


Abstract
In tumor immunity, invariant natural killer T (iNKT) cells play a pivotal role as a link between the innate and adaptive immune systems. With a precisely regulated activation mechanism, iNKT cells have the ability to respond quickly to antigenic stimulation and rapidly produce cytokines and chemokines, and subsequently an effective antitumor immune response. The development of iNKT cell-targeted active immunotherapy enables, not only an antitumor immune response through innate and acquired immunity, but also the conversion of an immunosuppressive into an immunogenic microenvironment. This review is focused on the activation mechanism and the role of iNKT cells after therapeutic active immunization. The therapeutic strategy targeting iNKT cells is expected to be applied to clinical practice in combination with surgery and chemotherapy.
The clinical development of checkpoint inhibitor-based immunotherapy has introduced a new era in cancer treatment (1, 2). Durable responses, characteristic of acquired immunity, can be seen clinically in patients with several types of cancers, including melanoma (3, 4). The emergence of strong, evidence-based therapy has enabled immunological therapeutic interventions in a clinical setting, which then has highlighted a need to demonstrate the effects of combinations with other types of therapy, including conventional and immunotherapy, as well as treatment that is still being developed.
Natural killer T (NKT) cell-based immunotherapy is one of the most promising types of immunotherapy currently in development. In tumor immunity, the immune systems participate in immune surveillance against tumor development and respond to the foreignness of tumor cells. The innate immune cell population recognizes tumor-associated antigens and danger signals from tumor cells and responds quickly to them. Effector cells typified by natural killer (NK) cells start to eliminate tumor cells directly. In addition, antigen-presenting cells such as dendritic cells (DCs) and macrophages take up the antigens, and are responsible for induction and enhancement of the acquired immunity, followed by the subsequent formation of antigen-specific B and T cells (5). One of the key players that link the innate and the adaptive immune systems is the NKT cells. They have the ability to respond quickly to antigenic stimulation and rapidly produce cytokines (6). Mediated by these cytokines, crosstalk between applicable immune cells activates the whole immune system and enables an effective antitumor immune response (7, 8). Invariant natural killer T (iNKT) cells comprise an innate-like subset of T lymphocytes that carry a distinctive invariant T cell receptor (TCR) α chain paired with a limited repertoire of TCR β-chains; these cells recognize a variety of lipid antigens when bound to a major histocompatibility (MHC) class I-like molecule, CD1d (9). Many researchers have explored the possibility of cancer immunotherapy using iNKT cells, that is an attractive tool to activate both innate and adaptive immunity. Although novel treatments including induced pluripotent stem (iPS) cells, chimeric antigen receptors, biphasic antibodies, and nanotechnology are being developed one after another (10, 11), this review focuses on iNKT cell-targeted active immunotherapy.
Activation of iNKT Cells
After activation, NKT cells can secrete large amounts of various types of cytokines and exhibit both proinflammatory and immunoregulatory characteristics. Depending on their T cell receptor (TCR), CD1d-restricted NKT cells are subdivided into type I or invariant NKT (iNKT) cells, and type II NKT cells. Type I NKT cells have an invariant TCR α chain, whereas type II cells have more diverse alpha-beta TCRs. The TCR of Type I NKT cells is composed of a single α chain (Vα14Jα18 in mice and Vα24Jα18 in humans) paired with limited diverse β chains (Vβ8.2, 7 or 2 in mice and Vβ11 in humans) (9). Type II NKT cell is a subset involved in immunosuppression in tumor-bearing hosts or in autoimmune disease (12). This report discusses iNKT cells.
iNKT cells receive signals through various types of receptors. The TCRs of iNKT cells recognize lipid antigens that comprise endogenous self-lipids and exogenous non-peptide lipids, mainly microbe-derived presented through molecular interaction with the CD1d molecule (13). CD1d is a non-polymorphic MHC class I-like molecule constitutively expressed on antigen-presenting cells such as DCs, B cells, and macrophages. The well-studied agonist for iNKT cells is α-galactosylceramide (α-GalCer or KRN 7000), which is isolated from a marine sponge (14).
