


Abstract
Background/Aim: Flavokawain B (FKB), is a natural chalcone isolated from kava root that induces apoptosis in cancer cells. Herein we investigated the effects of combination of FKB and daunorubicin (DNR) on human leukemic cells. Materials and Methods: Cell viability and death were assessed by the MTS assay and flow cytometry. NK-κB was detected by western blotting. Results: FKB alone and in combination with DNR reduced the viable cell numbers of four leukemic cell lines. FKB itself induced apoptosis of an acute myeloid cell line, HL-60. Because the additive effect of DNR and FKB was most obvious in HL-60 cells, subsequent experiments were performed with HL-60 cells. Combined treatment of the two compounds increased NF-κB activation at 12 h. Conclusion: A combination treatment of DNR and FKB may improve the anticancer effects of DNR in DNR-resistant acute myeloid leukemia.
Acute myeloid leukemia (AML) is characterized by an abnormal accumulation and rapid growth of white blood cells that interferes with the production and maturation of normal white blood cells in the bone marrow (1, 2). Approximately 13,780 people were diagnosed with AML in 2012 in the United States alone, with 10,200 patients estimated to die of the disease (3). Multidrug resistance is found in 33% of younger AML patients (age <56 years) and in 57% of older patients (age >75 years old) (4). Standard therapies and regimens confer an average survival of less than 12 months for adult AML patients (1, 2).
Daunorubicin (DNR) (also known as daunomycin), a member of the anthracycline family of compounds, originally isolated from Streptomyces peucetius, is standard chemotherapy for AML patients. DNR forms a complex with DNA base pairs and inhibits DNA synthesis by stabilizing the topoisomerase II-DNA complex and preventing relegation (5, 6). Anthracycline-based drugs induce immune-stimulatory stress pathways linked to autophagic stress, endoplasmic reticulum stress, lysosomal secretion of ATP, immunogenic stimulation by High mobility group box 1 (HMGB1) secretion, and stimulation of type-1 interferon responses (7). DNR is also used in conjunction with other therapeutic agents, such as cytarabine and mitoxantrone. However, their effects differ according to tumor type, severity, and mutational profile.
Because one of the major issues in the AML field in recent years has been relapses and drug resistance, new drugs are currently being developed to inhibit signaling pathways and proliferation of AML cells (8-12). Novel drugs may alter signaling pathways to overcome resistance, leading to cell death and growth inhibition (13, 14). Nonetheless, these signaling molecules and their interactions are complex and diverse. Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway is often activated in leukemic cells (15, 16). Tumor necrosis factor (TNF), a pro-inflammatory cytokine, is responsible for elevated NF-κB (17). Constitutive activation of signaling molecules has been seen in a variety of leukemia blasts (18, 19).
Flavokawain B (FKB), (E)-1-(2-hydroxy-4,6-dimethoxyphenyl)-3-phenylprop-2-en-1-one, is a natural chalcone that is isolated from kava (Piper methysticum) root extract and Alpinia pricei Hayata (20). FKB significantly hinders the growth of bladder, colon, and ovarian cancer as an anti-proliferative and pro-apoptotic agent (20-22). It activates mitochondrial apoptosis through up-regulation of reactive oxygen species (ROS) generation and activation of caspase-9, -3 and -8 (23). It down-regulates BCL-2 by inducing Bax activation (23). Others studies have reported loss of cell viability, morphological changes, genomic DNA fragmentation and sub-G1 phase accumulation in sarcoma and colon cancer cells (20, 23). To provide novel therapeutic regimens and overcome drug resistance, we investigated the antitumor effect of FKB in combination with DNR on human leukemic cell lines and the underlying molecular mechanisms of action, in particular, their effect on NF-κB signaling pathway.
Materials and Methods
Reagents. Pure FKB (purity >97%) was purchased from Abcam (Cambridge, UK), dissolved in DMSO at 25 mM, and stored at − 20°C. DNR hydrochloride was purchased from Tocris (Bristol, UK), dissolved in distilled water at 10 mM, and stored at −20°C.
