


Abstract
Background/Aim: In a previous study, we showed that amentoflavone promotes sorafenib-induced apoptosis in hepatocellular carcinoma (HCC) cells in vitro. However, whether amentoflavone augments anticancer efficacy of sorafenib in HCC in vivo is unknown. The aim of the present study was to verify the anticancer effect of amentoflavone combined with sorafenib in HCC in vivo. Materials and Methods: HCC SK-Hep1 tumor-bearing mice were treated with vehicle, sorafenib, amentoflavone, or combination for 14 days, respectively. Effect of sorafenib, amentoflavone, or their combination on tumor growth, anti-apoptotic potential, apoptotic signaling and general toxicity were evaluated with digital caliper, immunohistochemistry staining and body weight. Results: Our results demonstrated that amentoflavone significantly enhanced sorafenib-inhibited tumor growth and expression of ERK/AKT phosphorylation and anti-apoptotic proteins compared to single-agent treatment. Additionally, amentoflavone also triggered sorafenib-induced apoptosis through extrinsic and intrinsic apoptotic pathways. Conclusion: Amentoflavone boosts therapeutic efficacy of sorafenib through blockage of anti-apoptotic potential and induction of apoptosis in HCC in vivo.
Hepatocellular carcinoma (HCC) is a prevalent cancer type and also the leading cause of cancer death among Taiwanese. Genomic alterations affecting matrix metalloproteinase-1 (MMP-1), tumor necrosis factor-alpha (TNF-α), and DNA double-strand break repair gene XRCC7 genotypes result in increased HCC risk in Taiwanese males (1-3).
Sorafenib (Nexavar), an oral multi kinase inhibitor against Raf kinase and several receptor tyrosine kinases, has been approved for the treatment of advanced hepatocellular carcinoma (HCC) (4). Sorafenib blocks Raf kinase signaling transduction and inhibits activity of vascular endothelial growth factor receptor 2 (VEGFR2) and platelet-derived growth factor receptor (PDGFR) leading to inhibition of tumor growth and angiogenesis in HCC (5). Patients with advanced HCC have an unsatisfactory response to sorafenib treatment and resultant median overall survival is less than 11 months (6). Lu et al. observed longer survival in patients receiving optimal dose of sorafenib in comparison to those receiving suboptimal dose (7). Yoshimoto et al. suggested the reasonable use of sorafenib before surgery in advanced HCC patients particularly those whose tumor responded to sorafenib (8). Therefore, development of sorafenib sensitizers which ameliorate efficacy of sorafenib against HCC is very important for advanced HCC patients under sorafenib treatment.

Flow chart of experimental protocol.
Flavonoids, extracted from natural products, Chinese herbal medicines, and phytochemicals, possess anticancer properties through blockage of multiple signal transduction pathways, initiation of apoptosis, and up-regulation of anticancer immune response (9-10). Flavonoid compounds such as wogonin (5,7 dihydroxy 8 methoxyflavone), fisetin, and quercetin are presented as potent sorafenib sensitizers to enhance anticancer activity of sorafenib in HCC, melanoma, and glioma, respectively (11-13). Flavonoids and sorafenib combination may be a potential strategy for treatment of HCC.
Amentoflavone, a flavonoid derived from scutellaria baicalensis, has been indicated to inhibit metastatic potential through suppression of NF-κB activation in breast cancer and osteosarcoma cells (14-15). In a previous study, we found that amentoflavone promotes sorafenib-induced apoptosis through extrinsic and intrinsic pathways in hepatocellular carcinoma in vitro (16). However, whether amentoflavone increases therapeutic efficacy of sorafenib in HCC in vivo is ambiguous. The aim of the present study was to verify the therapeutic efficacy of amentoflavone combined with sorafenib in SK-Hep1 HCC-bearing mice. Here, we demonstrated amentoflavone as a sorafenib sensitizer which enhances therapeutic effi-cacy of sorafenib through suppression of anti-apoptotic potential and induction of apoptosis in HCC in vivo.
