


Abstract
Background/Aim: Natural killer (NK) cells are considered potential antitumor effector cells. The aim of this study was to establish a novel type of a chimeric antigen receptor (CAR) NK cell line (CAR-KHYG-1) specific for epidermal growth factor receptor variant III (EGFRvIII)-expressing tumors and investigate the anti-tumor activity of EGFRvIII-specific-CAR-KHYG-1 (EvCAR-KHYG-1). Materials and Methods: EvCAR-KHYG-1 was established by self-inactivated lentiviral-based transduction of the EvCAR gene and magnetic bead-based purification of EvCAR-expressing NK cells. The anti-tumor effects of EvCAR-KHYG-1 were evaluated using growth inhibition and apoptosis detection assays in glioblastoma (GBM) cell lines (EGFRvIII-expressing and non-expressing U87MG). Results: The findings demonstrated that EvCAR-KHYG-1 inhibited GBM cell-growth via apoptosis in an EGFRvIII-expressing specific manner. Conclusion: This is the first study to establish a CAR NK cell line based on the human NK cell line KHYG-1. Therapy with EvCAR-KHYG-1 may be an effective treatment option for GBM patients.
- Chimeric antigen receptor
- natural killer cells
- glioblastoma
- epidermal growth factor receptor variant III
Glioblastoma (GBM) is one of the most common and aggressive primary malignant brain tumors occurring in adults (1-3). Even after undergoing chemotherapy, radiation, and/or surgical resection, few patients survive more than 5 years (2-4). Thus, new therapeutic strategies are needed. Immune-based therapy for GBM is a promising alternative to conventional treatments, with the possible long-term benefit of generating a sustainable anti-tumor response, and the potential to target both localized and infiltrating tumor cells (4-11). Natural killer (NK) cells respond rapidly to transformed and stressed cells, and represent an important effector cell type for adoptive immunotherapy (12, 13). In addition to donor-derived primary NK cells, continuously expanding cytotoxic cell lines, such as NK-92, are under development for clinical applications (14, 15). Additionally, NK cells have advantages over T cells, as they express the Fc fragment of the IgG low affinity III receptor (FcyR III), which binds to the Fc fragment of antibodies to elicit antibody-dependent cell-mediated cytotoxicity. This specific feature of NK cells enables the combination of 2 targeted therapies that recognize different, or the same, tumor-associated antigens, namely chimeric antigen receptor (CAR)-expressing NK cells (CAR NK) and tumor-associated antigen-specific monoclonal antibodies. NK cells spontaneously kill tumor cells by recognizing diverse ligands via a variety of activating receptors, including natural cytotoxicity receptors.
CAR is an artificially modified fusion protein consisting of an extracellular antigen recognition domain fused to an intracellular signaling domain (4, 16), and redirects the specificity and function of T lymphocytes and other immune cells. The general premise for the use of CAR cells in cancer immunotherapy is the rapid generation of tumor-targeted T cells, thus bypassing the barriers and incremental kinetics of active immunization (17, 18).
CAR T cells (CAR T) have been effectively used for treatment of refractory chronic lymphocytic leukemia and acute lymphoblastic leukemia (19, 20). Specifically, CD19-targeting CAR T have been reported to result in complete response rates of 70% to 90% in patients with ALL (21). However, CAR T exhibit poor therapeutic efficacy when used to treat solid tumors (22-24).
Similar to CAR T, CAR NK have exhibited improved tumor-specific targeting and cytotoxicity against cancer cells in both in vitro and in vivo experiments (25, 26). Importantly, NK cells have many advantages as compared to T cells for use in CAR-targeted immunotherapy (27, 28). For example, allogeneic NK cells can be used as effector cells, since they are not responsible for graft-versus-host disease (GVHD) because they do not require HLA matching. Furthermore, CAR NK may be safer than CAR T, as they usually do not induce cytokine storms, which are seen with CAR T (29, 30). Additionally, NK cells can be generated from a variety of sources, such as peripheral blood, umbilical cord blood, human embryonic stem cells, induced pluripotent stem cells, and even NK cell lines (28).