Upon antigenic stimulation, iNKT cells respond within a few hours by producing large amounts of Th1, Th2, and Th17 cytokines (15). This rapid response can be attributed to their storage of cytokine mRNA in the cytoplasm before activation (16). Interestingly, iNKT cells have phenotypically activated and memory-like properties (17). In addition, the multifunctionality of cytokine production is due to the capacity of iNKT cells to differentiate into mature effector subsets that produce IFN-γ, IL-4, or IL-17 in the thymus. These subsets are designated as NKT1, NKT2, and NKT17 cells, respectively, and their lineage properties are determined by key transcriptional factors including promyelocytic leukemia zinc finger (PLZF), T-cell-specific T-box transcription factor (T-bet) in Th1 lineage, GATA binding protein 3 to DNA sequence (GATA-3) in Th2 lineage, and retinoic acid-related orphan receptor γt (ROR γt) in Th17 lineage, similar to the differentiation of conventional T cells (18, 19).
An important point in iNKT cell research is the method of administration of foreign antigens (Figure 1), which is termed active immunotherapy. Active immunotherapy is a therapeutic intervention that causes activation of the immune system in vivo by exogenous stimulation. After α-GalCer was identified as a specific ligand for iNKT cells, the effects of administration of free α-GalCer have been studied enthusiastically. A drawback of administering free α-GalCer is that it causes iNKT cells to adopt an anergic state, causing unresponsiveness to sequential stimulation with α-GalCer (13). Fujii and colleagues demonstrated prolonged IFN-γ production and antitumor effect after intravenous administration of bone marrow-derived DCs loaded with α-GalCer (DCs/Gal), that express a high level of CD1d (20).
In addition, from a clinical point of view, it should be noted that administration of α-GalCer may cause liver injury that often results in lethality (21). We showed that this adverse effect is due to antigen presentation by intrahepatic vascular endothelial cells and that it is rarely lethal in DCs/Gal administration (22).
The Role of iNKT Cells in Targeted Active Immunotherapy
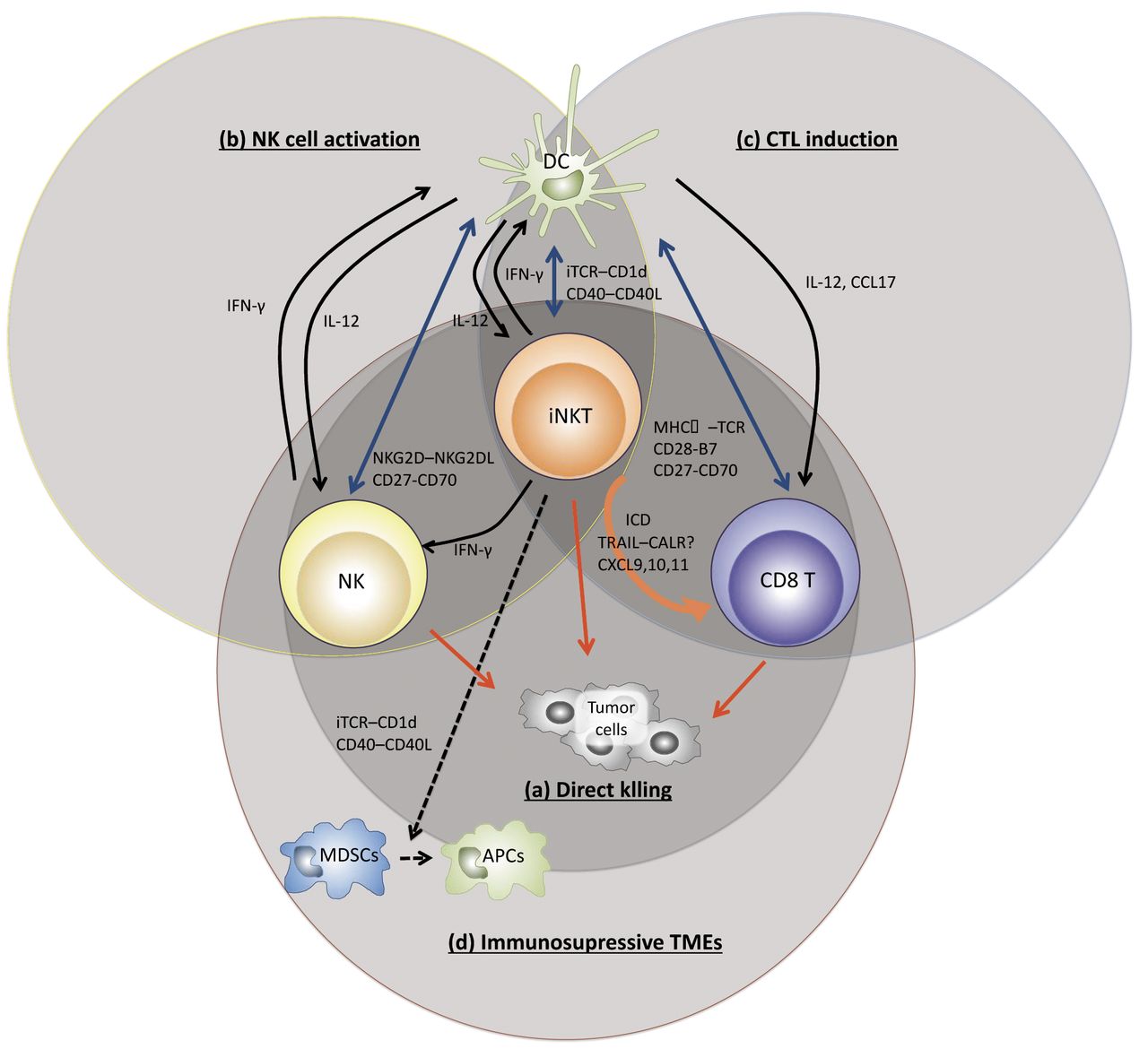
