


Abstract
Background/aim: Autophagy can be either tumor promotive or suppressive. We previously identified an autophagy-inducing activity in the 30-100 kDa fraction of areca-nut-extract (ANE 30-100K) and showed that several tumor cells subjected to chronic ANE 30-100K stimulation (CAS) exhibited higher resistance against stressed environments including serum-free (SF) conditions in vitro. Herein, we aimed to assess whether CAS can also provide growth advantages for tumor cells in vivo and the therapeutic effect of autophagy inhibition on CAS-treated tumors. Materials and Methods: Esophageal CE81T/VGH cells and nude mice were used as experimental models. Autophagy inhibitors 3-methyladenine (3-MA) and chloroquine (CQ), as well as another anticancer drug cisplatin (DDP), were chosen to challenge CAS-treated CE81T/VGH cells in vitro and in vivo. Results: CAS-treated CE81T/VGH cells expressed higher levels of microtubule-associated protein 1 light chain 3A/B-II (LC3-II) and beclin 1 proteins, and showed stronger resistance to SF and hypoxia conditions, that were mitigated by CQ or 3-MA in vitro. Furthermore, CAS-treated CE81T/VGH cells induced significantly larger tumors in mice, which were also attenuated by single 3-MA or CQ treatment. Finally, the combined treatment of 3-MA or CQ with DDP further up-regulated DDP-induced caspase-3 activity in vitro and exhibited synergistic anti-tumor effects on mice. Conclusion: CAS may up-regulate tumoral autophagy and provide growth advantage for tumors both in vitro and in vivo. Furthermore, autophagy inhibition alone or in combination with DDP may achieve positive therapy for tumors encountered with CAS.
Autophagy is a conserved mechanism for cells to maintain homeostasis by degrading and recycling cytoplasmic proteins and organelles. Abnormality of autophagy can lead to diseases including diabetes, autoimmunity, cardiovascular diseases, myopathies, neurodegeneration, and cancers (1, 2). Although autophagy was originally thought to inhibit tumor growth, it is now known to play both suppressive and promotive roles in tumor initiation and progression (3). In a tumor-suppressive aspect, autophagy removes damaged mitochondria to prevent excessive production of reactive oxygen species (ROS) and DNA mutation; therefore, reduction or malfunction of autophagy leads to genomic instability, oxidative stress, and activation of DNA damage response (4-6). On the other hand, autophagy can increase the survival rate for invasive or metastasizing tumors under serum-limited environment (3, 7). Thus, defining the role of autophagy in individual cancers is essential for application of autophagy-modulated therapy (3).
Areca nut (AN) is an addictive human carcinogen used globally by about 600 million people and thought to cause various oral mucosal diseases such as oral cancers through the induced oxidative stress (8). Chewing AN is also associated with increased incidence of esophageal carcinomas as well as younger age of diagnosis, poor chemoradiotherapy response, and shorter overall survival (9, 10). ANE is mainly composed of carbohydrate, polyphenols, and lipids. Ingredients of AN have been shown to induce elevation of ROS levels, inflammation, genotoxicity, growth arrest, senescence, and apoptosis. Although small molecules like alkaloids present in ANE are less than 1%, they are thought to be the most active ingredients associated with AN-related pathology (11). However, in our studies, cytotoxic concentrations of both AN extract (ANE) and its 30-100 kDa fraction (ANE 30-100K) were shown to induce swollen cell morphology, condensed nucleus, microtubule-associated protein 1 light chain 3A/B-II (LC3-II) accumulation, acidic vesicles, autophagic vacuoles, and empty cytoplasm, before cell death, in several types of cancer cells (12). Moreover, ANE 30-100K-induced autophagy was also illustrated to be sensitive to both cellulase and proteinase K digestion suggesting that the active molecule contains both carbohydrate and protein moiety (13). We also showed that ANE 30-100K was capable of triggering autophagic flux through ROS in both normal fibroblasts and malignant cells, and ANE 30-100K-induced autophagy was beclin-1, Atg5, and MEK/ERK pathway dependent (14, 15).