In the present study, novel lentiviral vectors were constructed for a CAR line consisting of an epidermal growth factor receptor variant III (EGFRvIII)-specific single chain variable fragment (scFv) coupled to the motifs of a CD3ζ chain (T-cell receptor signaling module), and co-stimulatory CD137 (4-1BB) and CD28 (pELNS-3C10-CAR). The human NK cell line KHYG-1 was transduced with these lentiviral vectors and the EGFRvIII specific-CAR-KHYG-1 (EvCAR-KHYG-1) cell line was generated. The anti-tumor effects of this cell line were evaluated in vitro. This is the first study to establish a CAR NK line based on the human NK cell line KHYG-1.
Materials and Methods
Cell lines. Human GBM cell line U87MG and HEK293T cells were purchased from the American Type Culture Collection (ATCC; Manassas, VA, USA). EGFRvIII expressing-U87MG (U87MG-EGFRvIII) cells were kindly provided by Atsushi Natsume of Nagoya University School of Medicine. GBM cells were maintained in Dulbecco's Modified Eagle Medium (DMEM; Thermo Fisher Scientific Inc., Waltham, MA, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; MP Biomedicals, Santa Ana, CA, USA). Human NK cell line KHYG-1 cells were purchased from Japanese Collection of Research Bioresources (JCRB; Ibaraki, Osaka, Japan) and maintained in Roswell Park Memorial Institute 1640 medium (RPMI 1640; Thermo Fisher Scientific Inc.) with 150 international units (IU)/ml of recombinant human Interleukin (rhIL)-2 (Novartis Pharma AG, Basel, Switzerland) supplemented with 20% FBS. Each type of culture medium used in this study was supplemented with 100 units/ml penicillin and 100 μg/ml streptomycin (Thermo Fisher Scientific Inc.). The cell lines were cultured under standard culture conditions at 37°C in a humidified atmosphere containing 5% CO2.
Construction of self-inactivating (SIN) lentiviral vectors. The EGFRvIII-specific CAR carrying-self-inactivating (SIN) lentiviral vector was previously reported (31). mAb 3C10 was originally developed by immunization of mice with a 14-amino acid peptide incorporating an EGFRvIII-specific fusion junction (32), then its single chain variable fragment (scFv) was subsequently cloned (33). The scFv portion in pELNS-SS1CD28r1BBZeta was replaced with the cDNA for 3C10 scFV by gene synthesis (Genscript, Piscataway, NJ) to create pELNS-3C10-CAR.
Production of vesicular stomatitis virus G glycoprotein (VSV-G) pseudotype SIN-lentivirus and transduction of NK cells. Production of SIN-lentivirus was performed as previously described, with slight modifications (31). Briefly, HEK293T cells (8×106) were cultured in DMEM supplemented with 10% FBS in a 175-cm2 flask (Corning, CA, USA) for 24 h under standard culture conditions. Next, the cells were co-transfected with EGFRvIII-specific CAR carrying SIN lentiviral vector together with the packaging vectors pMDLg/pRRE, pRSV-Rev and pMD2.G, kindly provided by Atsushi Natsume of Nagoya University School of Medicine, using calcium phosphate transfection reagent Fugene 6 (Promega Co., Madison, WI, USA), then incubated for 48 h. The lentivirus-containing culture supernatant was harvested and filtered using a 0.45-μm filter unit (Millipore, Billerica, MA, USA) to remove cellular debris. Filtrated supernatant was mixed with PEG-it Virus Precipitation Solution (5×) (System Biosciences, Palo Alto, CA, USA) and incubated for 24 h at 4°C. That mixture was then centrifuged at 1500 × g for 30 mi at 4°C, and the pellet was re-suspended in a volume 1/10 of the original using cold sterile medium at 4°C and stored at −80°C.