Cell culture. HL-60, an acute promyelocytic leukemia cell line, K562, a chronic myelogenous leukemia cell line, and MOLT4 and CEM1, acute lymphoblastic leukemia cell lines (all from Korean Cell Line Bank, Seoul, Korea) were maintained in a 37°C incubator in a humidified atmosphere containing 5% CO2. Cells were cultured in RPMI 1640 medium (WelGene, Gyeongsangbuk-do, Republic of Korea) supplemented with 10% (v/v) heat-inactivated fetal bovine serum (Capricorn Scientific, Ebsdorfergrund, Germany), penicillin-streptomycin (Cellgro, Manassas, VA, USA), 5 mM sodium pyruvate (Pan Biotech, Aidenbach, Germany), and 55 μM 2-mercaptoethanol (Thermo Fisher Scientific, Waltham, MA, USA).
Cell survival assay. HL-60 and K562 cells were seeded in 96-well flat-bottom plates at 5000 cells/well and MOLT4 and CEM1 cells were seeded in 96-well flat-bottom plates at 8000 cells/well. Cells were treated with FKB, DNR, or DMSO as vehicle control at the designated concentrations and incubated for 2-3 days. Each well was treated with 20 μl of the CellTiter96®One Solution Cell Proliferation Assay (Promega, Madison, WI, USA) for 3 hours at 37°C in a humidified atmosphere containing 5% CO2. One Solution Reagent contains the tetrazolium compound, 3-(4,5-dimethylthiazol-2-yl)-5-3(-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS). Absorbance was measured at 490 nm in a Sunrise ELISA reader (Tecan, Mannedorf, Switzerland).
Annexin V/propidium iodide cell death assay. Cell death was analyzed using an Annexin V (AV) and propidium iodide (PI) assay. HL-60 cells were placed at 2×105 cells/tube in FACS tubes and were treated with 1% DMSO as vehicle control, FKB (10 μM), DNR (0.15 μM), or FKB+ DNR. The cells were cultured at 37°C in a humidified atmosphere containing 5% CO2. After 16 hours, 3 ml of 1× AV binding buffer was added to each sample tube and the tubes were centrifuged at 1,500 rpm for 5 min. The supernatant was discarded. After vortexing, 5 μl AV-FITC (BD Biosciences, Franklin Lakes, NJ, USA) and 4 μl PI (BioLegend, San Diego, CA, USA) were added and the tube was incubated for 15 minutes on ice in the dark. Then, 200 μl of 1×AV binding buffer was added. Fluorescence was measured using flow cytometry through a FACS Canto II (BD Biosciences) and analyzed using FlowJo software (FlowJo LLC, Ashland, OR, USA).
Western blotting. Expressions of p-p65, p-65, α-tubulin, and lamin B were detected by western blotting. After incubation with the designated drug(s), cells were harvested and washed twice with phosphate-buffered saline (PBS). The pellets were collected after centrifugation at 1500 rpm at 24°C for 5 min. The volume of the lysis buffer used was three time the volume of the cell pellet and was incubated for 15 minutes at room temperature. The lysis buffer contained 0.1% SDS (Elpis Biotech, Daejeon, Korea), 1× protease inhibitor (Roche, Basel, Swiss), 5 mM NaF (Sigma-Aldrich, St. Louis, MO, USA), and 2 mM Na3VO4 (Sigma-Aldrich) in NP-40 protein extraction solution (Elpis Biotech). Nuclear and cytosolic fractions were separated using a NE-PER® Nuclear and Cytoplasmic Extraction kit (Thermo Fisher Scientific). Total protein of 15 μg per sample was electrophoresed in 10% SDS-PAGE gels and transferred to polyvinylidene fluoride (PVDF) membranes (GE Healthcare, Pittsburgh, PA, USA). Then, blots were blocked with 3% BSA (Bovogen, East Keilor, Australia) in 1× PBS containing 0.1% Tween 20 (PBST) for 30 minutes and incubated with the following antibodies in a 4°C refrigerator overnight: anti-p-p65 (Ser536) (polyclonal; Santa Cruz, Dallas, TX, USA), anti-p65 (clone F-6; Santa Cruz), anti-α-tubulin (clone B-7; Santa Cruz), and anti-lamin B (clone C-20; Santa Cruz). The membranes were washed three times with TBST and incubated with secondary antibody for 1hour at room temperature. Secondary antibodies were as follows: goat anti-mouse, goat anti-rabbit, and donkey anti-goat (all from Santa Cruz). Finally, protein bands were detected using PicoEPD™ Western Reagent solutions A and B (1:1) (Elpis Biotech) and ImageQuant LAS4000 (GE Healthcare) and analyzed using Image Studio Lite software Ver 5.2 (LI-COR Bioscience, Lincoln, NE, USA).
