


Abstract
Background/Aim: Natural products extracted from plants can be potent for developing pharmaceutical products. Hispidin, a polyphenolic compound mainly derived from the medicinal mushroom Phellinus linteus, has been shown to have a therapeutic potential against cancer cells. Pancreatic cancer is one of the most aggressive solid malignancies with high resistance to existing drugs. Cancer stem cells (CSCs) are responsible for chemoresistance. The present study aimed to evaluate the anticancer effects of hispidin on pancreatic CSCs. Materials and Methods: The cytotoxic effects of hispidin on BxPC-3 and AsPC-1 pancreatic cancer cells and BxPC-3 CD44+ CSCs and the synergistic effects of gemcitabine and hispidin on CSCs were evaluated by a series of in vitro experiments including the 3-(4,5-dimethythiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), fluorescence-activated cell sorting, colony forming, Transwell assay, immunocytochemistry, sphere-forming, and western blot assays. Results: Hispidin exerted antitumor effects against both BxPC-3 pancreatic cancer cells and CSCs. Furthermore, it was found that hispidin sensitized pancreatic CSCs to gemcitabine and promoted the therapeutic efficacy of gemcitabine. Conclusion: Hispidin might be a novel chemosensitizer for gemcitabine and a potential synergistic agent for increasing the therapeutic index of gemcitabine as a treatment for pancreatic cancer.
Pancreatic cancer is one of the deadliest malignancies; it is currently the fourth leading cause of cancer-related deaths, with a median survival period of 5-8 months. It is assumed that pancreatic cancer will be the second leading cause of cancer-related deaths by 2030 (1). Only around 5% of patients with pancreatic cancer survive for more than 5 years after initial diagnosis (2). The mortality rate associated with pancreatic cancer is almost equal to its incidence rate because of late diagnosis and poor prognosis. The poor prognosis is attributable to early metastasis in pancreatic cancer development, a strong migratory nature, late presentation due to the difficulties associated with the initial diagnosis, and unresponsiveness to most conventional cancer therapies (3, 4). Pancreatic cancer stem cells (CSCs) sustain tumor formation, metastasis, and chemoresistance of pancreatic cancer. Conventional chemotherapy and radiotherapy only affect rapidly growing cancer cells and fail to eliminate CSCs (5). Recently, CD44, an adhesion molecule expressed on the surface of CSCs, was found to play a role in determining the characteristics of pancreatic CSCs and act as a master regulator of CSCs (6, 7). Therefore, in this study, we focused on CD44+ pancreatic cancer cells because improved new strategies are needed to target CSCs.
Surgery is not beneficial for the majority of patients with pancreatic cancer because only 15-20% of pancreatic tumors are local and therefore anatomically resectable upon diagnosis. Furthermore, even after surgical resection, the resulting prognosis is very poor due to the high rate of metastasis. Therefore, chemotherapeutic drugs are frequently needed, although the response rates to conventional drugs have been reported to be lower than 25% (3, 8). Gemcitabine plays a major role in chemotherapy for pancreatic cancer as the first-line chemotherapeutic drug, by regulating stemness and chemoresistance via phosphorylation of multiple proteins and signaling pathways such as the nicotinamide adenine dinucleotide phosphate-oxidase (NOX)/reactive oxygen species (ROS)/nuclear factor-kappa B (NF-κB)/signal transducer and activator of transcription 3 (STAT3) pathway (9). However, the reduced therapeutic efficacy of certain doses of gemcitabine because of intrinsic and acquired resistance of pancreatic cancer cells remains a major challenge, and there is an urgent need for diagnostic and treatment strategies to increase tumor sensitivity to gemcitabine (10). Recent studies have suggested that combination therapy with a targeted medicine that inhibits NF-κB activity increases the sensitivity of pancreatic cancer cells to gemcitabine, as the activation of NF-κB is associated with pancreatic cancer progression via up-regulation of cell proliferation, chemoresistance, invasion, angiogenesis, metastasis, and suppression of apoptosis (11, 12). However, the results from studies thus far have been inadequate and new approaches are needed to maximize the effectiveness of gemcitabine.
