


Abstract
Background/Aim: The aim of this study was to examine whether the Wnt/β-catenin signal activation is a cause of radioresistance in colon cancer by assessing the β-catenin localization and its correlation with cancer stem cells (CSCs). Materials and Methods: The nuclear levels of β-catenin, the hallmark of Wnt activation, were analyzed in HCT116 and SW480 cells by immunohistochemistry, before and after irradiation. Further, we assessed CSC populations by staining for aldehyde dehydrogenase-1 (ALDH1) and CD44. Results: β-catenin was localized predominantly in the nucleus and plasma membrane in SW480 and HCT116 cells, respectively. Compared to HCT116 cells, SW480 cells displayed higher Wnt activation. At 24 h after irradiation, most of the DSBs in SW480 cells were repaired, but were still present in HCT116 cells. Additionally, compared to HCT116 cells, a significantly higher proportion of SW480 cells were ALDH1- and CD44-positive. Conclusion: Colon cancers with nuclear β-catenin accumulation demonstrated greater radio-resistance with a higher number of CSCs.
There are two Wnt/β-catenin signaling pathways: the canonical pathway and the non-canonical pathway (1, 2). β-catenin is the key molecule in the canonical Wnt pathway. In Wnt-off state, β-catenin in the cytoplasm is within a protein complex composed of adenomatous polyposis coli (APC), Axin, glycogen synthase kinase 3β (GSK3β), and casein kinase 1α (CK1α), and phosphorylated by GSK3β and CK1α. Then, β-catenin is ubiquitinated and targeted for destruction by the proteasome. As a result, the amount of β-catenin is low. Conversely, in the Wnt-on state, Wnt ligands bind to the receptor complex composed of a Frizzled (Fz) receptor and low-density lipoprotein receptor-related proteins (LRP5/6). This receptor recruits Disheveled (Dvl) to the plasma membrane. Dvl forms a complex with Axin, Fz, and LRP5/6 and acts as a direct competitive inhibitor of GSK3β. Thus, β-catenin escapes phosphorylation and accumulates in the cytoplasm. The stabilized β-catenin translocates into the nucleus, where it serves as a transcriptional co-activator and influences gene transcription. Thus, β-catenin localization to the nucleus reflects Wnt activity (3-5).
The canonical Wnt/β-catenin signaling pathway plays an important role in cell proliferation, growth, and differentiation (6, 7). Mutations in Wnt signaling pathway components cause many diseases, including cancer. Furthermore, dysregulation of the Wnt signaling pathway is associated with the expansion of stem cell and progenitor cell lineages and is also highly associated with carcinogenesis (8-10). Most colorectal cancer (CRC) cases are caused by mutations in Wnt signaling pathway components (11-13). Originally, a mutation in the APC gene was associated with familial adenomatous polyposis (FAP). Approximately 1% of FAP cases progress to CRC (14, 15). Furthermore, 85% of sporadic intestinal neoplasia cases have mutations in APC, whereas activating mutations in β-catenin are found in approximately 50% of CRC tumors without APC mutations (16, 17). The Wnt signaling pathway is related to both carcinogenesis and treatment resistance (18). Several reports have evaluated the correlation between the Wnt signaling pathway and resistance to chemotherapy (19, 20). Several attempts to discover new drugs targeting the Wnt signaling pathway and to improve chemo-resistance have been made (21-23). Although the relationship between the Wnt signaling pathway and chemoresistance has been well studied, their relationship with radioresistance remains unclear.
The Wnt signaling pathway is responsible for designating cancer stem cells (CSCs) (24). The nuclear localization and subsequent activation of T cell factor (TCF) transcription is an important process in CSCs (25). CD44 is a multifunctional cell surface adhesion protein that reacts with extracellular matrix components. Increased CD44 expression leads to enhanced CSC properties in colon cancer cells (26, 27). There are also reports showing that the combination of CD44 and CD133 is correlated with features of CSCs (28). CD44 helps to facilitate cell-cell and cell-matrix interactions through its affinity for hyaluronic acid; it is also involved in cell adhesion and the assembly of growth factors on the cell surface (29). Increased resistance to radiation therapy has been reported in CD44high cells compared to CD44low cells (29-31).
