DB cells express PCNA and LLT1. (D) Cumulative data from analysis shown in (C). (E) Cells were cultured on a gelatin-coated cover slip, incubated with PCNA-AF488, CD44-PE, and Hoechst 33342 and viewed under confocal microscopy. Cells were not permeabilized to prevent intracellular PCNA staining. CD44 and Hoechst were used as a surface marker and nuclear marker, respectively. Median fluorescent intensity (MFI) ratio is indicated in the top right corner of all histogram plots; calculated as (MFI positive cells) / (MFI negative cells). Data are represented as means±SD (n≥3 independent experiments); mean value listed above columns; p values calculated by unpaired two-tailed Student’s t test. *p≤0.05, **p<0.01, ***p<0.001.
We then sought to confirm if HCT 116 cells express the CSC markers CD44 and CD133 as previously reported (31). HCT 116 (Figure 2A) consistently express CD44 and CD133. We then wanted to elucidate if cells co-express PCNA, CD44, and CD133. On average, 0.74% of cells are triple positive (PCNA+CD44+CD133+) and 3.5% of cells are double positive (PCNA–CD44+CD133+) (Figure 2C). Interestingly, the double positive population, which is currently regarded as the CSC population for colorectal cancer, is significantly larger than the triple positive population (32). The gating strategy to determine these subpopulations is also used for fluorescence activated cell sorting (FACS) (Figure 2B). Our lab has previously observed that cells grown to 90% confluency begin to naturally detach and the percent of PCNA+ cells is greater in cells that naturally detach versus cells that are detached via scraping (unpublished data). Based on this observation, we wanted to determine if the triple positive population is greater in cells that naturally detach. There is a 3.5-fold increase in the triple positive population of naturally detached cells compared to cells detached by scraping, and this increase is statistically significant (Figure 2D and E). Collectively, these data suggest that cells with surface PCNA may be more stem cell-like than cells lacking surface PCNA.
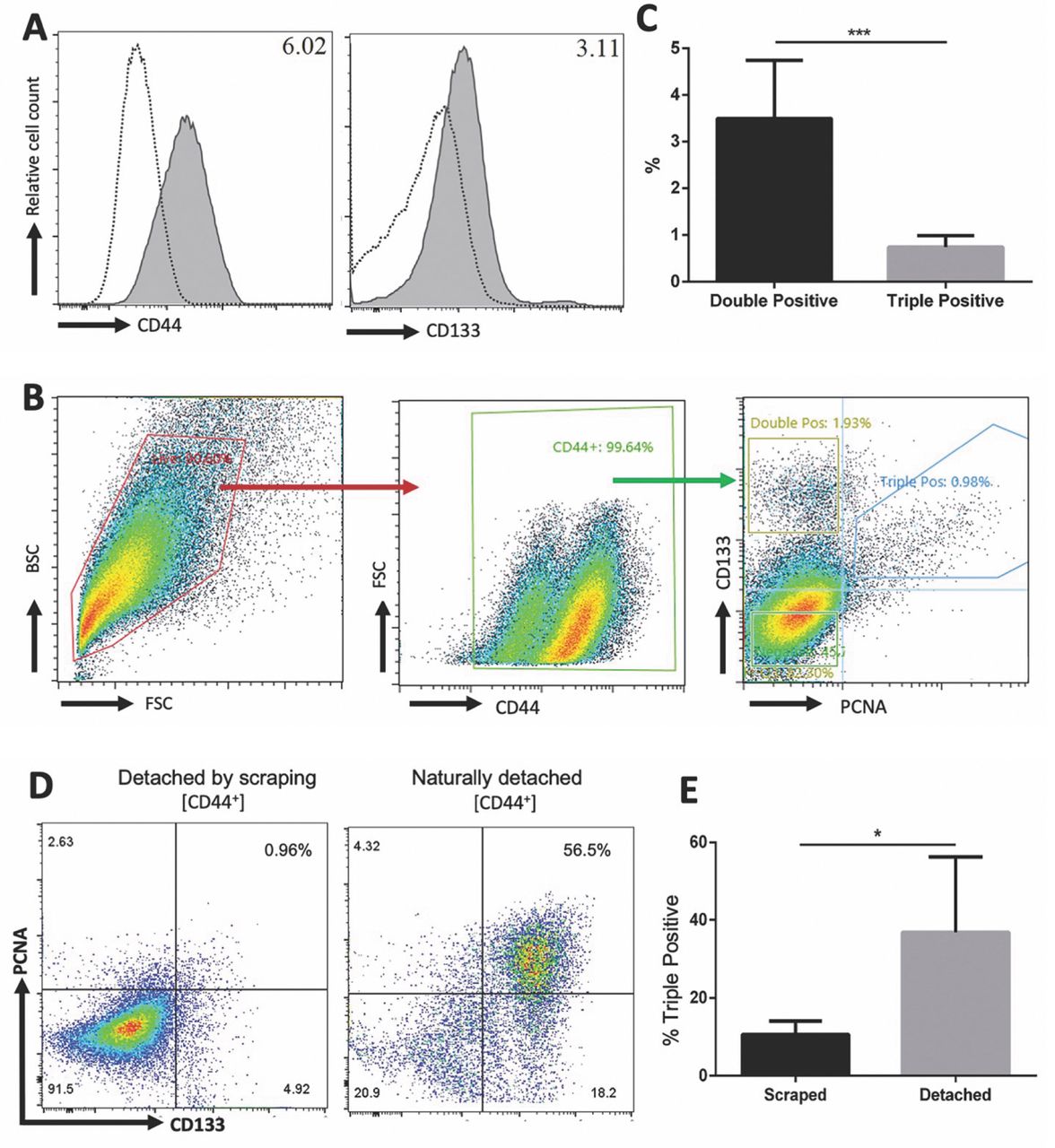