Hispidin is a polyphenolic compound, mainly derived from a valuable medicinal mushroom, Phellinus linteus (Figure 1A); it is well known for its strong anticancer, antioxidant, and DNA-damage protection activities. Studies have already reported the ability of hispidin to inhibit tumor growth and metastasis (13). Hispidin has also been reported to inhibit the transcriptional activity of NF-κB (14, 15). Furthermore, according to some recent studies, hispidin is cytotoxic against human lung, squamous, liver, and pancreatic cancers (16). Moreover, previous studies have revealed that hispidin inhibits cell viability and mediates apoptotic signals in CSCs by generating reactive oxygen species (17).
In the present study, we explored the unique anticancer activity of hispidin and its activity as a chemosensitizer of gemcitabine in malignant pancreatic cancer. Even though previous studies have shown the anticancer activity of hispidin against diverse types of cancer cells, to our knowledge, this is the first study to show the cytotoxic activity of hispidin in CSCs and show the effective and promising synergistic therapeutic effect of hispidin used in combination with gemcitabine in pancreatic cancer cells.
Materials and Methods
Reagent. Hispidin [6-(3,4-dihydroxystyryl)-4-hydroxy-2-pyrone; solid, ≥98% of purity by high-performance liquid chromatography, derived from P. linteus was purchased from the National Development Institute of Korean Medicine (NIKOM) (http://www.nikom.or.kr/eng/main.jsp), Gyeongsan, Republic of Korea.
Cell culture and treatments. BxPC-3 and AsPC1 human pancreatic cancer cell lines were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). Human pancreatic cancer cell lines BxPC-3 and AsPC1 were maintained in Dulbecco's modified Eagle's medium (DMEM) (Invitrogen, Carlsbad, CA, USA) with 10% fetal bovine serum (FBS; Welgene, Gyeongsan, Gyeongsangbuk-do, South Korea) and 1% penicillin (100 U/ml) and streptomycin (100 mg/ml). BxPC-3 CD44+ cells (CSCs) were cultured in the above complete media supplemented with 10 ng/ml human epidermal growth factor (hEGF) (Sigma Aldrich, St. Louis, MO, USA) and 20 ng/ml basic fibroblast growth factor (bFGF).
Transwell migration and Matrigel invasion. In vitro migration and invasion assays were performed using Transwell 24-well chambers with 8.0-μM pore polycarbonate membranes (Merck Millipore, Darmstadt, Germany) without (migration) or with (invasion) Matrigel. Briefly, 200 μl of BxPC-3 CD44+ cell suspensions treated with or without 150 μM of hispidin in 0.5% FBS-containing medium were added separately to the upper chambers (1×105 cells/chamber). The bottom chamber was filled with 800 μI of the medium supplemented with 20% FBS as a chemoattractant. Cells were then incubated for 24 h at 37°C and 5% CO2. Cells which passed through the coated membrane to the lower surface were then fixed with 4% paraformaldehyde and stained with 0.1% crystal violet for 1 h. Images were then captured using a microscope.
Immunocytochemistry. BxPC-3 CD44+ cells treated with or without hispidin or gemcitabine were fixed with 3.7% formaldehyde in 1× phosphate-buffered saline (PBS) for 10 min at room temperature. They were then blocked with 1× PBS containing 0.5% Triton X-100 and 1% bovine serum for 60 min at room temperature and incubated with primary antibodies to homing cell adhesion molecule (HCAM) (DF1485), NANOG (1E6C4), Sex-determining region Y)-box 2 (SOX2) (E-4), (Santa Cruz Biotechnology) for 18 h. On the following day, the cells were washed with 1× PBS with Tween 20 and incubated with secondary antibodies, goat anti-rabbit immunoglobulin (Ig)G (Alexa Fluor 488; Abcam, Cambridge, MA) and goat anti-mouse IgG-PE (Santa Cruz Biotechnology, Dallas, TX, USA) respectively for 2 h at room temperature. Nuclei were visualized by 4’,6-diamidino-2-phenylindole (DAPI) staining. Images were acquired and observed qualitatively after 20 min of DAPI staining using a microscope.