The aldehyde dehydrogenase (ALDH) family of enzymes are cytosolic isoenzymes responsible for oxidizing intracellular aldehydes and retinol to retinoic acid in early stem cell differentiation (32). ALDH1 is expressed in various normal stem and progenitor cells and contributes to cell proliferation and differentiation (27, 33). Some reports have shown that nuclear localization of ALDH1A1 is associated with a dismal prognosis in clinical CRC cases (34, 35). ALDH1 is also correlated with radioresistance (31, 36, 37).
The epithelial cell adhesion molecule (EpCAM) is also known as an epithelial-specific antigen. EpCAM was initially discovered as a predominant antigen in human colon carcinoma; it is a transmembrane glycoprotein that is highly expressed in rapidly proliferating carcinomas (38). Recent data have suggested a more multifaceted role of EpCAM in not only cell-cell adhesion but also cell signaling, migration, proliferation, and differentiation (39). EpCAM is also reported to be a putative CSC marker (40). EpCAM expression is also correlated with radioresistance (41).
This study focused on the localization of β-catenin and analyzed its correlation with radioresistance and cancer stem-like cells.
Materials and Methods
Cell culture. Two human CRC cell lines, HCT116 and SW480 (American Type Culture Collection, Manassas, VA, USA), were maintained in culture using standard protocols. The HCT116 cell line was cultured in McCoy's 5A medium (American Type Culture Collection, Manassas, VA, USA) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin. The SW480 cell line was cultured in Leibovitz's L-15 medium (American Type Culture Collection, Manassas, VA, USA) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin.
Irradiation. Photon irradiation was delivered by a Primus system (Siemens Healthcare, Malvern, PA, USA). The photon energy was 4 MV and the dose rate was 250 monitor units per min. Cells were plated into dishes at approximately 50% confluency. Twenty-four h after plating, a 1-cm bolus was put on dishes and the cells in dishes were irradiated.
Clonogenic assay. Each cell line was irradiated in dishes with 0 (control), 2, 4, and 8 Gy, respectively. Subsequently, 100 and 1,000 cells were re-plated. After 14 days of cultivation, the cells were rinsed with phosphate buffer saline. A 3-ml of 6.0% glutaraldehyde and 0.5% crystal violet was then added and incubated for 30 min. The glutaraldehyde crystal violet mixture was then rinsed with water and the plate was allowed to dry in normal air at room temperature. Finally, the colonies were counted.
Cell block. The cell samples were fixed for one h by mixing with 10% buffered formalin. After one h, the samples were centrifuged at 1000 x g for 15 min. The supernatant was discarded and a further fresh 100% and then 70% alcohol was added to the sediment and incubated for one day. On the following day, the sediment containing the cells was processed along with other routine histopathological specimens.
IHC staining. A labeled streptavidin-biotin (LSAB) method was used for immunohistochemical (IHC) staining of deparaffinized 3-μm-thick sections. The sections were placed in citrate buffer (10 mmol-1, pH 6.0) and then autoclaved at a chamber temperature of 121°C for 1 min to retrieve the antigen. They were then rinsed and blocked in 3% hydrogen peroxide. Non-specific binding sites were blocked in 0.01 M phosphate-buffered saline (pH 7.4) containing 2% bovine serum albumin (Wako Pure Chemical, Osaka, Japan) for 40 min. The primary antibodies used were purified mouse anti-β-catenin [mouse immunoglobulin (IgG), 1:100, BD bioscience, CA, USA], anti-gamma H2A.X (phospho S139) antibody (rabbit IgG, 1:100, abcam, CA, UK), anti-ALDH1A1 antibody (ab52492) (rabbit IgG, 1:500, abcam, CA, UK), purified mouse anti-human CD44 (550392) (mouse IgG, 1:25, BD Pharmingen, NJ, USA), and anti-EpCAM antibody (ab71916) (rabbit IgG, 1:50, abcam, Cambridge, UK).
Results
Localization of β-catenin in colorectal cancer (CRC) cell lines. To investigate the localization of β-catenin, the hallmark of Wnt activation, in CRC cell lines, we performed IHC with β-catenin antibody in SW480 and HCT116 CRC cell lines, which have similar protein expression levels in western blot analysis (42, 43). HCT116 cells express wild-type APC and mutant β-catenin, while SW480 cells express mutant APC. SW480 cells had one mutant allele with loss of the other allele (44). β-catenin expression was localized in the nucleus and cytoplasm of SW480 cells and in the membrane of HCT116 cells (Figure 1). The results indicated that SW480 cells had a different Wnt/β-catenin signaling pathway activation compared to HCT116 cells.