Whether ANE 30-100K is involved in pathological development remains undefined. We speculated that tumor cells that survived from chronic ANE 30-100K stimulation (CAS) might be endowed with some growth advantages under stress conditions. Our previous and preliminary in vitro studies showed that chronic treatment of different malignant cells with sublethal concentrations of ANE 30-100K exhibited higher tolerance against glucose deprivation, hypoxia, cisplatin (cis-diamminedichloroplatinum, DDP), 5-fluorouracil, and serum starvation. The increased resistance of drugs and serum depletion were diminished in the presence of autophagy inhibitors including 3-methyladenine (3-MA) and chloroquine (CQ) (15, 16).
In this study, we further assessed the impact of CAS on an esophageal carcinoma cell line, CE81T/VGH, both in vitro and in vivo, and analyzed the effects of autophagy inhibition (by 3-MA or CQ) alone or in combination with DDP on CAS-treated cells.
Materials and Methods
Preparation of ANE 30-100K. In Southeast Asia, mature (ripe) AN is commonly used for chewers; however, immature (tender) AN is also popular in Taiwan. We extracted one tender AN in a earthenware basin with 5 ml PBS (phosphate-buffered saline, containing 1.8 mM KH2PO4, 150 mM NaCl, 8.1 mM Na2HPO4, 2.7 mM KCl, pH 7.4) at room temperature, and the extracted solution was centrifuged at 12,000 g for 10 min at 4°C. The supernatant was used as ANE (areca-nut-extract). Next, ANE was filtered through cotton to remove the insoluble materials and then centrifuged at 2,900 g for 30 min at 4°C with 100 kDa-pored concentration tube (Z677906, Sigma-Aldrich Corp., St Louis, USA). The flow-through fraction was collected and further subjected to centrifugation as above with 30 kDa-pored concentration tube (430-30, Sigma-Aldrich). To remove small molecules like salts and phytochemicals, the materials of AN retained in 30 kDa-pored tube were washed twice with 2 ml H2O. The remaining substances were dissolved in water and lyophilized, designated as ANE 30-100K. Finally, the dried powder of ANE 30-100K was weighed and stored at −80°C.
Cell culture and treatment. Esophageal carcinoma CE81T/VGH cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (11995-065, Life Technologies Inc., Gibco/BRL Division, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS) (16000-044, Life Technologies Inc.) at 37°C in a humid atmosphere with 5% CO2.
For cell viability analysis, CE81T/VGH cells were seeded into a 96-well plate in a 5,000 cells/well manner and cultured for 24 h. Cells were then cultured in serum-free (SF) medium for another day, followed by various treatment and finally assessed by XTT reagents (11465015001, Roche Molecular Biochemicals, Indianapolis, IN, USA) as instructed by the manufacturer. As for Western blot analysis, 7×106 cells were seeded in a 10-cm plate for 24 h, followed by the same treatments as those of viability analysis.
Animal studies. Animal studies were conducted after the permission of the Institutional Animal Care and Use Committee (IACUC) of Chi Mei Medical Center, Approval No. 105122606, Duration: 2017/08/01~2020/07/31. We bought 6-week-old male Balb/c nude mice from BioLASCO Taiwan Co., Ltd. CE81T/VGH cells to be inoculated into mice were stimulated with or without ANE (10 μg/ml) and ANE 30-100K (1.75 μg/ml) for three weeks. After two days of mice arrival, untreated, ANE-treated, and CAS-treated 2.5×106 CE81T/VGH cells were suspended in 100 μl DMEM and mixed with equal volume of matrigel (RCF354248, Matrigel Matrix, Life Science, Marlborough, MA, USA), and then subcutaneously injected into the right flank of each mouse (n=6). Mice were sacrificed 6 weeks after tumor inoculation. Tumor volume was calculated weekly from caliper measurements by following the formula: volume=(W2×L)/2.
For subsequent drug treatment, mice inoculated with CAS-treated cells were randomly divided into four groups (n=6). They were intraperitoneally injected with PBS, CQ (60 mg/kg/day), DDP (5 mg/kg/week), and CQ+DDP (CQ administered first, followed by DDP after 20 min) one week after inoculation. Mice were sacrificed 18 days after treatment. Alternatively, these procedures were repeated except that CQ was substituted by 3-MA (30 mg/kg/day) and n=5.