Statistical analysis. Results are expressed as the mean±standard error of the mean (SEM). Statistical analyses were performed using Tukey-Kramer multiple comparisons tests (parametric ANOVA) for MTS assays. The Kruskal-Wallis test (nonparametric ANOVA) was used for western blotting results (InStat3 software, GraphPad, La Jolla, CA, USA). Differences were considered statistically significant at p<0.05.
Results
FKB reduces cell numbers of human leukemic cell lines and induces apoptosis. First, we assessed cell viability at various concentrations of FKB to evaluate its cytotoxic effects and optimal concentration for further study. Four human leukemic cell lines were exposed to FKB for 48 and 72 h and tested using the MTS assay. The viable cell numbers of all four cell lines were reduced in dose-dependent manners (Figure 1A). HL-60 and K562 cells were significantly reduced by 15 μM and 24 μM FKB, respectively, at 72 h, compared vehicle control. However, the cell lines showed different susceptibilities to FKB. K562 cells appeared to be quite resistant, whereas CEM1 cells were sensitive.
The results of the MTS assays could not determine whether FKB inhibited cell proliferation or induced apoptosis or necrosis. Thus, a cell death assay was performed using PI and AV. Compared with vehicle, FKB treatment increased both early and late apoptotic cell populations in a time- and dose-dependent manner (Figure 1B). The percentages of early apoptotic (PI−AV+) cells treated with 10 μM and 15 μM of FKB at 24 h were 2.17%, and 12.9%, respectively, whereas the percentages of late apoptotic cells (PI+AV+) were 2.31% and 8.45% in the 10 and 15 μM FKB-treated groups, respectively. The dose-dependent apoptotic cell death induced by FKB was more evident at 48 h. These results show that FKB exerts an anticancer effect by inducing apoptosis, rather than necrosis.
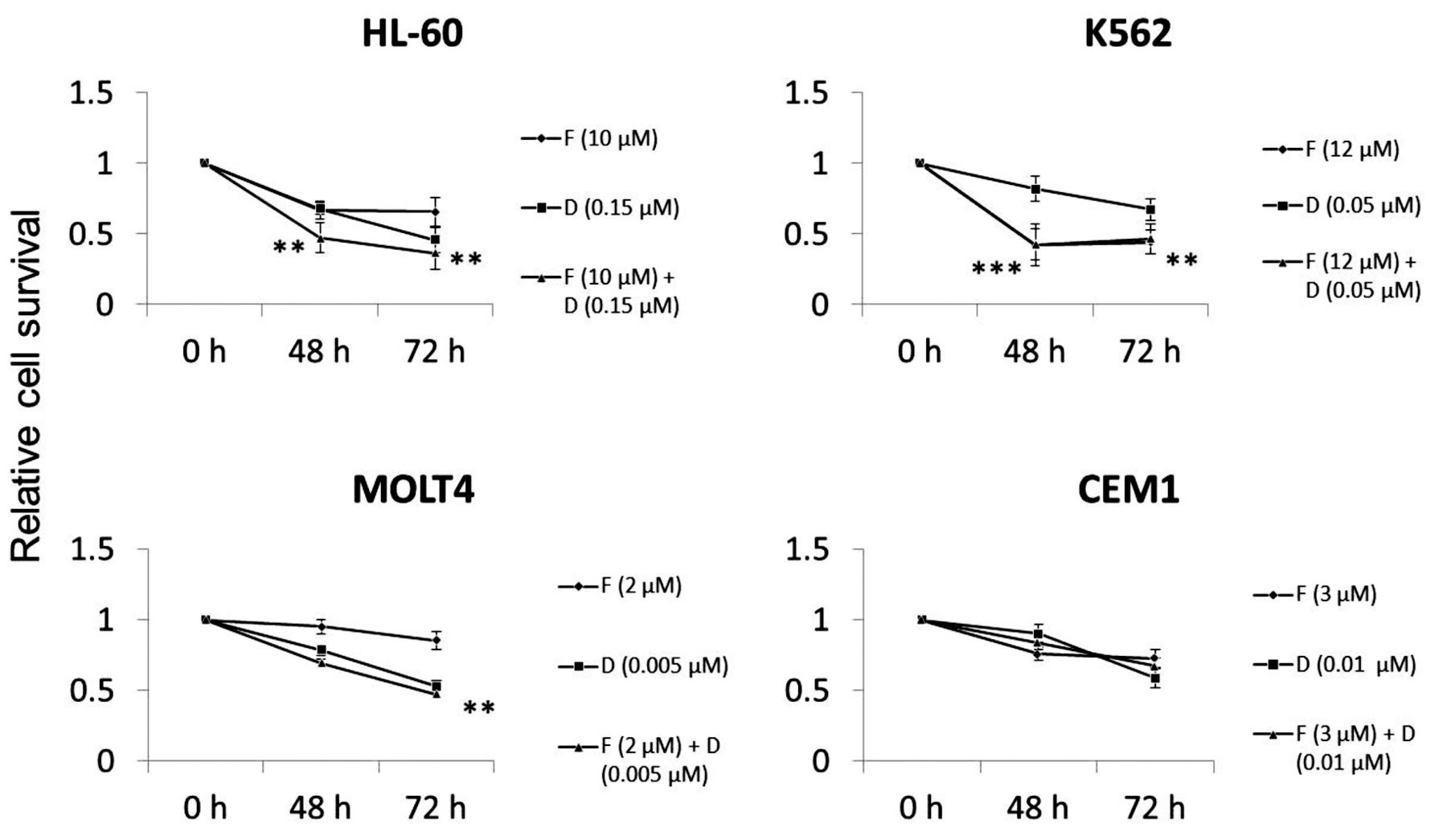
FKB in combination with DNR reduces viability of human leukemic cell lines. Next, the combined effect of FKB and DNR was evaluated. Because the aim of this study was to investigate the combination treatment of FKB and DNR, we selected the appropriate concentrations of FKB and DNR to be used for further study on the basis of the ability of FKB or DNR alone to induce moderate cell death (around 50%) (Figure 1A and data not shown). As shown in Figure 2, HL-60 cells showed the clearest additive effect of FKB and DNR. At 72 hours, the difference between the vehicle and co-treatment groups was significant (p<0.01), while the difference between the vehicle and FKB single treatment was not. HL-60 cells appeared to be the most resistant to the conventional chemotherapeutic drug DNR because they required the highest concentration of DNR to achieve moderate cell death. The additive effect of FKB and DNR was not marked in other cells. In summary, FKB alone had cytotoxic effects on human leukemic cell lines and, in combination with DNR, had an additive effect on HL-60 AML cells. To understand the mechanisms underlying the combined effect of FKB and DNR, we performed further experiments with HL-60 cells.
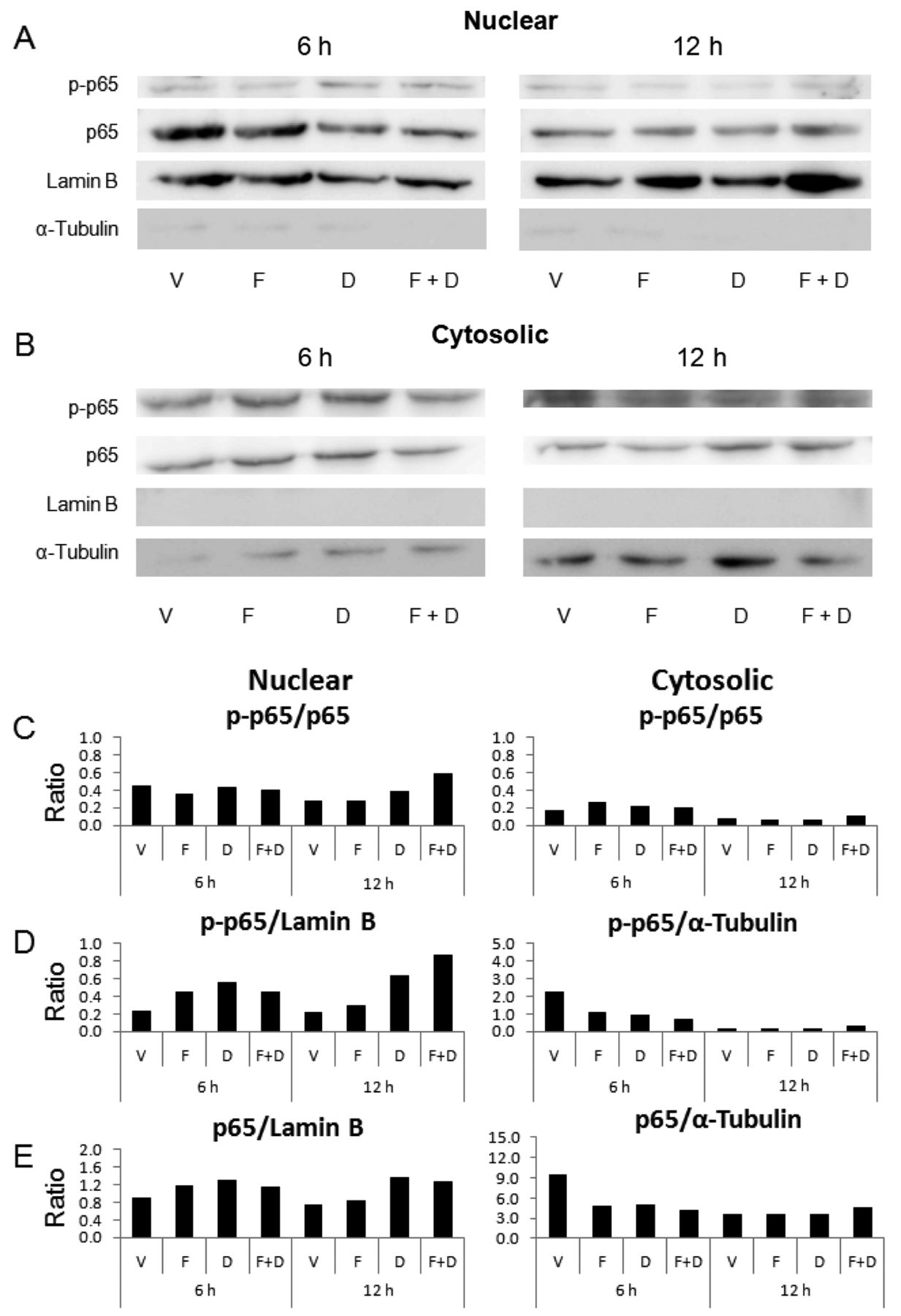