Sphere-formation assay. Pancreatic CSCs treated with 150 μM of hispidin for 48 h and non-treated CSCs (2×103) were seeded in 2 ml of cell culture medium [serum-free DMEM/F12 (Gibco, Grand Island, NY, USA)] containing B27 supplement (1:50; Invitrogen), 20 ng/ml EGF (Calbiochem), and 0.5% bovine serum albumin (Sigma Aldrich) per well in a 6-well ultra-low cluster plate (Corning Inc, Corning, NY, USA). The spheres were obtained after 7 days by using an inverted microscope. Spheres were fixed for 30 min with 30.03 g/mol formaldehyde solution. Cells were then rinsed twice with PBS and incubated in blocking solution consisting of 1× PBS containing 0.5% Triton X-100 and 1% bovine serum for 1 h. Cells were allowed to incubate overnight at 37°C with primary antibody solution consisting 0.1% Triton-X100, 10% NaNO3 and 1× PBS. Cells were rinsed twice in 1× PBS prior to incubating with secondary antibody for 2 h in the dark place at room temperature. Cells were then rinsed twice with 1× PBS and counterstained with DAPI diluted in 1× PBS for 20 min prior to visualization and image capturing using microscope.
Western blotting. Lysates from hispidin and gemcitabine-treated or untreated BxPC-3 and BxPC-3 CD44+ cells were collected in NP-40 lysis buffer (150 mM NaCl, 1% Nonidet p-40, 50 mM Tris, pH 8.0, and a protease inhibitor cocktail). Equal amounts of proteins from each cell lysate were resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis. Thereafter, the proteins were transferred to nitrocellulose membranes (BIO-RAD, Hercules, CA, USA), which were blocked with 10% skim milk and 0.1% Tween 20 in Tris-buffered saline for 1 h at room temperature. Primary antibodies [against CD44, NF-κB, p53, B-cell lymphoma 2 (BCL2), cleaved caspase-3, cleaved poly (ADP-ribose) polymerase (PARP), NANOG, and SOX2 were added overnight at 4°C, followed by incubation with a secondary antibody at room temperature for 4 h. Antibody expression levels were checked qualitatively against glyceraldehyde 3-phosphate dehydrogenase (GAPDH) which was used as the control and obtained from AbFrontier Technology (Seoul, South Korea).
Colony-formation assay. BxPC-3 CD44+ cells (1×103 cells/well) were plated in 6-well plates and maintained in an incubator at 37°C with 5% CO2 for 7 days. The cells were then washed with 1× PBS, fixed with 3.7% formaldehyde for 10 min, treated with methanol for 20 min, and stained with crystal violet for 30 min. The plates were washed three times with 1× PBS prior image capturing by using a camera.
Annexin V staining. BxPC-3 CD44+ cells treated withor without hispidin for 48 h were washed with 1× PBS, stained with annexin V binding buffer (BD Biosciences, San Diego, CA, USA) and labeled with fluorescein isothiocyanate-conjugated Annexin V according to the manufacturer's protocol. Cells were sorted on a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA, USA).
2’,7’-Dichlorodihydrofluorescein diacetate (DCF-DA) assay. The reactive oxygen species (ROS) in BxPC-3 CD44+ cells with and without 50, 100 and 150 μM of hispidin treatment were determined using DCF-DA (Invitrogen). Cells were incubated with 20 mM of DCF-DA for 15 min at 37°C and then washed with 1× PBS. DCF-DA% was analyzed on a FACSCalibur flow cytometer (BD Biosciences).
Data analysis. Statistical analyses were conducted using SPSS v.20.0.1 software (SPSS Inc., Chicago, IL, USA). The chi-square test or Fisher's exact test was used as appropriate. Differences with values of p<0.05 were considered statistically significant.
Results
Hispidin significantly down-regulates cell proliferation and stemness of BxPC-3 and AsPC1 pancreatic cancer cells. To confirm the cytotoxic effects of hispidin, we initially assessed the antiproliferative effect of hispidin on AsPC1 and BxPC-3 pancreatic cancer cells in a dose-dependent manner. As shown in Figure 1B, the MTT assay revealed that hispidin reduced the growth of the pancreatic cancer cells. As hispidin was more effective against BxPC-3 cells than AsPC1 cells, BxPC-3 cells were selected to further study the cytotoxic effects of hispidin on pancreatic cancer cells.