Expression of β-catenin in HCT116 and SW480 cells. In SW480 cells, β-catenin predominantly localized in the nucleus, whereas in HCT116 cells, it was primarily found in the plasma membrane. Arrow indicates cellular membrane. Arrowhead indicates nucleus. Scale bars: 50 μm.
Comparison of DNA repair rates after irradiation. To assess the level of DNA double-strand break (DSB) repair after irradiation, expression of γH2AX, a surrogate marker of DSBs, was evaluated in both SW480 and HCT116 cell lines after 4 Gy irradiation. The number of foci was significantly lower in SW480 cells than that in HCT116 cells at 24 h after irradiation (Figure 2). These results suggested that nuclear β-catenin, the hallmark of Wnt activation, is associated with DSB repair at 24 h after irradiation.
Sensitivity of CRC cells to X-rays assessed by clonogenic assay. To measure the survival of both SW480 and HCT116 cells after irradiation, clonogenic assays were performed following 0, 2, 4, and 8 Gy irradiation (Figure 3). Fourteen days after irradiation, colonies on each dish were counted. At each irradiation dose, the ratio of colony numbers to the control group (0 Gy) was calculated and expressed logarithmically. Figure 3 shows the results of clonogenic assays starting from 1,000 cells. There were significantly more colonies in SW480 cells than in HCT116 cells at 2, 4, and 8 Gy (p=0.0316, <0.0001, and 0.0002, respectively). Similarly, starting from 100 cells, the clonogenic assay showed significantly higher numbers of colonies in SW480 cells than those in HCT116 cells at 2, 4, and 8 Gy (data not shown). No colony formation was observed in the dishes where 100 HCT116 cells plated after 8 Gy irradiation. These results indicate that SW480 cells were more radioresistant compared to HCT116 cells. Taken together, Wnt-activated CRC cells may be radioresistant relatively to non-activated CRC cells.
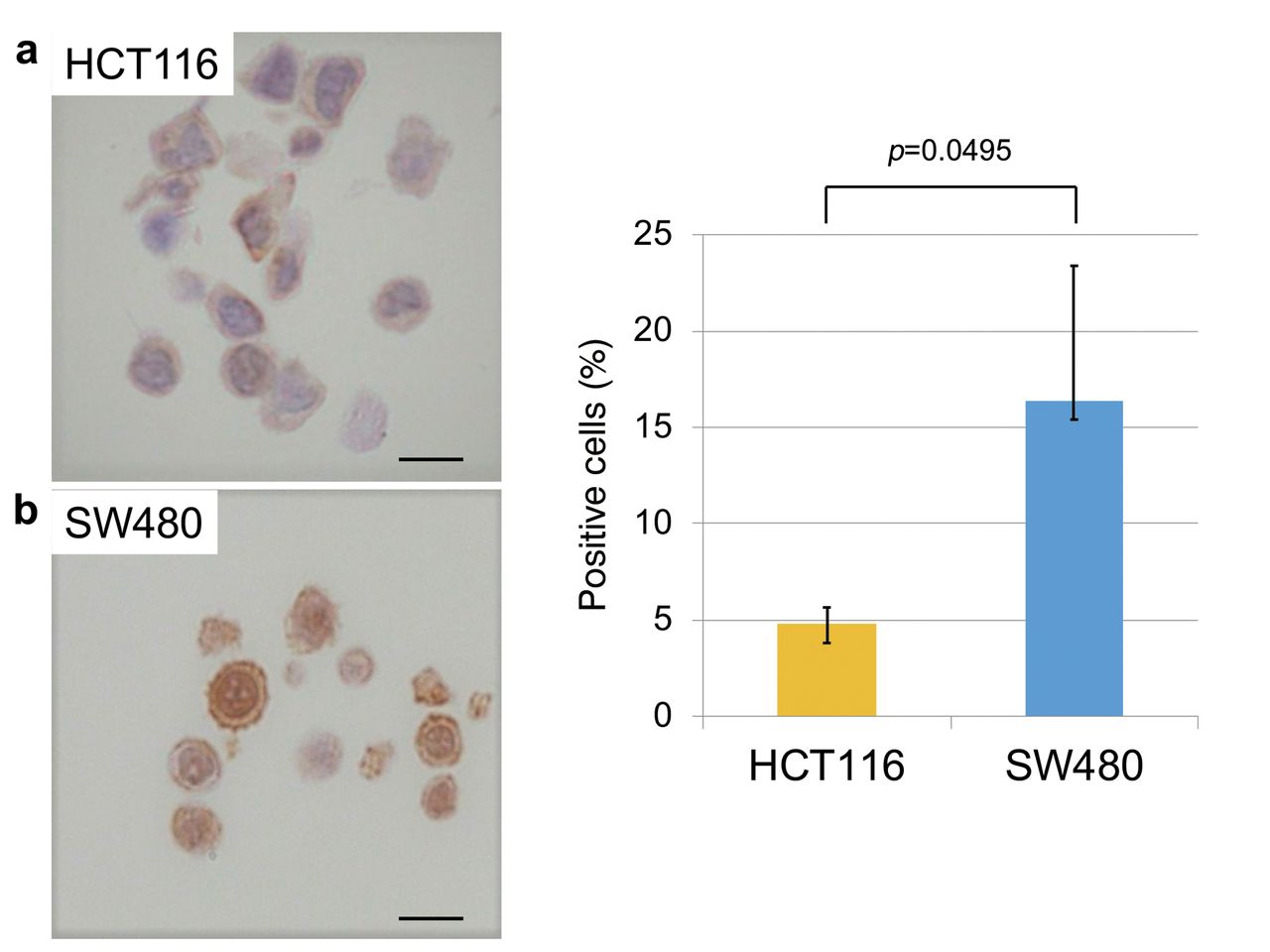
Expression of CSC markers in Wnt-activated and non-activated CRC cells. Next, to evaluate the correlation between putative CSC markers and nuclear β-catenin, IHC for ALDH1, CD44, and EpCAM was performed in each cell line. SW480 cells, which have high β-catenin accumulation in the nucleus, had a significantly higher proportion of ALDH1-positive cells compared to HCT116 cells, which showed almost exclusive membranous staining of β-catenin (Figure 4). Similar to ALDH1 expression, the proportion of CD44-positive SW480 cells was also significantly higher than that in HCT116 cells (Figure 5). In contrast, the proportion of EpCAM-positive SW480 cells was significantly lower than that in HCT116 cells (Figure 6). These results suggest that ALDH1 and CD44 expression may be radioresistant markers as well as nuclear β-catenin expression in CRC cells.