Western blot analysis. Lysate proteins (20 μg) from treated cells were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto a polyvinylidene difluoride (PVDF) filter. This filter was cut into lower and upper pieces and then respectively immunoblotted with the antibodies of LC3 (L7543, Sigma-Aldrich, 2,000-fold diluted) and β-actin (A5441, Sigma-Aldrich, 10,000-fold diluted). The secondary horseradish peroxidase-coupled goat anti-mouse-IgG (AP124P, Merck Millipore, Darmstadt, Germany) for β-actin antibody was 10,000-fold diluted and goat anti-rabbit-IgG (81-6120, Invitrogen Corporation, Camarillo, CA, USA) 2,000-fold diluted for LC3 antibody. Signals of LC3 and β-actin proteins were developed with Western Lightening Chemiluminescence Reagent (NEL103E001EA, Perkin–Elmer Life Sciences, Inc., Boston, MA, USA) and then digitalized with UnSCAN-IT software Automated Digitizing System, version 5.1.
Caspase-3 activity assay. CE81T/VGH cells (7×106) treated with or without CAS were seeded onto 10-cm dishes and serum starved overnight. Cell lysates were then prepared and lysate proteins (50 μg) were subjected to the analysis by Caspase 3 Assay Kit, Colorimetric (CASP3C, Sigma-Aldrich) in a triplicated manner as instructed. The reaction for color development was held at 37°C for 90 min, and the value of OD405 was determined.
Determination of synergistic effect. Determination of the synergistic interactions between autophagy inhibitor (AI, i.e., 3-MA or CQ) and DDP followed the analysis as described in our previous study (17). In brief, the referencing function was f(u)1,2=f(u)1×f(u)2; where f(u)1=the fraction unaffected by AI, f(u)2=the fraction unaffected by DDP, and f(u)1,2=the fraction unaffected by AI and DDP. If the interaction is additive, f(u)1,2 will be equal to the product of f(u)1×f(u)2. If the average f(u)1,2 is significantly lower than the average product of f(u)1×f(u)2, the interaction between AI and DDP will be considered as synergistic.
Statistical analyses. Two groups of data presented as mean±SD, including average LC3-II/β-actin ratios from western blot, OD450 values of XTT assays, tumor volumes developed in mice, and OD405 values of caspase-3 activity assay, were analyzed by one-way analysis of variance (ANOVA) followed by the Tukey Multiple Comparison Test using IBM SPSS Statistics 20.0.0 software. A value of p<0.05 was regarded as statistically significant.
Results
CAS increased the expression of autophagy-related genes and resistance against SF and hypoxia conditions of CE81T/VGH cells. We have shown that oral carcinoma OECM-1, leukemia Jurkat T, lymphoma U937, multiple myeloma RPMI8226, and tongue carcinoma SCC-15 cells expressed higher levels of Atg4B and/or LC3-II proteins in serum-deprived medium after long-term treatment with ANE or ANE 30-100K. These treated cells exhibited higher resistance against cisplatin treatment or serum withdrawal and both increased resistance could be reverted by 3-MA and CQ (15, 16). These in vitro studies suggested that tumor cells exposed to CAS might become more resistant to environmental stress through up-regulated autophagy.
To assess the role of CAS for tumor growth in vivo, CE81T/VGH cells and nude mice were chosen as experimental models. We previously showed that this esophageal carcinoma cell line was able to develop tumors in mice with severe combined immunodeficiency and others demonstrated its tumor forming ability in nude mice (18, 19). Before animal studies, we firstly analyzed the effects of CAS on CE81T/VGH cells in vitro.