To determine the mechanisms underlying the cytotoxic effects of hispidin, the expression of p53, NF-κB, apoptosis-related proteins, CSC marker CD44, and stemness genes in cells treated with 50, 100 and 150 μM of hispidin for 48 h as determined by western blotting was compared with that of non-treated cells. Interestingly, the expression of tumor-suppressor p53, cleaved caspase-3, and cleaved PARP was up-regulated with increasing concentration of hispidin, while the expression of NF-κB and BCL2 was down-regulated (Figure 1C). Furthermore, western blotting showed that hispidin down-regulated the expression of stemness markers NANOG and SOX2 and the CSC marker CD44 in a dose-dependent manner (Figure 1C). These results together suggest that hispidin suppresses cell stemness, NF-κB activation, and proliferation of pancreatic cancer cells, thereby inducing apoptosis.
Hispidin-induced apoptosis attenuates the proliferation of BxPC-3 CD44+ pancreatic CSCs. To further investigate the role of hispidin as an anticancer agent, its effects on pancreatic CSCs was assessed. Firstly BxPC-3 CD44+ pancreatic CSCs were separated as a subpopulation of cells from BxPC-3 cells. The proportion of CD44+ BxPC-3 cells was 90%. The effects of hispidin on cell viability, apoptotic cell population, and cell-cycle stages of CSCs were then analyzed. In response to hispidin treatment for 24 h, the cell viability of BxPC-3 CD44+ cell decreased in a dose-dependent manner. Continuous treatment with the same concentration of hispidin for a longer period (48 h) increased the effect of hispidin. Interestingly, the reduction in cell viability in response to hispidin treatment for 72 h was similar to that after 48 h (Figure 2A and B). Furthermore, the MTT assay showed that 150 μM hispidin was sufficient to suppress the proliferation of pancreatic CSCs after treatment for 48 h (Figure 2C).
Apoptosis has been implicated as one of the mechanisms by which hispidin inhibits cell proliferation (17). Increased apoptotic cell populations were observed with increasing concentrations of hispidin treatment for 48 h. Intriguingly, FACS analysis with Annexin V staining and DCF-DA assay revealed that 150 μM hispidin treatment for 48 h was sufficient to induce pancreatic CSC apoptosis, thereby suppressing cell proliferation (Figure 2D and E). The cell-cycle profile of cells treated with 150 μM hispidin for 48 h was also examined and it was found that the number of S-phase cells was reduced with an increase in the number of G1 phase cells and reduction in the number of G2 phase cells (Figure 2F). These findings suggested that the reduction of BxPC-3 CSC proliferation likely resulted from hispidin-induced G1 arrest and cell apoptosis, and it clearly suggested that hispidin exerts cytotoxic effects on pancreatic CSCs.
Hispidin reduces the stemness, migration, and self-renewal ability of CD44+ BxPC-3 pancreatic CSCs. To better understand the cytotoxic effects of hispidin on cancer stem-like properties of pancreatic CSCs, more detailed in vitro analyses were performed in CD44+ BxPC-3 cells treated with 150 μM hispidin for 48 h. A colony-formation assay showed that hispidin treatment significantly reduced the numbers of BxPC-3 CD44+ colonies, indicating the inhibition of cell proliferation by hispidin treatment (Figure 3A). A transwell assay with Matrigel and without Matrigel indicated that hispidin reduced the populations of migrating and invading CD44+ BxPC-3 cells (Figure 3B).