Lentivirus infection was performed as previously described (34). Briefly, 2×106 KHYG1 cells were suspended in 2 ml of lentiviral supernatant (MOI: 8) containing 8 μg/ml Polybrene (Sigma-Aldrich, St. Louis, MO, USA), 20% FBS, and 450 IU/ml rhIL-2. Suspended cells were seeded into 6-well flat-bottomed culture plates (Corning). Cells were centrifuged at 2600 rpm at room temperature for 45 min, then the plates were placed in an incubator at 37°C for 2 h. Infection was repeated on the same day and again the following day. Following the third transduction, cells were maintained in RPMI 1640 medium supplemented with 20% FBS and 150 IU/ml rhIL-2 at 37°C.
Detection and purification of CAR gene transduced-NK cells. Cells were stained with Biotin-SP-conjugated AffiniPure Goat Anti-Mouse IgG (Jackson Immuno Research Laboratories, West Grove, PA, USA) and Phycoerythrin (PE)-conjugated-Streptavidin (BD Biosciences, San Jose, CA, USA), then fixed with 1% paraformaldehyde/PBS at 4°C for more than 1 h. Stained cells were then acquired using a BD FACSCalibur flow cytometer and analyzed with CellQuest software (BD Biosciences). The ratio of fluorescence intensity (RFI) was calculated as the mean fluorescent intensity obtained with the sample divided by the mean fluorescent intensity of the control.
CAR NK were enriched using Biotin-SP-conjugated AffiniPure Goat Anti-Mouse IgG and Dynabeads® Biotin Binder beads (Thermo Fisher Scientific Inc.), which are streptavidin conjugated-magnetic beads, according to the manufacturer's instructions.
Growth inhibition assays. GBM cell lines were seeded into 24-well flat-bottomed culture plates (Corning) at 12×104 cells per well in 0.3 ml of DMEM supplemented with 10% FBS. Following attachment of GBM cells to the plates, 0.1 ml of NK cells was suspended in 10% FBS containing DMEM and added at NK cell-to-GBM cell (effector-to-target) ratios of 0:1, 1:1 and 5:1. The plates were then incubated for 48 h at 37°C in a humidified atmosphere containing 5% CO2. Following incubation, non-adherent cells were discarded, and adherent cells were trypsinized and stained with trypan blue dye. The number of viable non-stained cells was determined by manual counting.

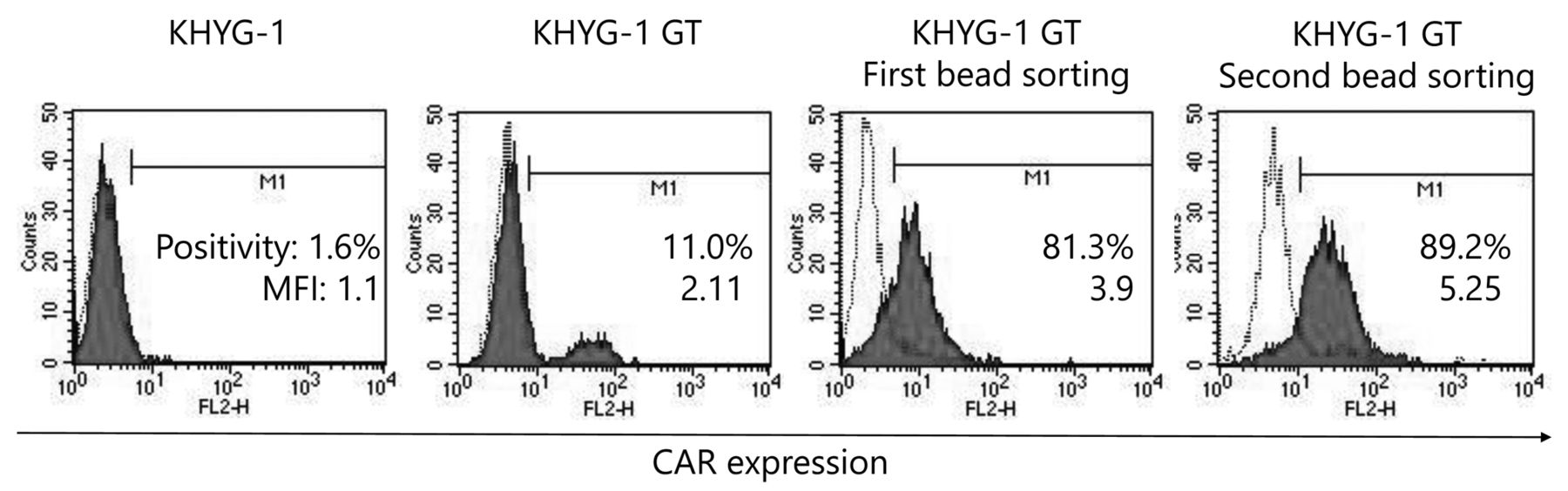
Establishment of EGFRvIII-specific CAR-expressing KHYG-1 cells. Initially, 11% of transduced CAR-KHYG-1 cells were positive for CAR expression on the cell surface (MFI: 2.11). Following a first round of magnetic bead sorting, EvCAR-expressing KHYG-1 cells were increased up to 81.3% (MFI: 3.90), then up to 89.2% after the second round of sorting (MFI: 5.25). Open histograms indicate the isotype control and filled red histograms chimeric scFv (EGFRvIII) expression on NK-KHYG-1 cells.