In this study, CAS was used in the culture of cells with ANE 30-100K (1.75 μg/ml) for 3 weeks. Herein, we further demonstrated that CAS significantly increased protein levels of LC3-II and beclin 1 in CE81T/VGH cells cultured in SF medium than untreated cells (Figure 1A). Furthermore, CAS-treated CE81T/VGH cells also exhibited better viability in SF environment, as revealed by the XTT assay, which could be significantly attenuated by either 3-MA or CQ (Figure 1B). Meanwhile, these cells showed stronger tolerance against hypoxia conditions, which was mitigated by 3-MA (Figure 1C) and CQ (Figure 1D). Collectively, these in vitro data suggested the capability of CAS to provide growth advantage for tumor cells in vitro by up-regulating their background autophagy activities under nutrient-limited conditions.
CAS promoted the growth of CE81T/VGH cells in nude mice in an autophagy-dependent manner. CE81T/VGH cells subjected to chronic ANE (10 μg/ml) stimulation for 3 weeks or to CAS were subcutaneously injected into the hind flank of each nude mouse (n=6). The results showed that both ANE- and CAS-treated cells developed larger tumors in the fourth week compared to untreated control cells and the promoting effect of CAS on tumor growth was more profound and statistically significant in the fifth week (Figure 2A). This experiment has been repeated with similar results.
We next analyzed the effects of CQ, 3-MA, and/or DDP on the growth of CAS-treated cells in mice. These drugs were applied one week after tumor inoculation. We observed that CQ alone significantly reduced tumor sizes than those treated with phosphate-buffered saline (PBS) (Figure 2B). Although DDP alone only achieved partial anti-tumor effect, the combined treatment of CQ+DDP exhibited synergistic activity in reducing tumor size. Similarly, treatment with 3-MA alone achieved significant therapeutic benefit and 3-MA+DDP also attenuated tumor growth in a synergistic manner (Figure 2C).
Because autophagy inhibition has been shown to activate the apoptotic pathway (20), we analyzed caspase-3 activity in cells subjected to single or combined drug treatment. The data showed that DDP, but not 3-MA activated caspase-3 activity in CAS-treated CE81T/VGH cells; however, 3-MA significantly increased DDP-induced caspase-3 activation (Figure 2D). Similarly, CQ alone had no effect on caspase-3, but triggered a significant elevation of DDP-induced caspase-3 activity (Figure 2E).
Discussion
We observed the elevation of LC3-II and Atg4B levels in Jurkat T, OECM-1, U937, RPMI8226, and/or SCC15 cells (15, 16), as well as that of LC3-II and beclin 1 in CE81T/VGH cells (Figure 1A), under SF conditions. These results suggest that CAS may boost autophagy activity in different types of tumors in nutrient-limited locale. Furthermore, increased resistance against serum depletion was demonstrated in CAS-treated U937, RPMI8226, and SCC15 cells (15), as well as in CAS-treated CE81T/VGH cells (Figure 1B). Together with the stronger tolerance of CAS-treated CE81T/VGH cells against hypoxia conditions (Figure 1C and D), CAS is thought to offer tumor cells increased adaptation to nutrient-limited environment to promote tumor growth through up-regulation of their autophagy status. We verified, for the first time, that CAS can facilitate tumor growth in vivo (Figure 2A), which is sensitive to autophay inhibition by 3-MA or CQ (Figure 2B and 2C). This feature may emphasize autophagy inhibition as the potential strategy for treating tumors survived from CAS.