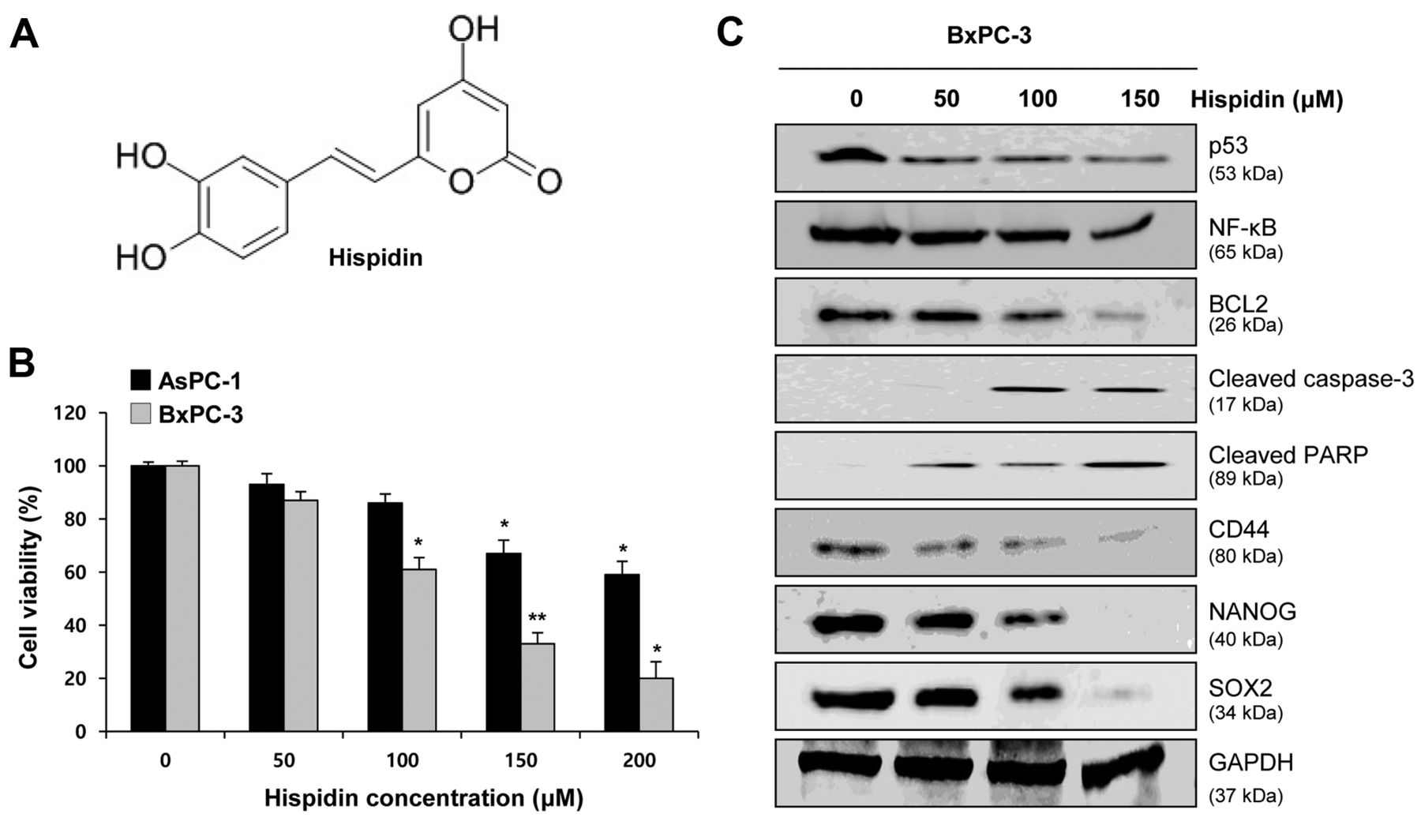
Effect of hispidin on pancreatic cancer cell proliferation and stemness. A: Two-dimensional structure of hispidin. B: 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay revealed a dose-dependent reduction of viability of hispidin-treated AsPC-1 and BxPC-3 pancreatic cancer cells. C: Western blotting showed dose-dependent effects of hispidin on tumor-suppressor p53; apoptosis-related proteins B-cell lymphoma 2 (BCL2), cleaved caspase-3, and cleaved poly (ADP-ribose) polymerase (PARP); cancer stem cell marker CD44; and stemness markers NANOG and Sex-determining region Y-box 2 (SOX2) in BxPC-3 cells. Data represent the mean±SEM (n=5 per group). Significantly different from untreated cells at *p<0.05, and **p<0.01. NFκB: nuclear factor-kappa B; GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
The results of immunocytochemistry assays showed that hispidin reduced the stemness of CD44+ BxPC-3 cells by lowering the expression levels of stemness markers NANOG and SOX2 and the pancreatic CSC marker CD44 in cells treated with hispidin (150 μM) for 48 h compared to the corresponding levels in untreated cells (Figure 3C and D). A western blot assay further showed that reduction of stemness was depended on hispidin concentration (Figure 3E). In the sphere-formation assay and sphere immunocytochemistry assay, treatment with 150 μM hispidin for 48 h significantly reduced the size, number, and CD44, NANOG and SOX2 expression of spheres, indicating the reduction of self-renewal ability (Figure 3F and G). Taken together, these findings show that hispidin negatively regulates stemness of pancreatic CSCs.