Apoptosis detection. Apoptosis detection assays were performed using a MEBCYTO Apoptosis Kit (Medical and Biological Laboratories, Ltd., Nagoya, Aichi, Japan), according to the manufacturer's instructions. Briefly, GBM cells were exposed at effector-to-target ratios of 0:1, 1:1, and 5:1 for 24 h at 37°C in a humidified atmosphere containing 5% CO2. Non-adherent and trypsinized-adherent cells were washed with PBS and stained with annexin V-FITC at room temperature for 15 min in the dark. Stained cells were then analyzed using a FACSCalibur and CellQuest software. NK cells were excluded by electronic gating based on forward-scatter and side-scatter characteristics. The frequency of the annexin V-positive apoptotic cells is expressed as percentage of apoptotic cells among the total number of counted cells.
Statistical methods. Values are shown as the mean±standard error (SE). Statistical significance of differences was determined using a t-test or one-way analysis of variance (ANOVA), followed by Scheffe's test. Double asterisks indicate p-values <0.01.
Results
Establishment of human NK cell line KHYG-1 transduced with EGFRvIII-specific CAR. A lentiviral vector for CAR was generated that recognizes EGFRvIII through a single-chain variable fragment (scFv) derived from the human EGFRvIII-specific monoclonal antibody (mAb) 3C10 (pELNS-3C10-CAR). In this construct, the EF1α promoter drives the CAR fusion protein integrating the 3C10-derived scFv, CD28 transmembrane (TM), and intracellular domains (ICDs), as well as the 4-1BB ICD and CD3ζ domains.
EGFRvIII-specific CAR (EvCAR) were transduced into the NK cell line KHYG-1 and expression levels of the transgene were evaluated (Figure 1). Transduced-KHYG-1 showed 11% positivity for CAR expression on the cell surface (MFI: 2.11), indicating a low level of transduction efficacy (Figure 1, second panel from left). Since the proportion of EvCAR-positive cells was low, EvCAR-KHYG-1 cells were enriched using magnetic beads. Following the first round of magnetic bead sorting, EvCAR-expressing KHYG-1 cells were increased by up to 81.3% (MFI: 3.90) (Figure 1, third panel from left), while that increase was up to 89.2% (MFI: 5.25) after the second round of magnetic sorting (Figure 1, first panel from right). The NK cells established in this manner were used in the following experiments.