HCT 116 cells express cancer stem cell (CSC) markers CD44 and CD133 and the CSC population increase in naturally detached cells. Cells were first blocked with Human IgG Fc fragment to prevent reverse binding of conjugated antibodies. Cells were then incubated with 0.5 μg CD44-APC, 1 μg PCNA-AF44 and/or 1 μg rabbit anti-human CD133 and detected with donkey anti-rabbit PE (filled histograms). Appropriate isotype controls are shown in empty histograms (A) HCT 116 cells express CD44 and CD133. (B) Gating strategy to determine triple positive (PCNA+CD44+CD133+) and double positive (PCNA-CD44+CD133+) subpopulations. (C) The double positive subpopulation is consistently larger than the triple positive population. (D) HCT 116 cells were grown to near confluency and cells flowing in media were collected; the attached cells were scraped off. Naturally detached HCT 116 cells have an increase in triple positive cells compared to cells detached by scraping. (E) Cumulative data of (D) for triple positive cells. Median fluorescent intensity (MFI) ratio is indicated in the top right corner of all histogram plots; calculated as (MFI positive cells)/(MFI negative cells). Data are represented as means±SD (n≥3 independent experiments). p-values calculated by unpaired two-tailed Student’s t-test (C) and ratio paired two-tailed Student’s t-test (E). *p≤0.05, **p<0.01, ***p<0.001.
Expression of CSC transcription factors in sorted populations varies among cancer cell lines. Zhou et al. identified that CD44+CD133+ HCT 116 cells express higher levels of CSC-related genes (31). To determine if there is differential expression of CSC transcription factors in sorted populations, we used DB cells, which have given consistent PCR results in the past. DB cells were sorted via FACS, RNA was isolated from PCNA+ and PCNA– cells, mRNA was converted to cDNA, and qRT-PCR was performed to determine the levels of NANOG, SOX-2, and Oct-4. In line with our previous studies, PCNA+ DB cells expressed significantly higher levels of NANOG, SOX-2 and Oct-4 compared to PCNA– DB cells (Figure 3A). When HCT 116 cells were sorted on PCNA expression, no difference was seen between PCNA+ and PCNA– cells (data not shown). To examine if there was a difference in CSC transcription factor expression, cells were sorted into triple positive (PCNA+CD44+CD133+), double positive (PCNA–CD44+CD133+), and negative (PCNA– CD44+CD133–) subpopulations. Although HCT 116 cells did not have any statistically significant difference between subpopulations, a consistent trend was observed. Interestingly, triple positive cells had higher levels of CSC transcription factors than double positive cells, which are typically regarded as the CSC population in literature (Figure 3B). These data suggest that surface PCNA expression may in fact be a marker of CSC.