Discussion
When the Wnt/β-catenin signaling pathway is active, β-catenin is not degraded; it accumulates and translocates into the nucleus. Thus, increased β-catenin levels in the nucleus indicate Wnt-active status. In contrast, when Wnt is inactive, β-catenin is degraded by the proteasome and the levels of β-catenin in the cytoplasm are decreased. Translocation into the nucleus does not occur because the cytoplasmic β-catenin level is not high. Therefore, higher β-catenin levels in the plasma membrane rather than the nucleus indicates inactive Wnt. In SW480 cells, β-catenin was predominantly observed in the nucleus, whereas it was primarily observed in the plasma membrane in HCT116 cells. SW480 cells showed higher Wnt/β-catenin signaling pathway activation compared to HCT116 cells.

γH2AX expression after irradiation. (a) γH2AX foci after 4 Gy irradiation in HCT116 and SW480 cells. At 24 h after irradiation, most of the double-strand breaks (DSBs) in the SW480 cells are repaired, whereas they remain in the HCT116 cells. (b) The number of γH2AX foci in SW480 cells was significantly lower than that in HCT116 cells at 24 h after irradiation (Student's t-test; N=10). Scale bars: 10 μm.
Photon beams have an anti-tumor effect by damaging tumor DNA. The DSB levels were assessed via γH2AX, a surrogate marker of DSBs. At the early time point, there was no difference in DNA damage foci formation between HCT116 and SW480 cells. However, 24 h after irradiation, most of the DSBs in SW480 cells were repaired, but remained in HCT116 cells. The number of γH2AX foci in SW480 cells was significantly lower than that in HCT116 cells at 24 h after irradiation. Wnt signaling is related to DNA damage repair, suggesting increased repair with high Wnt activity (45). This is considered to be one of the causes of resistance to treatment. Moreover, the results of the colony formation assay showed that SW480 cells, with higher Wnt activity, formed higher number of colonies and showed resistance to irradiation compared to HCT116 cells, with lower Wnt activity. This result was similar regardless of the irradiation dose and numbers of seeding cells.