EvCAR-KHYG-1 inhibits growth of EGFRvIII-expressing U87MG. To explore the growth inhibition effects of EvCAR-KHYG-1 on GBM in vitro, we performed co-cultures with the GBM cell lines U87MG and U87MG constitutively expressing EGFRvIII (U87-EGFRvIII) at a defined NK:GBM cell ratio for 72 h. In co-cultures with EvCAR-KHYG-1, the ratios of NK to GBM cells were 0:1, 1:1, and 5:1. The viable cell numbers of U87MG cells were 47.1±3.1×104 (mean±SE), 48.6±1.9×104, and 56.4±2.8×104, respectively, and of U87MG-EGFRvIII cells were 37.4±2.3×104, 36.2±2.1×104, and 13.8±2.6×104, respectively (Figure 2). On the other hand, in co-cultures with parental KHYG-1 and NK:GBM cell ratios of 0:1, 1:1, and 5:1, the viable cell numbers of U87MG cells were 41.7±6.9×104, 50.7±3.1×104, and 50.4±4.2×104, respectively, and of U87MG-EGFRvIII cells were 41.1±4.4×104, 47.0±3.7×104, and 43.8±3.4×104, respectively (Figure 2). These findings revealed that EvCAR-KHYG-1 significantly inhibited U87MG-EGFRvIII growth at an NK:GBM cell ratio of 5:1 as compared to parental U87MG. Parental KHYG-1 showed no inhibition of the growth of U87MG or U87-EGFRvIII cells.
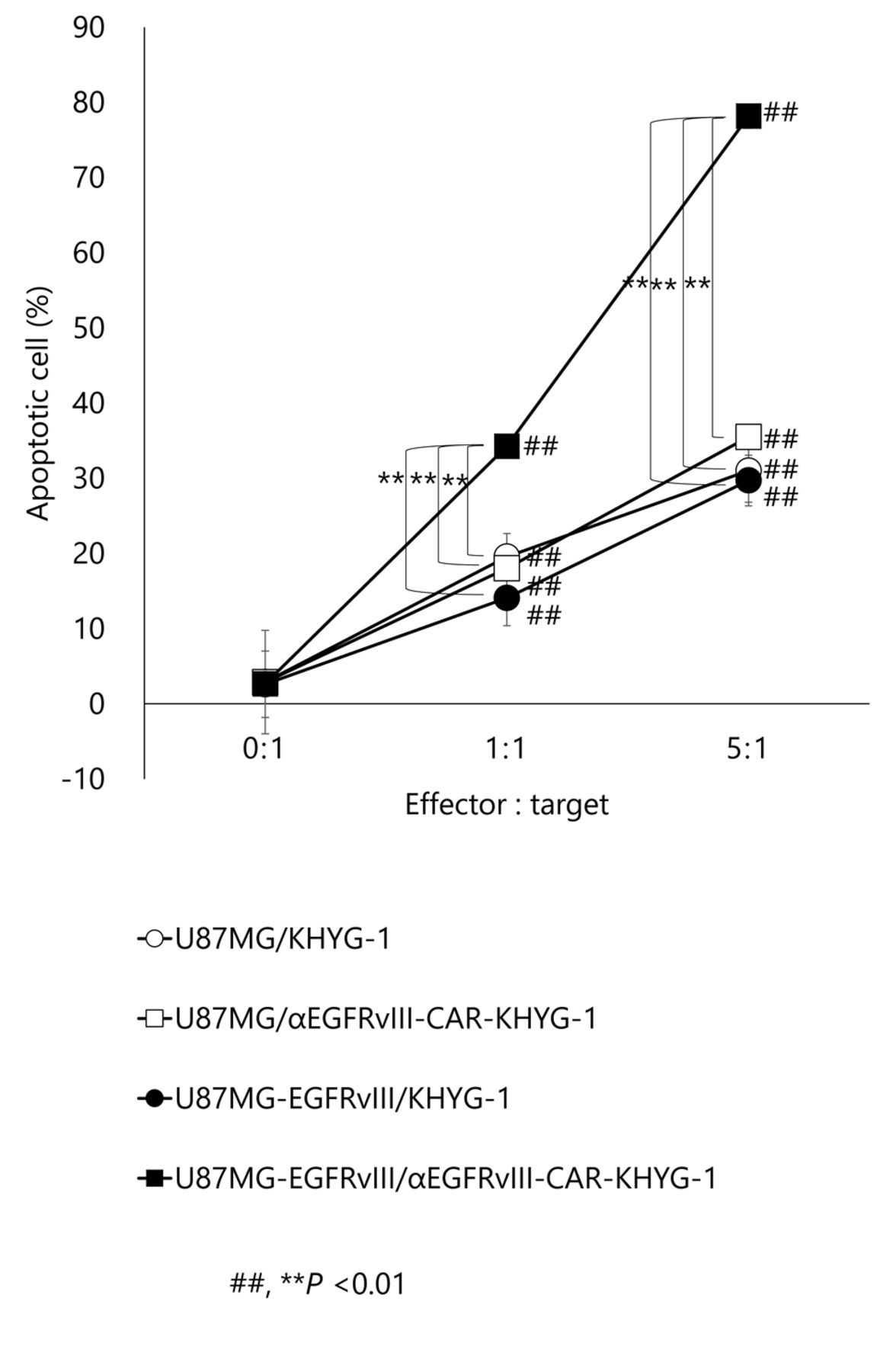
Constitutive EGFRvIII expression in U87MG cells enhances induction of apoptosis by EvCAR-KHYG-1. To investigate whether EvCAR-KHYG-1 induces tumor antigen-specific apoptosis, apoptosis detection assays with flow cytometry were used. At NK:GBM cell ratios of 0:1, 1:1, and 5:1, parental KHYG-1 induced apoptosis of U87MG cells by 2.9±0.6%, 19.5±1.1%, and 31.0±0.8%, respectively, while KHYG-1 induced U87MG-EGFRvIII cell apoptosis by 2.6±0.1%, 14.1±0.5%, and 29.7±1.3, respectively. Furthermore, EvCAR-KHYG-1 induced apoptosis of U87MG cells by 2.9±0.5%, 18.0±0.9%, 35.5±0.9%, respectively, and EvCAR-KHYG-1 strongly induced U87MG-EGFRvIII cell apoptosis by 2.6±0.1%, 34.2±1.4%, and 78.2±1.6%, respectively, at those NK:GBM cell ratios (Figures 3 and 4). Compared to parental KHYG-1, there was a significant increase in apoptosis of U87MG-EGFRvIII induced by EvCAR-KHYG-1.