In the antitumor immune response following active immunotherapy, the mechanism of action of iNKT cells are as follows: (a) direct tumor killing, (b) induction and activation of NK cells, (c) induction of cytotoxic T lymphocytes (CTLs), (d) modulation of immunosuppressive tumor microenvironment (Figure 2). This type of immunotherapy activates iNKT cells and enables NK cells to eliminate MHC-negative tumors, and antigen-specific CD8 cytotoxic T cells to kill MHC-positive tumors.
Direct tumor killing activity of iNKT cells. The direct killing ability of iNKT cells has been analyzed against tumors expressing CD1d in vitro. The function is exerted through the perforin/granzyme B (23), Fas - Fas ligand system (24), and tumor necrosis factor-α-related apoptosis-inducing ligand (TRAIL) (25). These studies revealed that tumors expressing CD1d may be the targets of iNKT cells. But declining expression of CD1d on tumor cells does not necessarily reflect disease progression (26).
Induction of NK cells by iNKT cell activation. With regard to the response of NK cells, the activated iNKT cells secrete IL-2 and IFN-γ, and directly activate NK cells (27). In addition, after activation, iNKT cells express CD40L and promote maturation of DCs through a CD40-CD40L signal. DCs produce IL-12 and expresses NKG 2 D ligands and CD 70 (CD 27 ligand) (28, 29). Each of these signals can further enhance the response of NK cells and are essential for NK cells to exhibit sufficient antitumor effect (30); we clarified that this is important in a mouse B16 melanoma lung metastasis model (31).
Clinically, in cancer patients, it is difficult to secure adequate amounts of DCs. To solve this problem, we showed that allogenic bone-marrow-derived DCs /Gal that meet the above conditions, produce a guaranteed therapeutic effect (32).
Efficient induction of CTLs against tumor. The induction of CTLs is accompanied by maturation of DCs. In the tumor tissue, most of DCs are immature. But maturation of DCs is essential to induce a sufficient CTL-mediated reaction. DCs and iNKT cells interact with CD1d/antigen complexes. Ligation of APC-expressed CD40 with up-regulated CD40L on iNKT cells induces maturation of DCs that express co-stimulation molecules on the cell surface and gain the capacity of IL-12 production. IL-12 secreted by mature DCs induces IFN-γ production by NKT cells (33). IFN-γ secreted by iNKT cells and NK cells promotes maturation of DCs, but expression of co-stimulatory molecules, such as CD40, CD80, CD86, on DCs are important for the CTL priming (34, 35). Moreover, iNKT cells and fully activated DCs produce CCL17, which attracts CCR4-positive CD8 T cells, for CTL priming (36).