Wnt signaling is also involved in CSC control (46-48). CD44, CD133, ALDH1, Lgr5, EpCAM, etc. are putative markers of CSC in colorectal cancer (26-28, 33, 35, 40, 49). Among them, we evaluated CD44, ALDH1, and EpCAM by IHC. Koh et al. have reported that activation of gastrin by β-catenin is an early event of colon carcinogenesis that promotes tumor development (50). Wnt signaling is enhanced by the secretion of progastrin, a direct target of the oncogenic Tcf/β-catenin pathway (51, 52). Jin et al. have reported an increased proportion of CD44-positive cells following the addition of prosgastrin in a colon cancer cell line, indicating an increase in putative CSC (53). In addition, Giraud et al. have reported that decreased proportion of ALDHhigh cells by suppression of progastrin resulted in decreased tumorigenesis and glycolytic ability and restricted the self-renewal ability of CSC (54). Thus, there may be a relationship between the Wnt signaling and CSC. In this study, SW480 cells with high Wnt activity had a significantly higher proportion of CD44-positive cells than HCT116 cells. A similar result was observed for ALDH1. SW480 cells with high Wnt activity had a significantly lower proportion of EpCAM-positive cells than HCT116 cells. While there are reports that EpCAM may be a marker of CSC, this finding remains controversial. EpCAM expression has been linked to poorer disease-free and overall survival in breast cancer (55). Gosens et al. have observed strong EpCAM staining in the tumor center and a progressive loss at the tumor front associated with poor local and overall recurrence-free survival in colorectal cancer (56). Lugli et al. have reported a correlation between the loss of membranous EpCAM and tumor invasiveness and progression (57). EpCAM overexpression was related to lymph node