Combined treatment with FKB and DNR induces NF-κB activation. Because p65 is a functional subunit of NF-κB, changes in NF-κB activation were measured using western blot analysis of p65 phosphorylation. Early time points were evaluated because the changes in signaling molecules precede cell death. As shown in Figure 3A and B, nuclear and cytosolic proteins were fractionated because activated NF-κB is translocated to the nucleus to promote transcription. Lamin B is a nuclear marker, whereas α-tubulin is a cytosolic marker. To quantitatively evaluate the changes in p65 activation, band intensities were assessed and calculated as relative amounts to those of loading controls, that are lamin B and α-tubulin, as seen in Figure 3C-E. FKB and DNR did not reduce NF-κB activation, but rather increased it. The increases in activated NF-κB were in line with those induced in cell death by FKB, DNR, and their combination. Compared with vehicle control, DNR itself increased the relative amounts of phosphorylated p65 (p-p65) to p65 (Figure 3C) and to lamin B (Figure 3D) by 35% and 100%, respectively, at 12 hours. In particular, compared with vehicle control, the relative amount of p-p65 to total p65 was increased up to 110% and that to lamin B up to314% in the nucleus by combined treatment for 12 hours. In contrast, the relative amounts of p-p65 and p65 to α-tubulin were decreased in the cytosol, suggesting that the increase in activated NF-κB in the nucleus was due to translocation of activated NF-κB from the cytosol to nucleus, rather than de novo synthesis of NF-κB proteins. Taken together, combined treatment with FKB and DNR activated NF-κB.