Invariant natural killer T (iNKT) cell activation by active immunization. A: Administration of free α-galactosylceramide (α-GalCer). The problem of administration of free α-GalCer is to bring about an anergic state and lethality. B: Administration of dendritic cells (DCs) loaded with α-GalCer (DCs/Gal) with or without tumor antigen. Administration of DCs/Gal has less capacity to induce antigen-specific CD8 T cells.C: Administration of adjuvant vector cells, including tumor cells loaded with α-GalCer or tumor antigen mRNA-transfected, allogeneic CD1d-positive cells loaded with α-GalCer. TCR: T cell receptor; MHC I: major histocompatibility I; iTCR: invariant TCR; mRNA: messenger ribonucleic acid.
Utilization of iNKT cells in cancer vaccine treatment. Co-administration intravenously of tumor antigen with α-GalCer brings about rapid DC maturation and induction of an effective adaptive immune response to tumors (34, 35). Several optimal conditions are necessary for successful treatment. The context is that immature DCs phagocytose tumor antigens and, after adequate maturation by iNKT cells, they present tumor antigens to antigen-specific CTLs. Therefore, it is a necessary condition that activated iNKT cells are presented with tumor antigen and endogenous immature DCs simultaneously. The administration of cells expressed high level of CD1d such as B cells or myeloid-derived suppressor cells (MDSCs) and, loaded with α-GalCer and peptide of tumor antigen, can induce CTL responses (37, 38), but cannot generate memory formation easily. Fujii and colleagues indicated the essential conditions to generate a memory response as follows: (1) the numbers and functions of iNKT cells and APCs, (2) the ligands, (3) the properties of host APCs, and (4) the level of CD1d expression (39). Under these conditions, the magnitude of the innate immune response can shape the subsequent immune response with adequate antitumor effect. At present, the most effective cell option for adjuvant vectors is antigen mRNA-transfected fibroblasts, called artificial adjuvant vector cells (aAVC) (40). More recently, a new NKT cell glycolipid ligand with strong adjuvant effect called RK have been identified. RK-loaded DCs induced strong antitumor effect and long-lasting memory reaction (41). The development of these therapies, not only dramatically pushed research to preclinical models, but also strongly indicated that this approach may eventually provide a clinically promising treatment.

The role of invariant natural killer T (iNKT) cells in antitumor responses in tumor microenvironments (TMEs). A: Direct tumor killing. iNKT cells exert direct cytotoxicity to tumor cells. B: Induction and activation of natural killer (NK) cells. Activated iNKT cells directly activate NK cells via interleukin (IL)-2, interferon (IFN)-γ, and indirectly via IL-12 secreted by dendritic cells (DCs) and natural killer group 2 member D (NKG 2 D) ligands and CD 70 (CD 27 ligand) expressed on dendritic cells. C: Induction of CTL priming and generation of memory response. The vector cells are killed by iNKT cells and NK cells, and then tumor antigen can be captured by endogenous DCs. The DCs promote iNKT cell-induced maturation. The activated DCs can then induce an antigen-specific T cell response. D: Modulation of immunosuppressive TMEs. iNKT cells covert immunosuppressive to immunogenic microenvironments by Th1 cytokines production, maturation of myeloid-derived suppressor cells (MDSCs) to antigen-presenting cells (APCs), and induction of immunogenic cell death (ICD) of tumor. TCR: T cell receptor; MHC I: major histocompatibility I; iTCR: invariant TCR; CXC17: chemokine ligand 17; CXCR4: chemokine receptor 4.