Hispidin potentiates the therapeutic efficacy of gemcitabine in pancreatic CSCs by inhibiting stemness. Inhibition of the stemness in pancreatic cancer stem cells was reported to enhance the efficacy of gemcitabine-based chemotherapy (18). In our study, stemness of pancreatic CSCs was reduced by hispidin treatment. Therefore, we investigated whether hispidin synergizes the effects of gemcitabine thereby increasing the therapeutic efficacy of gemcitabine against pancreatic CSCs.
An MTT assay revealed a significant reduction in cell viability by gemcitabine when used in combination with hispidin compared to that when either treatment was used singly (Figure 4A and B). All treatments were conducted for 48 h. To further study the mechanisms underlying the chemosensitizing effect of hispidin on gemcitabine, the expression of NF-κB, apoptosis-related proteins, stemness markers, and the pancreatic CSC marker CD44 was determined by western blotting. As shown in Figure 4C, the expression of stemness markers NANOG and SOX2, the CSC marker CD44, BCL2, and NF-κB was significantly suppressed and the expression of cleaved caspase-3 was up-regulated by combination treatment with gemcitabine and hispidin. Reduced expression of stemness markers and CD44 was confirmed in the immunocytochemistry assay (Figure 4D and 4E).
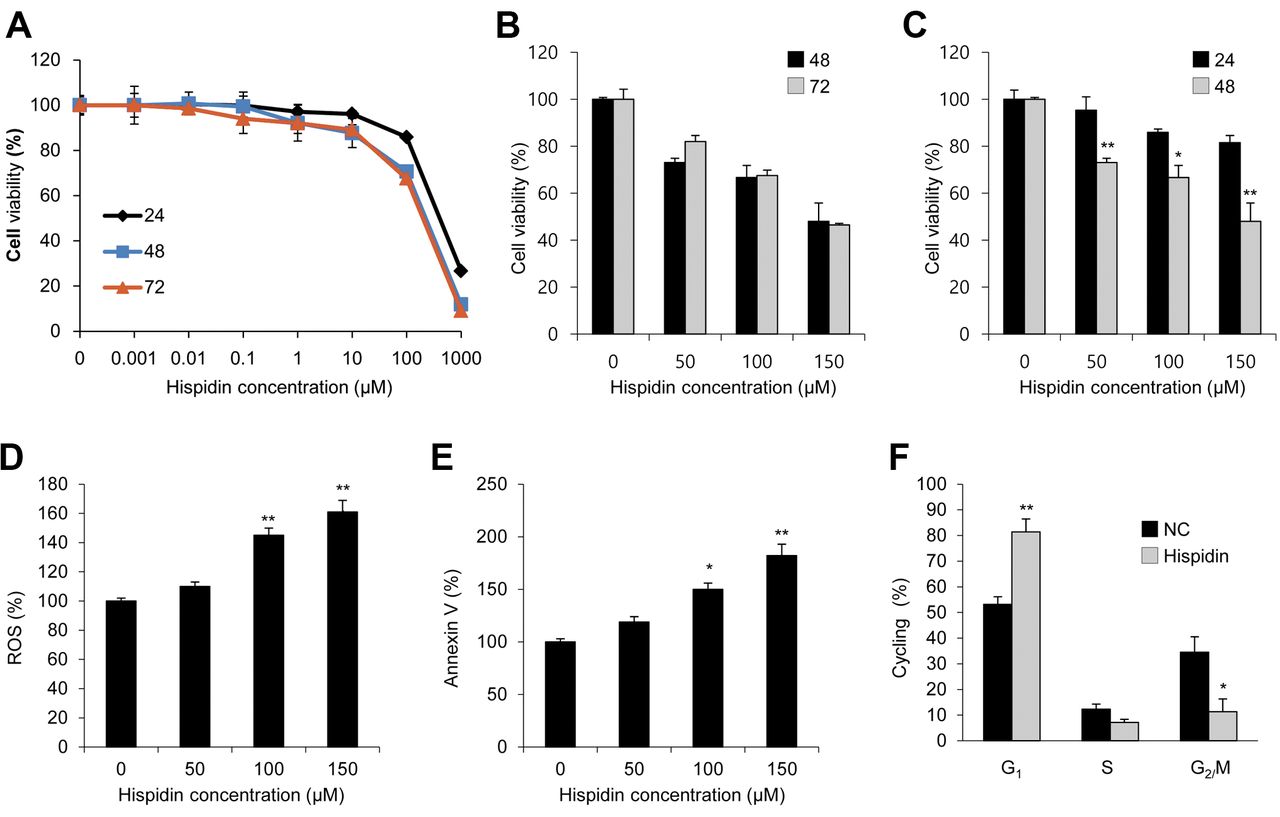
Hispidin induces cell apoptosis and reduces the viability of pancreatic cancer stem cells (CSCs). A: 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay showed dose-dependent (0–1000 μM) reduction in the viability of hispidin-treated CD44+ BxPC-3 cells. The reduction in viability in response to the treatment at 48 h was greater than that at 24 h but was similar to that at 72 h. B, C: Cell viability in response to dose-dependent hispidin treatment for 24, 48, and 72 h. D: 2,7-Dichloro-dihydro-fluorescein diacetate (DCF-DA) assay demonstrated that hispidin increased reactive oxygen species (ROS). E: Hispidin-induced apoptosis as shown by fluorescence-activated cell sorting analysis with annexin V staining. F: Cell-cycle arrest shown by cell-cycle distribution study. Data represent the mean±SEM (n=5 per group). Significantly different from untreated cells at *p<0.05, and **p<0.01.
These results showed the enhanced therapeutic effects of the combination treatment compared to that when either treatment was used singly. Taken together, these in vitro results showed that hispidin exerted cytotoxic effects and synergistically enhanced the therapeutic efficacy of gemcitabine by increasing the chemosensitivity of pancreatic CSCs through abrogation of stemness.