Discussion
In this study, FKB, a natural product found in the kava plant, was found to kill human leukemic cell lines through apoptosis, as assessed by an MTS assay and PI/AV assay. HL-60 cells appear to be the most resistant to the conventional chemotherapeutic drug DNR because they required the highest concentration of DNR to induce moderate cell death. Thus, the additive effect of FKB may improve the outcomes of chemotherapy for drug-resistant AML. Notably, FKB in combination with DNR efficiently removed HL-60 cells, which are DNR-resistant AML cells, among others, although the results from other cell lines may require further investigation using various concentrations of the two reagents. HL-60 cells are multidrug resistant (24) and have three mutant genes: CDKN2A, NRAS, and TP53. The DNA methylation status of CDKN2A affects its susceptibility to doxorubicin, another anthracycline, in breast cancer (25). Up-regulation of CDKN1A inhibits apoptosis by DNR in human myeloid cells (26). A myeloid cell line expressing an activating mutation of NRAS shows resistance to doxorubicin (27). Loss of function, inactivation, or deficiency of p53 is suggested to be associated with resistance to anthracycline in breast and colorectal cancers (28-30). In the clinical situation, genetic analysis to identify drug-resistant mutations may help to optimize chemotherapeutic regimes for AML patients. An important prerequisite is the identification of all mutations and polymorphisms causing drug resistance.
Multidrug resistance is expressed in 33% of younger AML patients (age <56 years) and in 57% of older patients (age >75 year old) in the United States (4) and contributes to its high mortality. There are thus urgent unmet needs for novel therapeutic regimens for drug-resistant leukemia. Greater attention has been paid to the development of antibodies and immune cell therapies for B and T cell ALL. For example, chimeric antigen receptor (CAR)-T cells specific to CD19 have been produced for B cell ALL. However, AML is a heterogeneous disease, which complicates the identification of a single molecule as a therapeutic target. It is thus imperative to improve chemotherapy for AML. Natural products have proved very useful in the development of anticancer drugs. Out of a total of 175 small-molecule anticancer drugs introduced into therapy over a 70-year period, approximately 49% were either directly obtained from organisms or were derived from natural products (31). Daunorubicin is one such compound. Our results suggest that the outcomes of DNR-resistant AML patients might be improved, if FKB is used as a sensitizer or a supplement to DNR. The combination of FKB with other chemotherapeutics may also improve its cytotoxic effects and help overcome drug resistance.

FKB eliminates human leukemic cells by apoptosis. A. FKB reduces viable cell numbers of human leukemic cell lines. HL-60, K562, MOLT4, and CEM1 cells were exposed to the designated concentrations of FKB for 0, 48, and 72 h. Cell viability was assessed by an MTS assay. The results displayed are the relative cell viability to that of the vehicle control (V). When the results are statistically significant compared with vehicle controls at 72 hours, the results are marked in the figure. In HL-60 cells at 48 h, **V vs. F 15 μM, *F 5 μM vs. F 15 μM; and at 72 h, **V vs. F 15 μM, *F 5 μM vs. F 15 μM. In K562 cells at 72 h, *V vs. F 24 μM, *F 3 μM vs. F 24 μM, and *F 6 μM vs.F 24 μM. In CEM1 cells at 48 h, ***V vs. F 5 μM, ***F 1 μM vs. F 5 μM, *F 2 μM vs. F 3 μM, ***F 2 μM vs. F 5 μM, and *F 3 μM vs. F 5 μM; and at 72 hours, *V vs. F 3 μM, ***V vs. F 5 μM, **F 1 μM vs. F 5μM, and **F 2 μM vs. F 5 μM. *p<0.05, **p<0.01, ***p<0.005. Data are means±SEM. N=4 for HL-60, 2 for K562, 4 for MOLT4, and 2 for CEM1. Each experiment was performed as three independent experiments with six replicates each. B. FKB promotes apoptosis in HL60 cells. HL-60 cells were treated with 10 or 15 μM FKB for 24, and 48 h and then the apoptotic cells were assessed using FITC-conjugated AV and PI by flow cytometry. Ethanol was used as a positive control for the last 30 min while the negative control was not treated with any reagent. The results are representative of at least 3 independent experiments.