Modulation of immunosuppressive tumor microenvironment. MDSCs play a crucial role to establishing an immunosuppressive tumor microenvironment (42). The ability of iNKT cells to convert an immunosuppressive to an immunogenic microenvironment is based on their capacity to promote the maturation of MDSCs into mature myeloid cells, which results in resolution of MDSC-mediated immunosuppression (37, 43). Next, the ability to convert the tumor microenvironment of iNKT cell-targeted therapy in a poor immunogenic tumor will be described. Many researchers are convinced that induction of T cells is the key to increase the success of checkpoint inhibitors against tumors. The immunogenic cell death (ICD) refers to a unique tumor cell death mediated through activation of innate immune cells and facilitation of the acquired immune response of antigen-specific T cell towards dead cell-derived antigens (44). When tumor cells die by necrosis, they expose on the cell surface or secrete molecules called danger-associated molecular patterns (DAMPs). The DAMPs are molecules that are present in tumor cells and released outside the cell at the time of cell death, but they function as immunostimulatory molecules and trigger ICD to convert the tumor microenvironment from a T-cell poor to a T-cell rich. DAMPs intimately associated to NKT cells, are high mobility group box1 (HMGB1), adenosine triphosphate(ATP), and calreticulin(CALR). Presumably, HMGB1 binds to TLR4 on the surface of CD1d-positive cells, which results in activation of iNKT cells (45). However, a series of cellular vaccine therapies are considered TLR-independent (34).
ATP regulates immunomodulatory receptors on immune cells and enhances the strength of the TCR signal even with weak stimulation (46).
In addition, CALR normally resides in the lumen of the endoplasmic reticulum (ER), from where it translocates to the surface of apoptotic tumor cells on induction of ICD. The tumor cells can then be recognized and taken up by DCs. As mentioned above, iNKT cells up-regulate TRAIL that directly kills target cells. Interestingly, TRAIL signaling induces CALR on tumor cells. CALR directly interacts with TRAIL and its receptor-signaling complex, as well as with other TNF family members. While further studies are required on how the CALR-TRAIL interaction is related to the antitumor effect mediated by iNKT cells, this interaction is expected to be the key to elucidate the mechanism of iNKT cell-mediated immunotherapy in various cancers (47). iNKT cell-targeted active immunotherapies cause ICD. In fact, aAVC against melanoma increased the concentrations of CXCL9, 10, and 11 in the tumor sites and in DCs, and eventually induced antigen-specific T cells into the tumor (48). Especially, CXCL10 is one of DAMPs components, that stimulate and recruit activated T cells (49). Furthermore, CD70, 4-1-BBL, and IL-15Rα on DCs are essential for generating memory in antigen-specific CTLs.
Combination therapy with surgery or chemotherapy. In many solid cancer types, surgery is the best-choice treatment for localized disease with the objective of achieving complete resection of the tumors. However, once a postoperative recurrence occurs, it is directly related to death. In the relapse model after resection of the primary tumor in a triple-negative breast tumor, treatment with α-GalCer-loaded DCs limited tumor metastasis, prolonged survival, and provided curative outcomes (50). We elucidated that the frequency of MDSCs of the peripheral blood can be a recurrence marker in a B16 melanoma model (51). The aim of this treatment is to regulate the number of MDSCs and show its effect by inducing antigen-specific reactions. In a similar model, the effectiveness of combination of α-GalCer-loaded DCs with gemcitabine or cyclophosphamide, that is a chemotherapy that causes ICD, has also been proposed (52).
Conclusion
Currently, based on preclinical models, iNKT cell-based immunotherapies are expected to increase the rate of complete response after resection of tumor or to maintain a state of long-term stable disease. In order to realize this therapeutic success, the generation of effective CTL memory cells is necessary by inducing ICD and establishing an effective innate immune response. For this purpose, further analysis of iNKT cell-targeted immunotherapy should focus on development of effective specific ligands and elucidation of their detailed interaction with tumor antigens.
Acknowledgements
This work was supported by a Grant-in-Aid from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (KY: No. 16K10541).
Footnotes
This article is freely accessible online.
Conflicts of Interest
The Authors have no conflict of interest in regard to this study.
- Received April 11, 2018.
- Revision received May 16, 2018.
- Accepted May 17, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}