classification and tumor border configuration (pushing or infiltration) but not to local recurrence. These results suggest that the loss of EpCAM at the tumor border results in reduced adhesion and induces the epithelial-mesenchymal transition, resulting in lymph node and distant metastasis. Pastushenko et al. have analyzed cells that did not express EpCAM and isolated cell populations based on their expression of the mesenchymal-cell receptor proteins CD51, CD61 and CD106. The authors identified two cell populations that were the most likely to metastasize; EpCAM(−), CD106(−), CD51(−), CD61(−) and EpCAM(−), CD106(+), CD51(−), CD61(−) (58, 59).These findings suggest that EpCAM affects metastasis and prognosis but has little direct effect on radiosensitivity.

Colony formation after irradiation in SW480 and HCT116 cells. (a) Colony formation assay in HCT116 and SW480 cells. (b) Comparison of radioresistance between SW480 and HCT116 cells (Student's t-test; N=3). Scale bars: 10 μm.
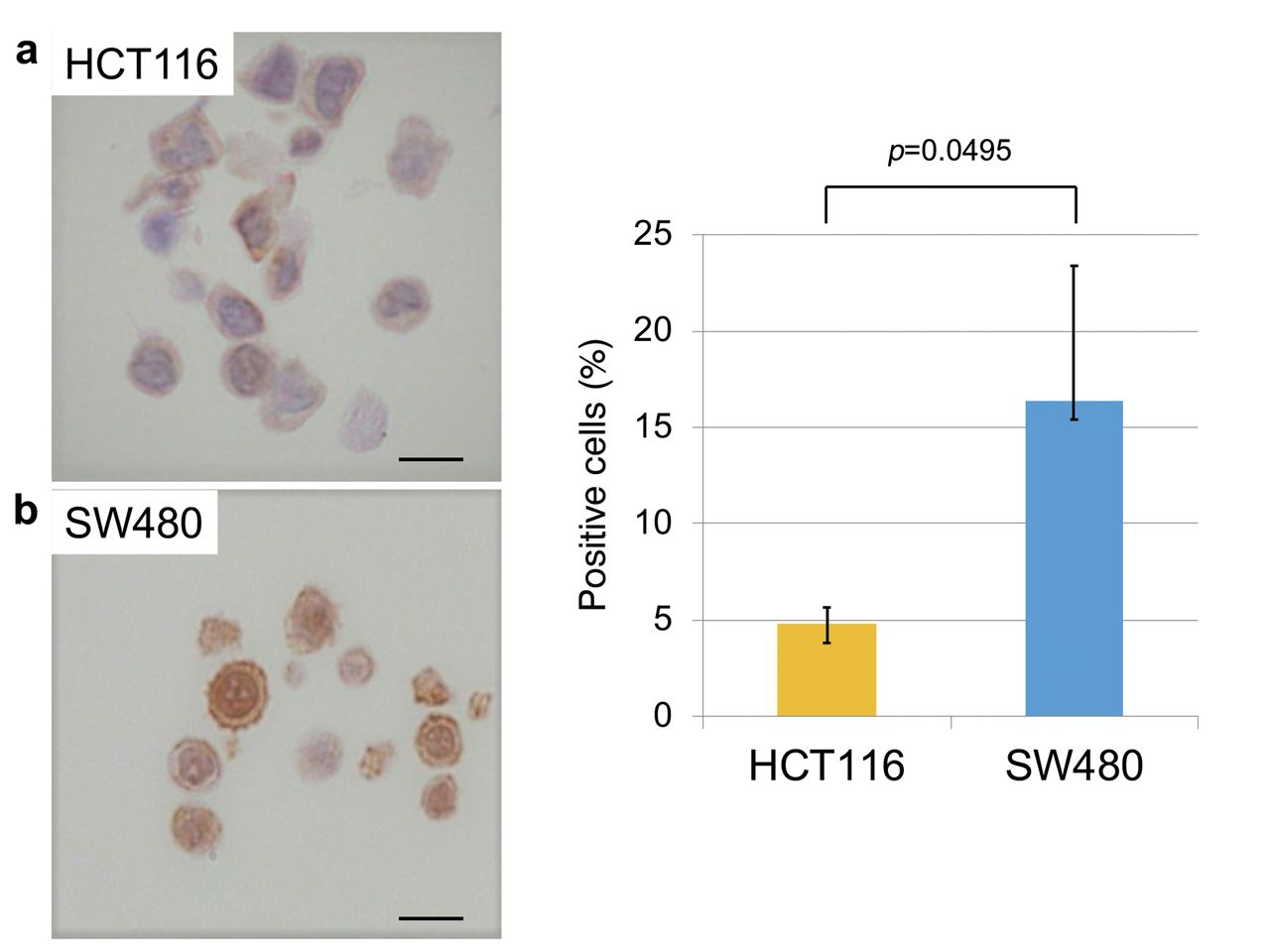
ALDH1 expression in HTC116 and SW480 cells. (a) Expression of ALDH1 in HTC116 and SW480 cells. (b) SW480 cells had a significantly higher proportion of ALDH1-positive cells compared to HCT116 cells (Student's t-test; N=5). Error bars represent the standard deviation of the mean value. Scale bars: 10 μm.
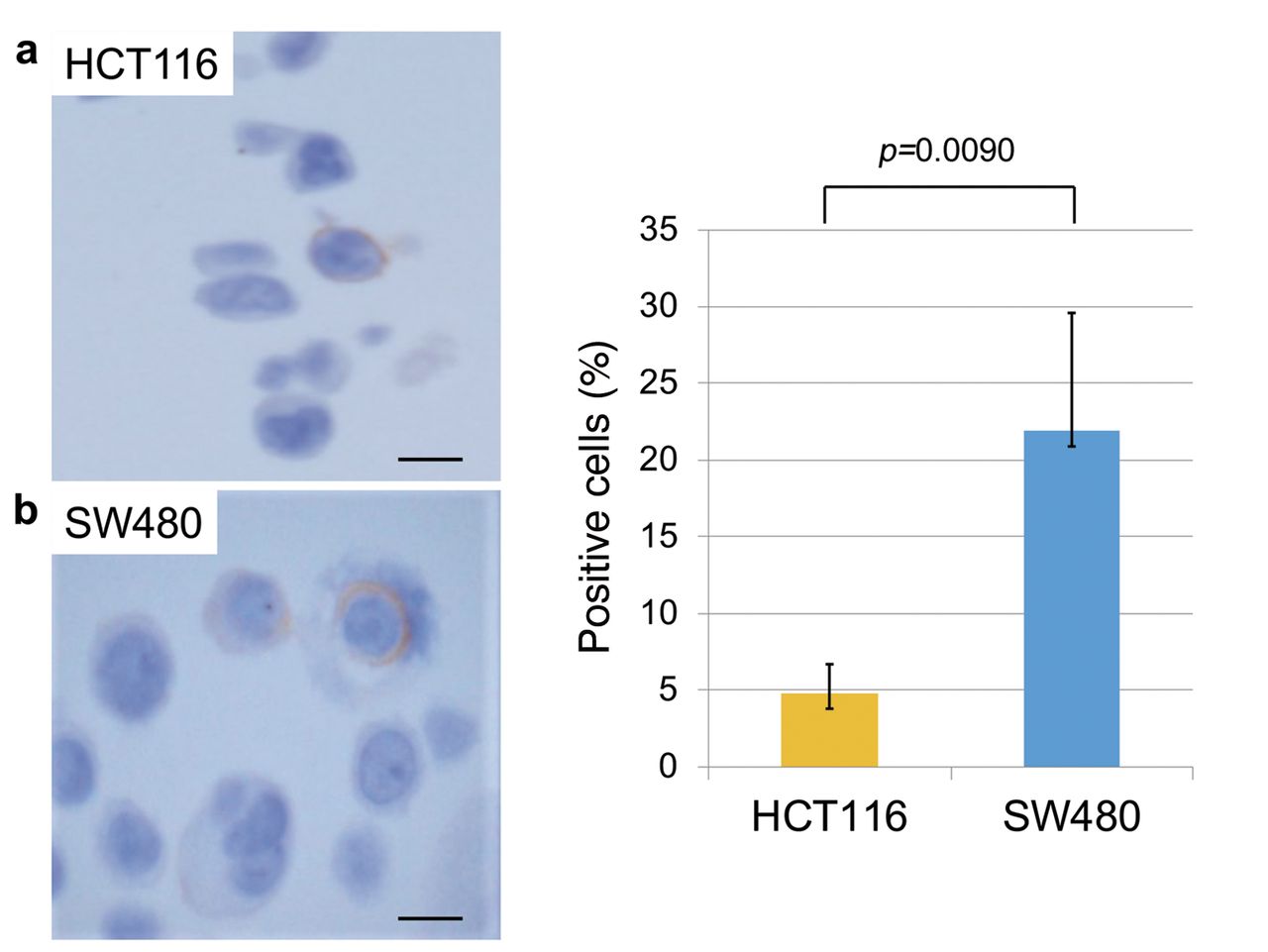
CD44 expression in HTC116 and SW480 cells. (a) Expression of CD44 in HTC116 and SW480 cells. (b) SW480 cells had a significantly higher proportion of CD44-positive cells compared to HCT116 cells (Student's t-test; N=5). Error bars represent the standard deviation of the mean value. Scale bars: 10 μm.