Discussion
NK-92, a continuously growing human NK cell line, has been shown to be broadly cytotoxic against a spectrum of malignant cells (35-38), while other reports have noted that NK-92 cells prolonged survival of immunocompromised mouse models that underwent xenotransplantation with various types of human cancer (37, 38). On the other hand, there are few studies available regarding the human NK cell line NK-KHYG-1. This is the first report of CAR-KHYG-1 cells showing tumor-antigen specific cytotoxic effects towards EGFRvIII expressing GBM cell lines. CAR-KHYG-1 cells were found to exhibit potent antigen-specific and cytotoxic activities towards U87 GBM cells stably expressing EGFRvIII.
For genetic engineering of primary CAR NK cells, viral vectors, such as retrovirus- and lentivirus-based vectors, have typically been applied, due to their stable integration into the genome. Lentiviral vectors have a moderate number of insertion mutations, though their lentiviral transduction efficiency is quite low (8-16%) with PB-derived NK cells (28). The present findings also showed a low transduction rate (11%), however following two rounds of enrichment of EvCAR-KHYG-1 cells with magnetic beads increased their presence to 89.2%, indicating a good level of purification. CAR-engineered T cells targeting the B-cell differentiation antigen CD19 have demonstrated remarkable clinical efficacy in patients with lymphomas and leukemias of B-cell origin (19, 20). NK cells represent another important cell type for such adoptive immunotherapy, though CAR-mediated retargeting of NK cells has not been frequently attempted. Currently, only a few clinical trials using CAR-transduced NK cells, including CAR-modified NK-92 and primary NK cells, are in progress (28, 39).

Growth inhibition assay findings. An E:T ratio of 5:1 resulted in significant suppression of the increase in the number of viable cell targets, U87MG or U87MG-EGFRvIII. (##, **p<0.01; n=4).