Co-treatment of FKB with DNR had additive effects on HL-60 cells. HL-60, K562, MOLT4, and CEM1 cells were treated with the drugs as follows: FKB alone, DNR alone, and their combination. Cell viability was assessed by an MTS assay. The results displayed are the relative cell viability to vehicle control (V). In the figure, only statistically significant differences between V and F+D groups are displayed. In HL-60 cells at 48 h, *V vs. F 10 μM, **V vs. F 10 μM + D 0.15 μM; and at 72 h, **V vs. D 0.15 μM, **V vs. F 10 μM + D 0.15 μM. In K562 cells at 48 h, ***V vs. F 12 μM, ***V vs. F 12 μM + D 0.05 μM, *F 12 μM vs. D 0.05 μM, D 0.05 μM vs. F 12 μM + D 0.05 μM; and at 72 hours, ***V vs. F 12 μM, **V vs. D 0.05 μM and V vs. F 12 μM + D 0.05 μM. In MOLT4 cells at 72 hours, **V vs. D 0.005 μM, **V vs. F 2 μM + D 0.005 μM and F 2 μM + D 0.005 μM. *p<0.05, **p<0.01, ***p<0.005. Data are means±SEM. N=3-4 for HL-60, 3-8 for K562, 2-9 for MOLT4, and 9 for CEM1. Each experiment was performed as three independent experiments with six replicates each. V: Vehicle; F: flavokawain B; D: daunorubicin; F+D: flavokawain B + daunorubicin.
To understand the mechanisms underlying the additive effect of FKB and DNR, we investigated NF-κB activation that is involved in the pathogenesis of AML and usually overactivated in AML patients (32-34). Therefore, it was initially expected that their activation status would be down-regulated by FKB and/or DNR. DNR indeed reduced p-AKT and p-ERK at 6 h, but the reduction was not maintained until 12 h (data not shown). DNR increased p-p65 at 12 h, as previously described (35), and increased it more in combination with FKB. The induction in activated NF-κB was concomitant with that of cell death by FKB, DNR, and their combination. Some reagents, such as SMAC mimetic and nicotine, also activate the NF-κB pathway, resulting in apoptosis (36, 37). DNR is suggested to increase p-ERK in U937 human leukemic cells at 30 min and in K562 cells at 24 h (38, 39). Taken together, over-activated signaling molecules could be accompanied with cell death.
Cytotoxicity due to DNR is a result of intercalation-induced distortion of the double helix or stabilization of the cleavable complex formed between DNA and topoisomerase II, inhibiting DNA synthesis (40). It is also suggested that ROS are generated through the direct interaction of DNR with cell membranes and subsequent lipid peroxidation. FKB induces G2/M accumulation and ROS generation in human colon cancer cells and activation of caspase-9, -3, and -8 (23). It further down-regulates BCL-2 and increases Bax activation (23). FKB inhibits the growth of and induces apoptosis of CCRF-CEM, RS4-11, and Jurkat acute lymphoblastic leukemia cells by increasing the expression of p53 (41). These reports support our current results showing that FKB killed AML cells by apoptosis. Taken together, the additive effect of FKB and DNR might be the result of cell cycle inhibition at the DNA synthesis and mitosis phases, as well as increased ROS production by both reagents.

Combined treatment with FKB and DNR increased phospho-p65 in the nucleus. (A and B) HL-60 cells were treated with vehicle (1:1,000 DMSO), FKB (10 μM), or DNR (0.15 μM) for 6 and 12 h. The expression levels of phospho-p65 (p-p65), p65, lamin B, and α-tubulin were determined by western blot analysis. Lamin B and α-tubulin were used as loading controls for nuclear (A) and cytosol (B) fractions, respectively. The results are representative of three independent experiments. (C-E) The graphs show the relative expression of p-p65 to p65 (C, left panel), p-p65 to lamin B (D, left panel), and p65 to lamin B (E, right panel) in the nucleus, and p-p65 to p65 (C, right panel), p-p65 to α-tubulin (D, right panel), and p65 to α-tubulin (E, right panel) in the cytosol. The ratios were calculated from the band intensities measured by densitometry and are the means of three independent experiments. V: Vehicle; F: flavokawain B; D: daunorubicin; F+D: co-treatment of F and D.
In conclusion, our present findings suggest that combination treatment of DNR and FKB may improve the anticancer effects of DNR in patients with DNR-resistant leukemia. FKB induces apoptosis, presumably through changes in NF-κB activation. However, further investigation is required on the effect of FKB itself and its mechanisms of action with more elaborate experimental schemes, along with preclinical studies.
Acknowledgements
This study was supported by a grant from the National R&D Program for Cancer Control, Ministry of Health & Welfare, Korea (1520060).
Footnotes
↵* These Authors contributed equally to this work.
- Received February 27, 2018.
- Revision received March 21, 2018.
- Accepted March 26, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}