Surface PCNA is associated with stem cell-like phenotype, alters NK cell IFN-γ secretion, and blocking PCNA, NKp44, or LLT1 increases NK cytotoxicity. (A) PCNA+ DB cells have significantly higher expression of cancer stem cell (CSC) associated transcription factors NANOG, SOX-2, and Oct-4 than PCNA-DB cells. (B) While there is no difference between populations, there is a trend that triple positive HCT 116 cells express higher levels of CSC transcription factors, double positive cells express an intermediate level, and negative cells express the lowest levels. Cells were first blocked with Human IgG Fc fragment to prevent reverse binding of conjugated antibodies. Cells were then incubated with 0.5 μg CD44-APC, 1 μg PCNA-AF44 and/or 1 ug rabbit anti-human CD133 and detected with donkey anti-rabbit PE; cells were then sorted via FACS and then washed to remove bound antibodies. (C) HCT 116 cells were either unsorted or sorted into triple positive (PCNA+CD44+CD133+), double positive (PCNA-CD44+CD133+), and negative (PCNA–CD44+CD133–) subpopulations. Sorted cells were then incubated with NK-92MI cells for 18 h in the presence of anti-PCNA, anti-NKp44, or isotype control antibodies, and interferon-γ secretion was determined by ELISA. Blocking PCNA decreased IFN-γ secretion in the triple positive, negative, and unsorted populations. Blocking NKp44 increased IFN-γ secretion in the unsorted population. (D) Target cells were loaded with 51Cr and incubated with 1 μg anti-PCNA, 1 μg anti-LLT1 antibody, or isotype control. NK cells were incubated with 1 μg anti-NKp44, 1 μg isotype control, or no antibodies. Target cells were then incubated with NK cells at 25:1, 5:1, and 1:1 effector to target (E:T) cell ratios for 4 h at 37°C. Killing was compared to cells incubated with isotype antibody. Data are represented as means±SD; n≥3 independent experiments (panels A to C). Data as means±SEM; one independent experiment performed in triplicate (panel D). p values calculated by unpaired two-tailed Student’s t-test. *p≤0.05, **p<0.01, ***p<0.001.
Surface PCNA alters IFN-γ secretion. Our previous studies demonstrated that DB cells expressing surface PCNA were able to inhibit cytotoxicity of both primary human NK cells and the NK92-MI cell line (10). Additionally, it has been demonstrated that PDGF-DD binding to NKp44 stimulates IFN-γ secretion (7). Therefore, we sought to investigate if NK cell effector functions were altered in PCNA expressing HCT 116 cells and whether there is a difference in subpopulations. HCT 116 cells were sorted into triple positive, double positive, and negative subpopulations. Unsorted cells were used as a control. Sorted cells were then washed 5 times to remove bound antibodies. To block PCNA-NKp44 interactions, sorted or unsorted cells were incubated with either 1 μg IgG2a isotype control antibody or 1 μg anti-PCNA antibody and NK cells were incubated with either 1 μg IgG1 isotype control antibody or 1 μg anti-NKp44 antibody. After 30 min, cells were incubated with NK-92MI cells at an E:T ratio of 1:3 for 18 h. Supernatants were then collected, and an IFN-γ ELISA was performed. Interestingly, in HCT 116 cells, blocking PCNA led to a statistically significant decrease in IFN-γ secretion in the triple positive, negative, and unsorted subpopulations (Figure 3C, left). Blocking PCNA on HCT 116 cells also led to a decrease in IFN-γ secretion in the double positive population, but the decrease was not statistically significant. This decrease in IFN-γ secretion implicates that PCNA may function as an activating ligand in context with other ligand interactions. Blocking NKp44 led to a statistically significant increase in IFN-γ secretion when co-incubated with unsorted HCT 116 cells (Figure 3C, right). This increase in IFN-γ secretion may be attributed to soluble NKp44 ligands in the supernatant, potentially PDGF-DD, however supernatants were not tested for soluble ligands. Presumably, blocking NKp44 prevents inhibition by preventing PCNA-NKp44 interaction, however, this antibody may also trigger stimulation by cross-linking receptors, which may also increase IFN-γ secretion.
Surface PCNA and LLT1 function as NK cell immune evasion strategies. A standard chromium release assay was performed to determine if blocking PCNA, LLT1, or NKp44 would alter cytotoxicity in NK-92MI cells. Briefly, target cells were loaded with 51Cr and incubated with blocking antibodies or isotype control antibodies and NK cells were incubated with blocking antibodies, isotype control, or no antibodies. Target cells were then incubated with NK cells at 25:1, 5:1, and 1:1 effector to target cell (E:T) ratios for 4 h at 37°C. Blocking PCNA led to a statistically significant increase in specific lysis at the 5:1 ratio when compared to isotype control (Figure 3D). Additionally, blocking PCNA increased specific lysis at the 25:1 and 1:1 ratios, but statistical significance was not reached (p=0.06). Blocking NKp44 on NK cells led to a statistically significant increase in specific lysis at the 25:1 and 5:1 ratios. Blocking LLT1 on target cells increased specific lysis at all ratios, and the increased lysis was statistically significant (25:1 p=0.05, 5:1 p=0.02, 1:1 p=0.02). Interestingly, cells do not co-express PCNA and LLT1, suggesting that CSC may preferentially express PCNA as an evasion strategy (data not shown). These data confirm that PCNA and LLT1 can function as immune evasion ligands and blocking LLT1 or the PCNA-NKp44 interaction can increase lysis of target cells.