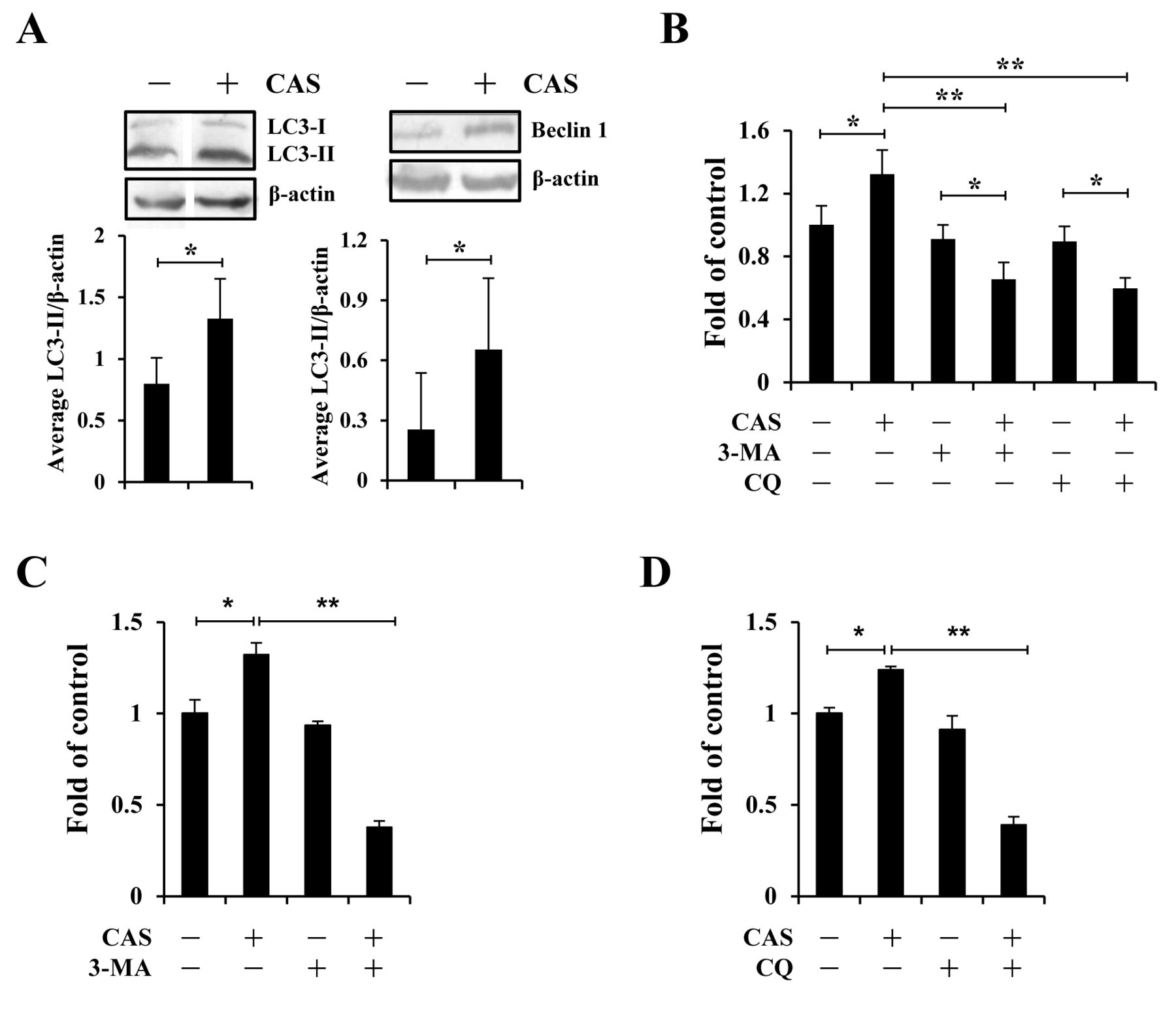
CAS increased the tolerance of CE81T/VGH cells against serum-free and hypoxia conditions through up-regulated autophagy. (A) CE81T/VGH cells treated with (+) or without (−) CAS were serum starved for 24 h. LC3 and β-actin (left) and beclin 1 and β-actin (right) proteins in these two cells were then analyzed by western blots. Average LC3-II/β-actin ratios (±SD) from three independent experiments were plotted under each lane. (B) Alternatively, during serum starvation, these two cell lines were treated with or without 3-MA (1 mM) and CQ (25 μM) for 24 h, followed by XTT assays for viability analysis. Average OD450 values (±SD) from triplicated wells of each treatment against that of control were plotted. Cells cultured in 0.1% O2 conditions for 24 h in the presence of 3-MA (1 mM) (C) or CQ (25 μM) (D) were subjected to XTT assays. Average OD450 values (±SD) from triplicated wells were presented as (B). *p<0.05, **p<0.01.