Discussion
In this study, we describe the cytotoxic activity of a natural phenolic compound, hispidin, extracted mainly from the medicinal mushroom P. linteus. Hispidin may be beneficial for patients with pancreatic cancer when used as a combination therapy with gemcitabine. An in vitro analysis of cellular mechanisms suggested that hispidin increased the sensitivity of pancreatic CSCs to gemcitabine via inhibiting stemness.
NF-κB is a crucial transcription factor that is expressed ubiquitously in pancreatic cancer cells. It is involved in cellular functions such as cell–cycle control, stress adaptation, inflammation, and apoptosis. It also induces tumorigenesis, invasion, metastasis, and resistance to anticancer therapies and plays a major role in oncogenesis (19, 20). It has been reported that inhibition of NF-κB, which induces resistance of pancreatic cancer cells to anticancer agents such as gemcitabine and paclitaxel (21), enhances the antitumor effects of chemotherapy (12, 22). Moreover, the down-regulation of stemness genes such as NANOG and SOX2 is also involved in the inhibition of tumorigenicity, metastasis and chemoresistance (23). In the present study, we showed that hispidin exerts cytotoxic effects against BxPC-3 and AsPC-1 pancreatic cancer cell lines by inhibiting stemness and NF-κB. The results indicated that hispidin induced BxPC-3 cell apoptosis, thereby reducing cell proliferation and cancer stemness. NF-κB plays a significant role in regulating the pancreatic CSC population, which was recently identified as candidate minor cell population responsible for resistance to chemotherapy and radiation therapy (24). Pancreatic CSCs are generally identified as CD44+/CD24+/epithelial-specific antigen (ESA+) cells. These antigens determine the characteristics of CSCs. Stemness of CD44+ pancreatic CSCs is involved in the resistance to chemotherapy and radiation therapy. Moreover, CD44 has been indicated as a marker of poor prognosis of pancreatic cancer (25-28). Interestingly, we found that hispidin inhibited stemness markers and NF-κB in BxPC-3 CD44+ cells. We also observed that hispidin had the ability to inhibit BxPC-3 CSC proliferation by inducing apoptosis and cell-cycle arrest, thereby indicating the effects of hispidin against pancreatic CSCs. Further in vitro assays showed that hispidin diminished colony-forming ability, self-renewal ability and metastasis of pancreatic CSCs.