EpCAM expression in HCT116 and SW480 cells. Expression of EpCAM in HCT116 and SW480 cells. (b) HCT116 cells had a significantly higher proportion of EpCAM-positive cells compared to SW480 cells (Student's t-test; N=5). Error bars represent the standard deviation of the mean value. Scale bars: 10 μm.
In conclusion, nuclear accumulation of β-catenin is an indicator of active Wnt signaling. Cell lines with higher nuclear β-catenin accumulation showed resistance to radiation therapy compared to cell lines with lower accumulation, as well as a significantly higher positive proportion of putative CSC. These findings suggest that the Wnt/β-catenin signaling pathway is related to radioresistance and stemness of CSC.
Acknowledgements
The paper has been edited and rewritten by an experienced scientific editor, who has improved its grammar and stylistic expression. The Authors thank Enago, Crimson Interactive Pvt, Ltd.
Footnotes
↵* These Authors contributed equally to this study.
Authors' Contributions
HTan, FH, HTom, Tmor, AH, and MM conceived and designed the experiments. HTan, MK, Tmiy, FH, Htom, Tmor, AH, and MM analyzed the data. HTan, MK, SS, RI, Htom, and Tmor wrote the first draft of the manuscript. HTan, MK, SS, Tmiy, RI, FH, Htom, Tmor, AH, and MM contributed to the writing of the manuscript. HTan, MK, SS, Tmiy, RI, FH, Htom, Tmor, AH, and MM agreed with the manuscript results and conclusions. HTan, MK, SS, Tmiy, RI, FH, Htom, Tmor, AH, and MM jointly developed the structure and arguments for the paper. HTan, MK, SS, Tmiy, RI, FH, Htom, Tmor, AH, and MM made critical revisions and approved the final version. HTan, MK, SS, Tmiy, RI, FH, Htom, Tmor, AH, and MM reviewed and approved the final manuscript.
Conflicts of Interest
The Authors declare that they have no competing interests regarding this study.
- Received September 29, 2019.
- Revision received November 16, 2019.
- Accepted November 18, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}