The tumor microenvironment is known to be deficient in nutrients and oxygen (21). Our previous and current studies have shown the growth advantage provided by CAS for six distinct cell lines (OECM-1, Jurkat T, U937, RPMI8226, SCC-15, and CE81T/VGH) against SF challenge. We also noticed the increased survival rates of CAS-treated OECM-1 (16) and CE81T/VGH (Figure 1C and D) cells under hypoxia conditions. In the meantime, all these CAS-induced increase of cell viability under SF and hypoxia conditions was sensitive to either 3-MA or CQ. Therefore, CAS may help tumor cells to survive hostile microenvironment through upregulation of tumoral autophagy.
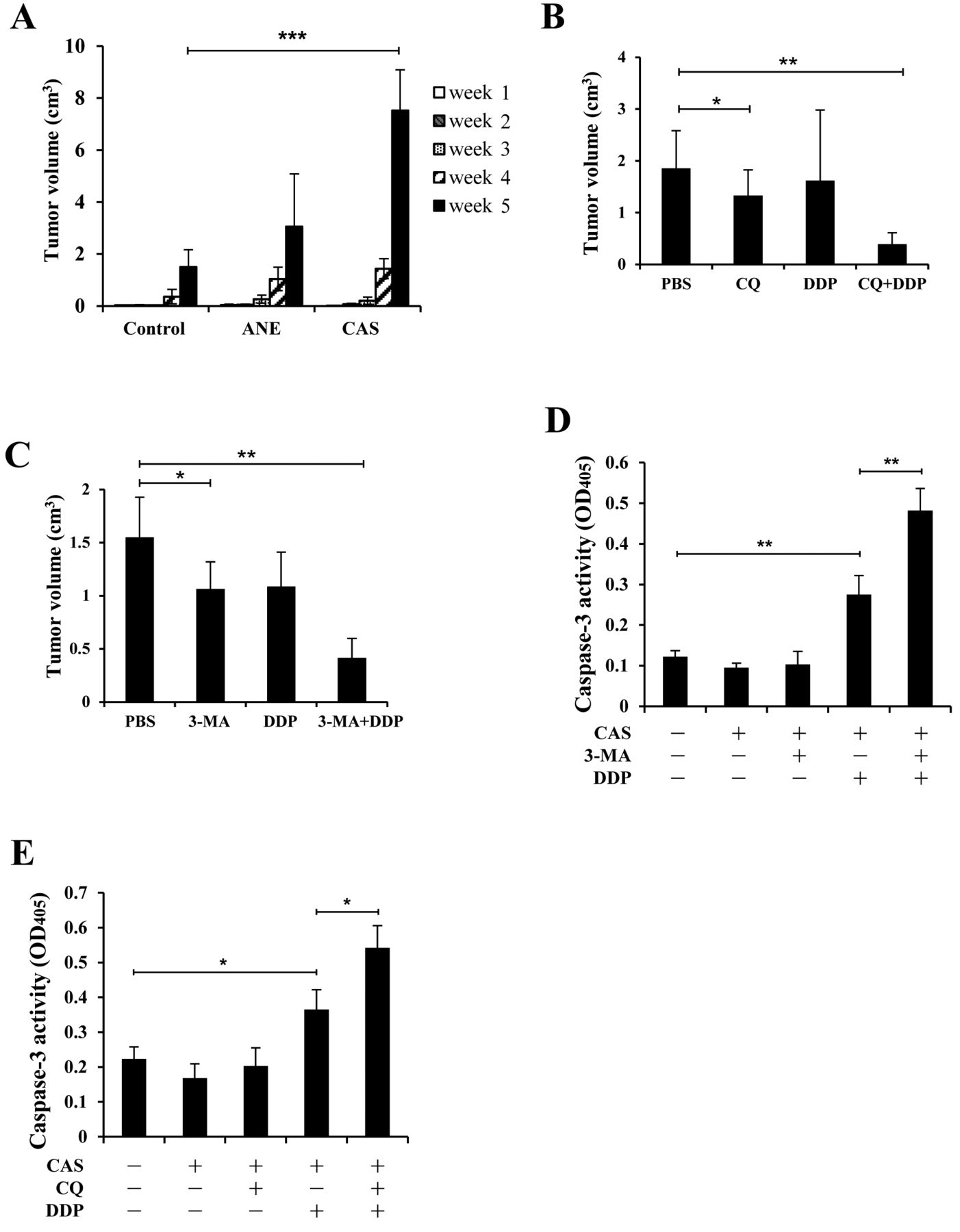
CAS promoted the growth of CE81T/VGH cells in nude mice in an autophagy-dependent manner. (A) Untreated (control), ANE-treated (ANE), and CAS-treated (CAS) CE81T/VGH cells were injected subcutaneously into mice (n=6). Tumor volumes were determined weekly and the average tumor volume of each treatment (±SD) was plotted. (B) CAS-treated cells were identically inoculated as (A) and grown in mice for a week, followed by treatment with CQ (60 mg/kg/day), DDP (5 mg/kg/6 days), or both (CQ+DDP). The average tumor volumes (±SD) were plotted as (A). (C) The same experiment was performed as (B) except that CQ was substituted by 3-MA (30 mg/kg/day). CAS-treated CE81T/VGH cells cultured in SF medium for 24 h in the presence of 3-MA (1 mM) or DDP (21.5 μM) (D), or in the presence of CQ (25 μM) and/or DDP (21.5 μM) (E), were subjected to caspase-3 activity assay. Average OD405 values (±SD) from triplicated wells of each treatment were plotted. *p<0.05, **p<0.01.
Autophagy is currently regarded as being tumor-promotive in most malignant cells (2). However, the suppressive role of autophagy for tumor growth cannot be ignored because autophagy inhibitors (Lys05 and SBI-0206965) and inducers (rapamycin and obatoclax) were both shown to exhibit anti-tumor activities (22-25). In addition to the individual difference among different cancer cells, tumors are known to be heterogenic (26). It is, thus, possible that the dependence of cells on autophagy varies among distinct tumors and among different regions of the same tumor. This feature may lead to autophagy modulation-based therapy being unpredictable.