Materials and Methods
Chemicals and agents. Amentoflavone and DMSO were purchased from Sigma-Aldrich (St. Louis, MO, USA). Sorafenib (Nexavar) was kindly provided from Bayer Health Care Pharmaceuticals, Inc. (Whippany, NJ, USA). Culture related products, including Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), L-glutamine and penicillin-streptomycin (PS) were obtained from Gibco/Life Technologies (Waltham, MA, USA). Immunoperoxidase secondary detection kit was bought from Millipore (Billeri-ca, MA, USA). Matrigel matrix was purchased from Corning Incorporated (Corning, NY, USA).
Cell culture. Human hepatocellular carcinoma SK-Hep1 cells were provided by professor Jing-Gung Chung at Department of Biological Science and Technology, China Medical University, (Taichung, Taiwan). Cells were cultured with DMEM containing 10% FBS, 2 mM L-glutamine, and PS (100 U/ml and 100 μg/ml) and incubated at 37°C under a humidified atmosphere of 95% air and 5% CO2 (17).
Establishment of tumor-bearing animal model. Six-week-old nude mice were brought from the National Laboratory Animal Center, Taipei, Taiwan. 1×107 SK-Hep1 cells were suspended in 150 μl mix-ture of serum-free DMEM and matrigel (2:1) and inoculated subcutaneously in the right legs of nude mice (18).

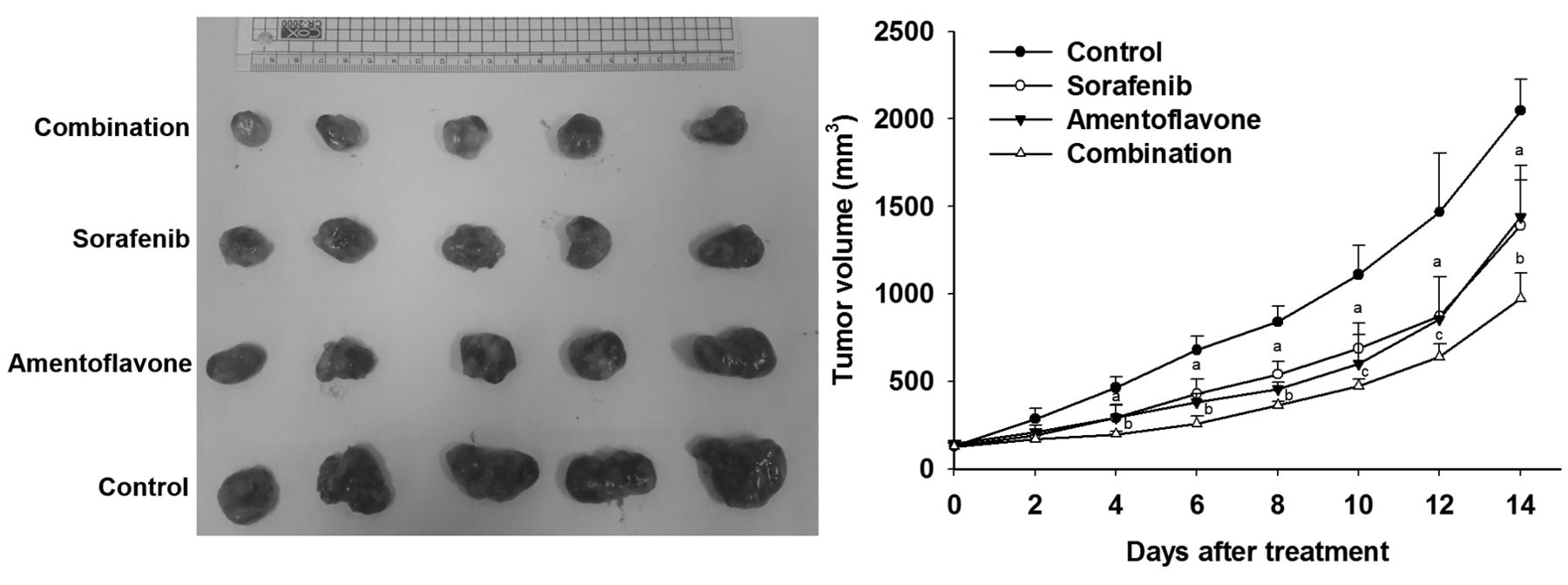
Effect of sorafenib, amentoflavone, and their combination on tumor growth in SK-Hep1 HCC bearing mice. Mice were treated with vehicle (100 μl PBS plus 50 μl DMSO per day), sorafenib (10 mg/kg/day), amentoflavone (50 mg/kg/day), or their combination (sorafenib 10 mg/kg plus amentoflavone 50 mg/kg per day) by gavage for 14 days, respectively. Tumor growth was measured by digital caliper and tumor volume was calculated using the formula 0.523 × length × width × thickness. ap<0.01 compared to vehicle group. bp<0.01 and cp<0.05 compared to sorafenib or amentoflavone group.