Despite the early success of CAR T therapy, particularly as treatment for hematological malignancies, large-scale clinical application of CAR T therapy may be limited by individualized preparation methods and various potential side effects (28, 40). Furthermore, CAR T therapy can result in cytokine storms (30) and may induce graft-versus-host disease (GVHD) in an allogeneic transplantation setting (29).
NK cells have been proposed to be superior as CAR drivers for T cells (27) and shown to have several advantages when used in CAR-based immunotherapy as compared to T cells (28, 41, 42). EvCAR-KHYG-1 has the ability to suppress an increasing number of target cells in a tumor antigen specific manner, as indicated by the present tumor growth inhibition assay findings (Figure 3). Moreover, apoptosis detection assay results showed that EvCAR-KHYG-1 significantly increased EFGRvIII tumor antigen-specific apoptosis.
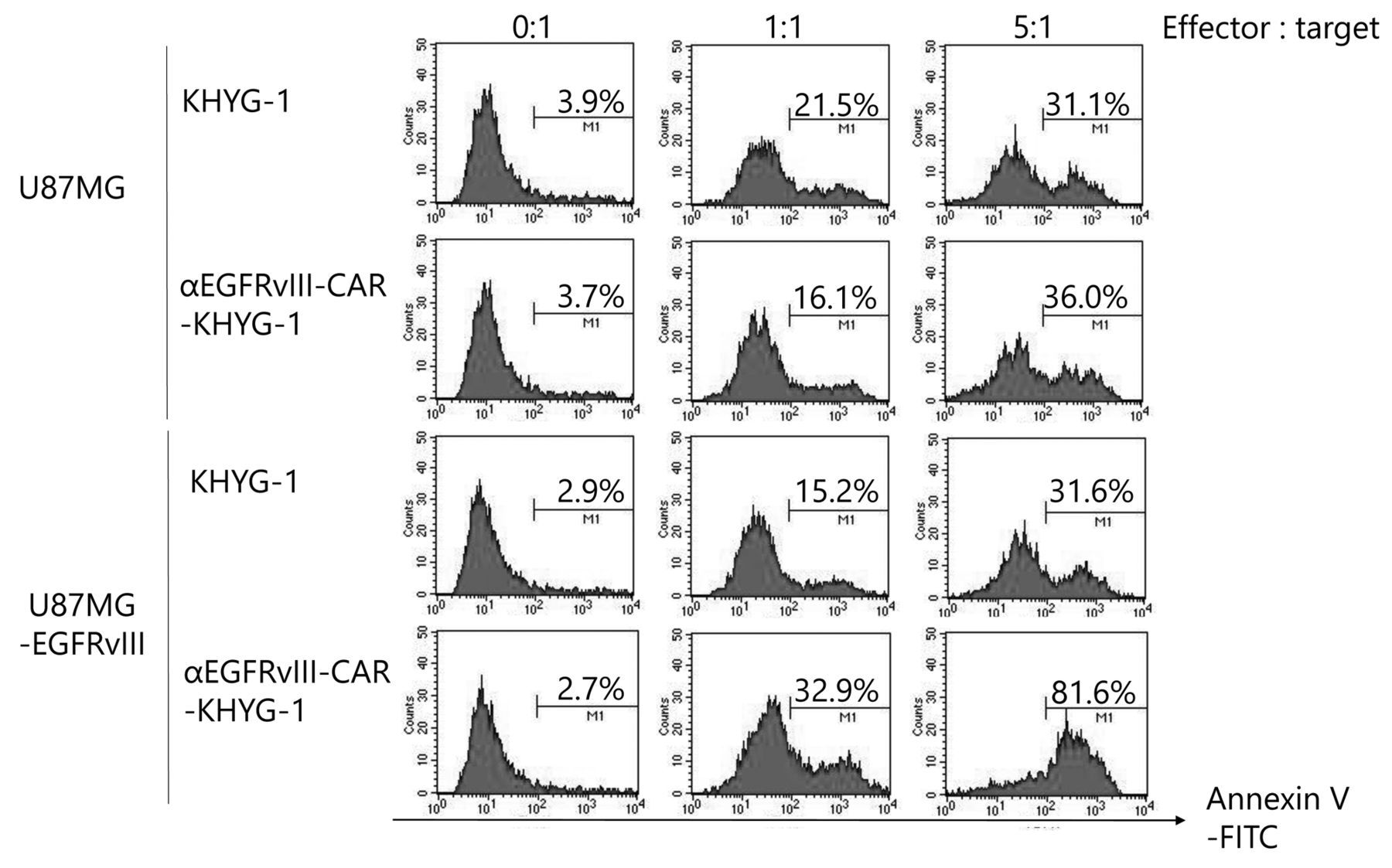
Representative findings of apoptosis detection assays. EvCAR-KHYG-1 at NK:GBM cell ratios of 0:1, 1:1, and 5:1 strongly induced U87MG-EGFRvIII apoptosis by 2.6±0.1%, 34.2±1.4%, and 78.2±1.6%, respectively.