Single treatment with autophagy inhibitors like 3-MA and CQ received differential anti-tumor results in mice. For example, 3-MA exhibited a mild therapeutic effect on mice carrying RITA-resistant head and neck cancer cells, but not on those bearing salivary adenoid cystic carcinoma ACC-M cells (27, 28). Similar therapy was also observed in CQ-treated bladder cancer J82 cells and hypopharyngeal carcinoma FaDu cells, respectively (29, 30). Our in vitro and in vivo investigations suggest that CAS may serve as a driving force for more tumor populations to become autophagy addicted to overcome the inhospitable microenvironment and more sensitive to autophagy inhibition.
CQ and its derivative hydroxychloroquine are currently approved drugs for autophagy inhibition by FDA; however, their effectiveness and specificity have been questioned (7). Fortunately, there are accumulating FDA-approved drugs, such as verteporin, clomipramine, and desmethylclomipramine, found to block autophagy through altering the acidification of lysosome or preventing the fusion of autophagosome and lysosome by new screening technology (31). As autophagy inhibition is a new attractive direction for cancer therapy, our current studies further highlight the possible benefits to include autophagy inhibitor alone or in combination with DDP for treating tumors encountered with CAS.
Acknowledgements
This study was supported by the Ministry of Science and Technology (MOST 104-2314-B-366 -001-MY3), Chi Mei Medical Center (CMFH10706 and 10805), and An Nan Hospital, China Medical University (ANHRF107-2).
Footnotes
↵* These Authors contributed equally to this study.
Author's Contributions
Y-C.L. and M-H.L. conceived and designed the experiments, B-Y.L., Z-Y.S., C-Y.W., and J-L.D. performed experiments, Y-C.L. analyzed data, and wrote the manuscript. C-T.C. and M-H.L. critically reviewed the manuscript. C-T.C., S-Y.L, and C-Y.Y. provided infrastructure and research facility to performed aforementioned experiments.
Conflicts of Interest
The Authors declare no conflicts of interest.
- Received November 14, 2019.
- Revision received November 21, 2019.
- Accepted November 25, 2019.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.