Experimental protocol (Figure 1). When average tumor volume reached about 130 mm3, mice were randomized into four groups (n=5/group), vehicle group [treated with 100 μl phosphate-buffered solu-tion (PBS) plus 50 μl dimethyl sulfoxide (0.1% DMSO) by gavage daily for 14 days] and sorafenib group (treated with 10 mg/kg/day by gavage for 14 days), amentoflavone group (treated with 50 mg/kg/day by gavage for 14 days), and combination group (treated with 10 mg sorafenib plus 50 mg amentofla-vone/kg/day by gavage for 14 days). Initiation of treatment on day 1, tumor growth and body weights of mice were measured on day 0, 2, 4, 6, 8, 10, 12, 14 after treatment. Mice were sacrificed on day 14 for IHC staining of tumor tissues from each group. All experiments were repeated at least three times and complied with the guidance of institutional animal care (IACUC number approval by Taipei Medical University: LAC-2016-0028).
Immunohistochemistry (IHC) staining. Mice were sacrificed on day 14 after treatment and tumors were fixed with 4% paraformaldehyde (PFA) at 4°C. Paraffin-embedded tumor tissue section with 5 μm thickness was performed by Bio-Check Laboratories Ltd. (New Taipei City, Taiwan). According to the instructions provided with the kit, IHC staining was used to determine protein levels in tumor tis-sues. The sections were immunohistochemically stained with MCL-1 (BioVision, Milpitas, CA, USA), AKT (Ser473), C-FLIP, active caspase-3 (Cell Signaling Technology, Inc., Danvers, MA, USA), XIAP, active caspase-8, and -9 (Thermo Fisher Scientific, Fremont, CA, USA) antibodies, respectively. The stained sections were scanned at 100X magnification by using the microscopy-based TissueFAXS plat-form (TissueGnostics, Vienna, Austria). Positive expression of IHC indices in tumor tissues was quantified with ImageJ software version 1.50 (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis. The difference between two groups (treatment and control group) was compared by Student's t-test. Results are showed as means±standard error. The p-value smaller than 0.05 was de-fined as statistically significant difference.
Results
Amentoflavone enhances sorafenib-inhibited tumor growth in SK-Hep1 tumor-bearing mice. Effect of amentoflavone, sorafenib, or combination treatment on tumor growth in SK-Hep1 HCC bearing mice was evaluated with digital caliper. All the treatment groups significantly reduced tumor growth as compared to that of control group. Tumor volumes of the sorafenib group were similar to that of amentoflavone group. Combination group had significantly smaller tumor volumes as compared to sorafenib or amentoflavone group on day 4-14 (Figure 2).