Discussion
NK cells are a unique population of lymphocytes, which are one of the first to respond to infection and tumor formation. NK cells play a pivotal role in immune surveillance by distinguishing healthy and cancerous cells through interaction of cell surface ligands with activating and inhibitory receptors on the NK cell surface. NKp44 and NKp30, NCRs which were originally described as activating receptors, also function as inhibitory receptors. NKp44 can bind to the PNCA-MHC I complex, and a soluble fragment of BCL2-associated athanogene 6 (BAG-6, also known as BAT3) can bind to NKp30 and inhibit cytotoxicity and cytokine secretion (10, 33).
The last few years has seen many advances in the understanding of cancer immune evasion strategies. It has also been demonstrated that there is a loss of Fas expression during early cancer development in colorectal cancer, as well as mutations in HLA class-1 antigen presentation (34, 35). Surface PCNA and LLT1 expression may also be functioning as an immune evasion strategy by colorectal cells. It is now well established that tumor cells express ligands for NK cell inhibitory receptors and thus, evade NK cell-mediated killing. We have demonstrated that prostate cancer and triple negative breast cancer evade NK cell-mediated killing by expressing LLT1 on the cell surface (17, 18). Tumor cells also escape NK cell-mediated killing by secreting soluble ligands for NK cell activation receptor NKG2D (36). Several studies have confirmed that in various cancers, including cervical cancer, pancreatic cancer, and melanoma, NKG2D receptor expression is significantly decreased on NK cells and CD8+ T cells, thereby promoting tumor immune escape (37). Changes in NKG2D expression in NK cells may be regulated by a variety of factors, including changes in cellular activity factors and the physicochemical features of the tumor microenvironment.
In this report, we identified that colon cancer (HCT 116) cells with cell surface PCNA also express CSC markers CD44 and CD133 as well as increased expression of CSC transcription factors NANOG, SOX-2, and Oct-4. Colon cancer cells expressing surface PCNA are enriched in naturally detached suspension cells, compared to attached cells, indicating cell surface PCNA may be a marker for metastasizing cancer stem cells. Additionally, cell surface PCNA or LLT1 may facilitate escape from NK cell killing by transmission of an inhibitory signal. Blocking LLT1 or the NKp44-PCNA interaction with antibodies enhanced the specific killing of cancer cells by NK cells. Interestingly, blocking PCNA decreased IFN-γ secretion by NK cells co-incubated with PCNA+CD44+CD133+ and PCNA– CD44+CD133– HCT 116 cells; thus, implicating PCNA may also function as an activating ligand. Although these findings are promising, future in vivo studies utilizing immune deficient mice will be required to confirm the tumorigenesis ability of PCNA expressing cells. This study suggests that LLT1 or PCNA based immunotherapy might be beneficial in conjunction with traditional chemotherapy for colorectal cancers. A clear understanding of the NK receptor ligands on colorectal cancer cells will help lead to a more personalized treatment plan.
Acknowledgements
This study was supported in part by the National Institute of Health/National Institute of Neurological Disorders, Bethesda, MD, USA and Stroke grant R03 NS101481 awarded to Porunelloor A. Mathew. Additionally, Joseph D. Malaer received 12 months of support from the predoctoral grant number T32 AG020494-16A1 awarded to UNTHSC by the National Institute of Health/National Institute on Aging, Bethesda, MD, USA.
Footnotes
Authors’ Contributions
Joseph D. Malaer: Conceptualization, data curation, methodology, formal analysis, writing – original draft, writing – review and editing. Porunelloor A. Mathew: conceptualization, funding acquisition, formal analysis, writing – review and editing.
This article is freely accessible online.
Conflicts of Interest
The Authors have no conflicts of interest to declare regarding this study.
- Received October 29, 2020.
- Revision received November 11, 2020.
- Accepted November 12, 2020.
- Copyright © 2020 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.