Inhibition of pancreatic cancer stem-like properties by hispidin treatment. A: Colony-formation assay showing the effect of 150 μM hispidin on BxPC-3 cancer stem cell (CSC) proliferation. B: Effect of hispidin on cell mobility was assessed by transwell migration and invasion assays. C, D: Immunocytochemistry assay showed 150 μM hispidin suppressed the stemness of CD44+ BxPC-3 CSCs in a dose-dependent manner through reduction of expression of stemness markers. E: Western blotting confirming findings shown in C and D. F, G: Sphere-forming assay and sphere immunocytochemistry assay revealed that hispidin reduced the self-renewal ability of pancreatic CSCs. Data represent the mean±SEM (n=5 per group). SOX2: Sex-determining region Y-box 2; GAPDH: glyceraldehyde 3-phosphate dehydrogenase; DAPI: 4’,6-diamidino-2-phenylindole staining.

Hispidin enhances the sensitivity of CD44+ BxPC-3 stem cells to gemcitabine by inhibiting stemness. A: Viability of cells treated with the combination of hispidin and gemcitabine was significantly reduced in a dose-dependent manner compared to cells subjected to either treatment alone. B: Cell viability of CD44+ BxPC-3 CSCs. The concentrations of hispidin and gemcitabine used were 150 μM and 1 μM, respectively; the combination therapy of hispidin and gemcitabine was performed for 48 h. C: Western blotting showed significantly reduced expression of stemness markers, the CD44 stem cell marker, and apoptosis-related proteins by synergistic treatment. Immunocytochemistry assays confirmed a greater reduction in D: CD44 and E: NANOG and Sex-determining region Y-box 2 (SOX2) expression after the combination treatment than that after either treatment alone. Data represent the mean±SEM (n=5 per group). Significantly different from untreated cells at *p<0.05, and **p<0.01. NFκB: nuclear factor-kappa B; BCL2: B-cell lymphoma 2; GAPDH: glyceraldehyde 3-phosphate dehydrogenase; DAPI: 4’,6-diamidino-2-phenylindole staining.
NF-κB plays a critical role in the development and progression of chemoresistance to existing drugs for pancreatic cancer, including gemcitabine, a standard chemotherapeutic agent. Accumulating evidence strongly suggests that agents that can block NF-κB may reduce chemoresistance to gemcitabine and may serve as novel synergistic therapeutic regimens in combination with gemcitabine for pancreatic cancer (29-31). Furthermore, natural products can play an important role as chemosensitizers in the down-regulation of stemness, thereby inhibiting the chemoresistance of pancreatic cancer cells to existing drugs (22, 32). Similarly, our results showed that hispidin acts as a chemosensitizer of gemcitabine by down-regulating NF-κB expression and stemness synergistically with gemcitabine.
Taken together, the results of this study indicate that hispidin has the potential to exert cytotoxic effects against pancreatic CSCs alone and synergize therapeutic efficacy of gemcitabine by inhibiting stemness when used in combination. Although this was an in vitro study, the results provide evidence for the need to conduct more in-depth investigations to understand the mechanisms associated with combined effects of gemcitabine and hispidin, especially with regard to effects on stemness.
Acknowledgements
The Authors gratefully acknowledge screening of natural products for drug discovery support from 2017 Research and Education (R&E), Jeju Science High School (Students: Seok Won Jeong, Da Hyun Kang, Tae Woo Lee, Ji Min Han and schoolteacher: Woo Yong Jin), Jeju, Republic of Korea. This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2016R1A6A1A03012862)
Footnotes
Conflicts of Interest
The Authors declare no conflict of interest in regard to this study.
- Received May 9, 2018.
- Revision received May 31, 2018.
- Accepted June 4, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.