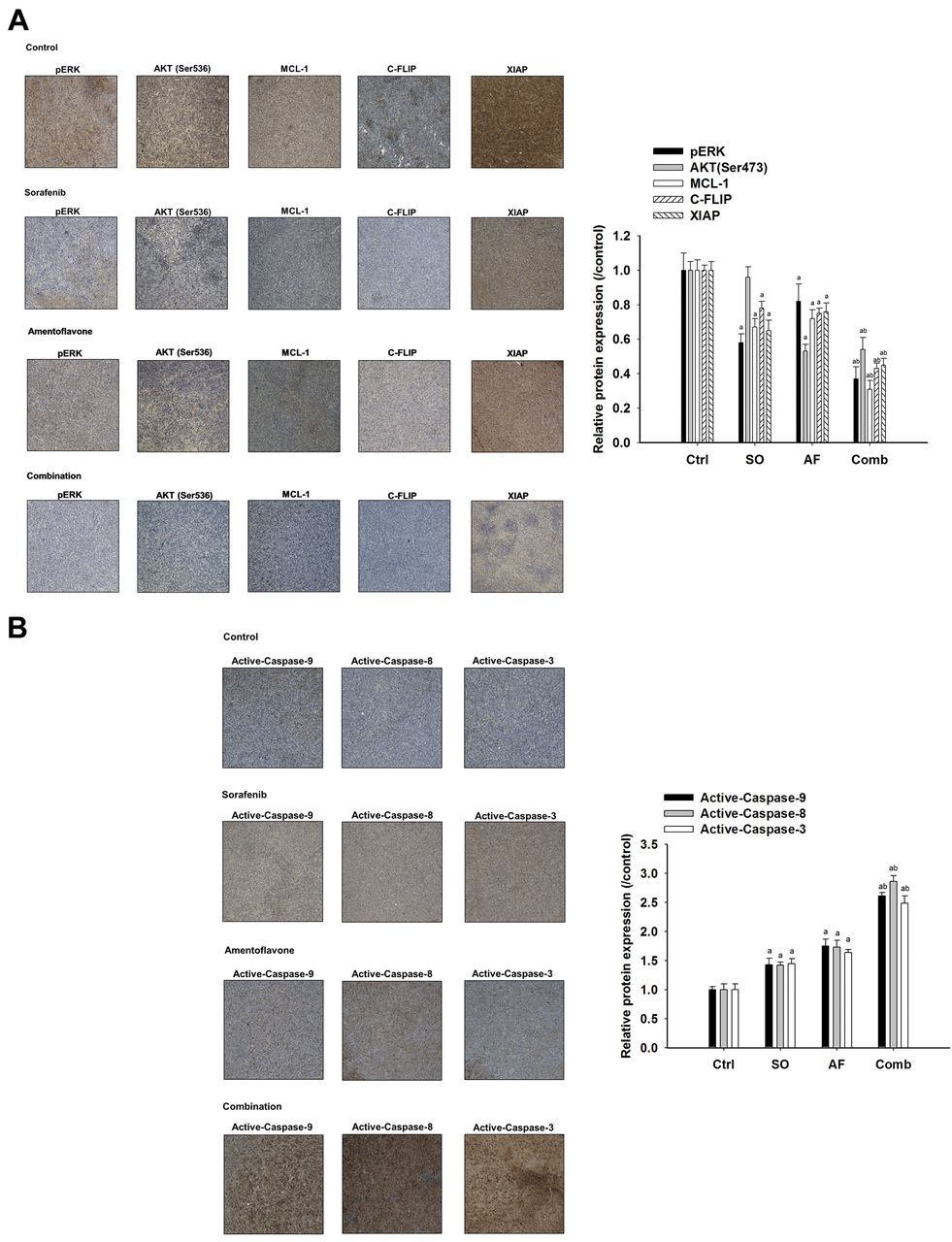
Amentoflavone promotes sorafenib-inhibited anti-apoptotic potential and triggers sorafenib-induced apoptosis in SK-Hep1 tumor bearing mice. All the treatment groups significantly decreased expression of anti-apoptotic proteins (MCL-1, C-FLIP, and XIAP) as compared to that of control group. Combination group had significantly lower protein levels of MCL-1, C-FLIP, and XIAP as compared to that of sorafenib group or amentoflavone group. Notably, the protein levels of AKT (Ser473) was not changed by sorafenib treatment. Both amentoflavone and combination group significantly inhibited protein levels of AKT (Ser473) by 57 and 56% compared to that of control, respectively (Figure 3A). All treatment groups significantly triggered activation of caspase-9, -8, and -3 compared to that of the control group. Combination group had significantly higher activation of caspase-9, -8, and -3 as compared to that of sorafenib or amentoflavone group (Figure 3B).