CARs offer several advantages as compared with traditional T-cell receptor (TCR)-mediated targeting of tumor antigens. Unlike TCRs, CARs do not require antigen presentation by the major histocompatibility complex (MHC), which is often down-regulated in gliomas (43, 44). Use of CARs has evolved over the recent decade, with progressively increasing co-stimulatory activity gained (16, 45). In addition to a single signaling unit derived from the CD3ζ chain, the second generation of CARs incorporates the intracellular domain of CD28, a co-stimulatory molecule, or the tumor necrosis factor (TNF) receptor family member CD137 (4-1BB). Incorporation of both CD28 and CD137 has been shown to enhance the ability of these receptors to stimulate cytokine secretion and, hence, the antitumor efficacy of third generation CARs (4, 31). In general, second generation CARs in NK cells are more active than the first generation (28). The important co-stimulatory molecule 4-1-BB is combined with CD3ζ as an intracellular domain, which significantly enhances the targeted lysis of CAR-NK cells (28, 46-48).
EGFRvIII expression has been reported in a variety of tumor types, including GBM, while it is rarely observed in normal tissues (11, 12, 49-55). Another report noted that it was expressed in 24% to 67% of GBM cases and that expression was found to be an independent negative prognostic indicator in patients surviving ≥1 year (56). As an off-the-shelf therapeutic modality, continuously expanding CAR-KHYG-1 cells may show clinical usefulness for treatment of cancer patients, particularly in cases where the infrastructure or suitable donors for a CAR T approach are not readily available. Even at low E/T ratios, EvCAR-KHYG-1 cells displayed more pronounced cell killing than KHYG-1 cells. That may contribute to both direct and indirect antitumor effects, and should be considered as part of further development of this approach for clinical applications.
In summary, the EvCAR-KHYG-1 cell line was established by self-inactivated lentivirus-based transduction of the EvCAR gene and magnetic bead-based purification of EvCAR-expressing NK cells. The EvCAR-KHYG-1 cell line inhibited GBM cell growth via apoptosis in an EGFRvIII-expressing specific manner. Thus, EvCAR-KHYG-1 may become an effective treatment option for patients with GBM.

Apoptosis of targets induced by parental KHYG-1 cells was generally <13% at an E:T ratio of 1:1 and <30% at a 5:1 ratio. In contrast, apoptosis induced by EvCAR-KHYG was significantly greater, up to 31-37% at an E:T ratio of 1:1 and 75-81% at a ratio of 5:1. (##, **p<0.01; n=3).
Acknowledgements
This study was supported by a Grant-in-aid for Scientific Research (No. 16K10767) from the Japan Society for the Promotion of Science.
Footnotes
↵* These Authors contributed equally to this study.
Conflicts of Interest
None of the Authors have conflicts of interest to declare in regard to this study.
- Received July 21, 2018.
- Revision received August 2, 2018.
- Accepted August 6, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}