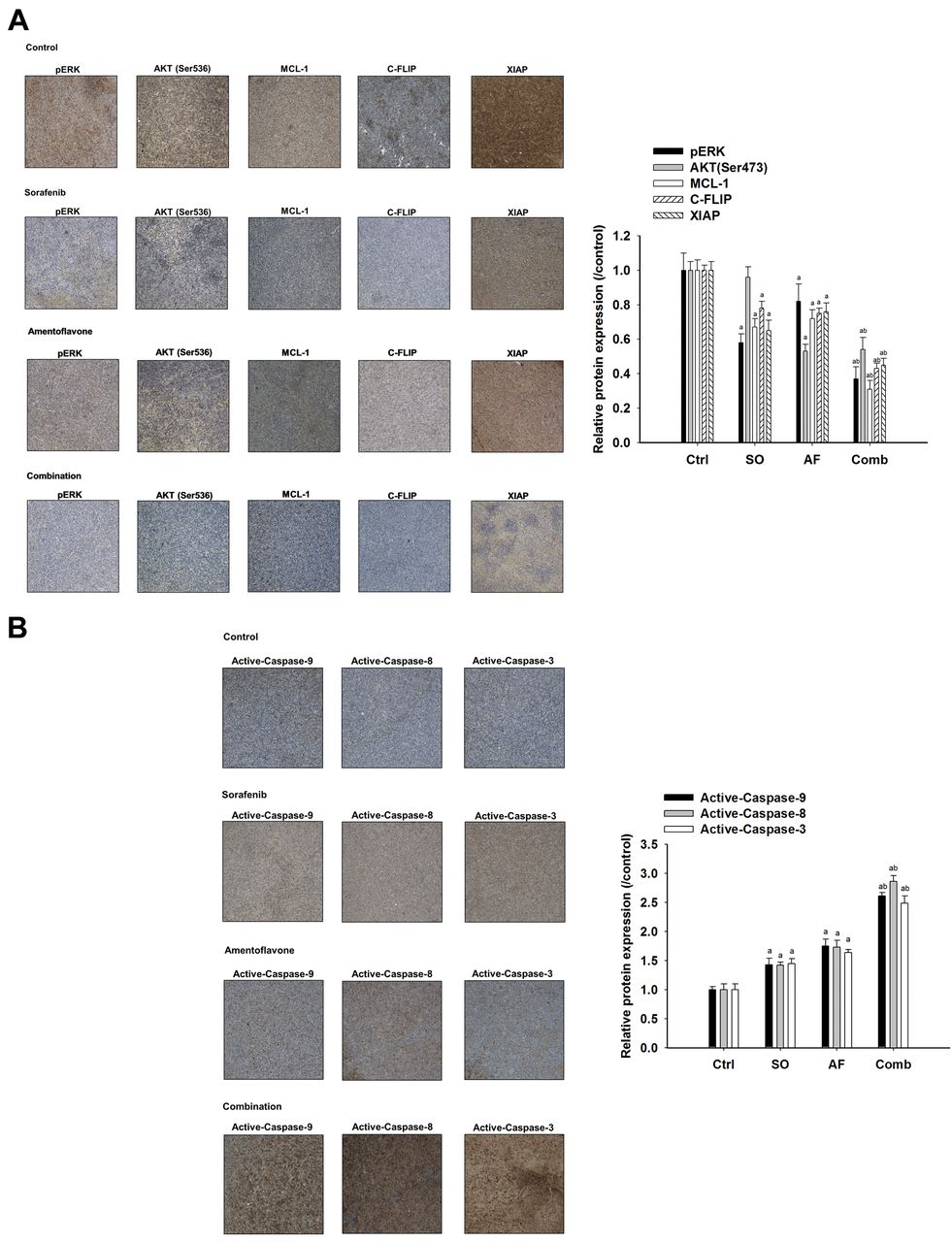
Effect of sorafenib, amentoflavone, or their combination on anti-apoptotic potential and apoptosis signaling in SK-Hep1 tumor-bearing mice. Mice were sacrificed on day 14 after treatments and protein expression of HCC tissues were investigated with IHC staining. A) Protein levels of pERK, AKT (Ser473), Mcl-1, C-FLIP, and XAIP. B) Protein expression of active-capase-9, -8, and -3. ap<0.01 compared to vehicle group and bp<0.01 compared to sorafenib or amentoflavone group.

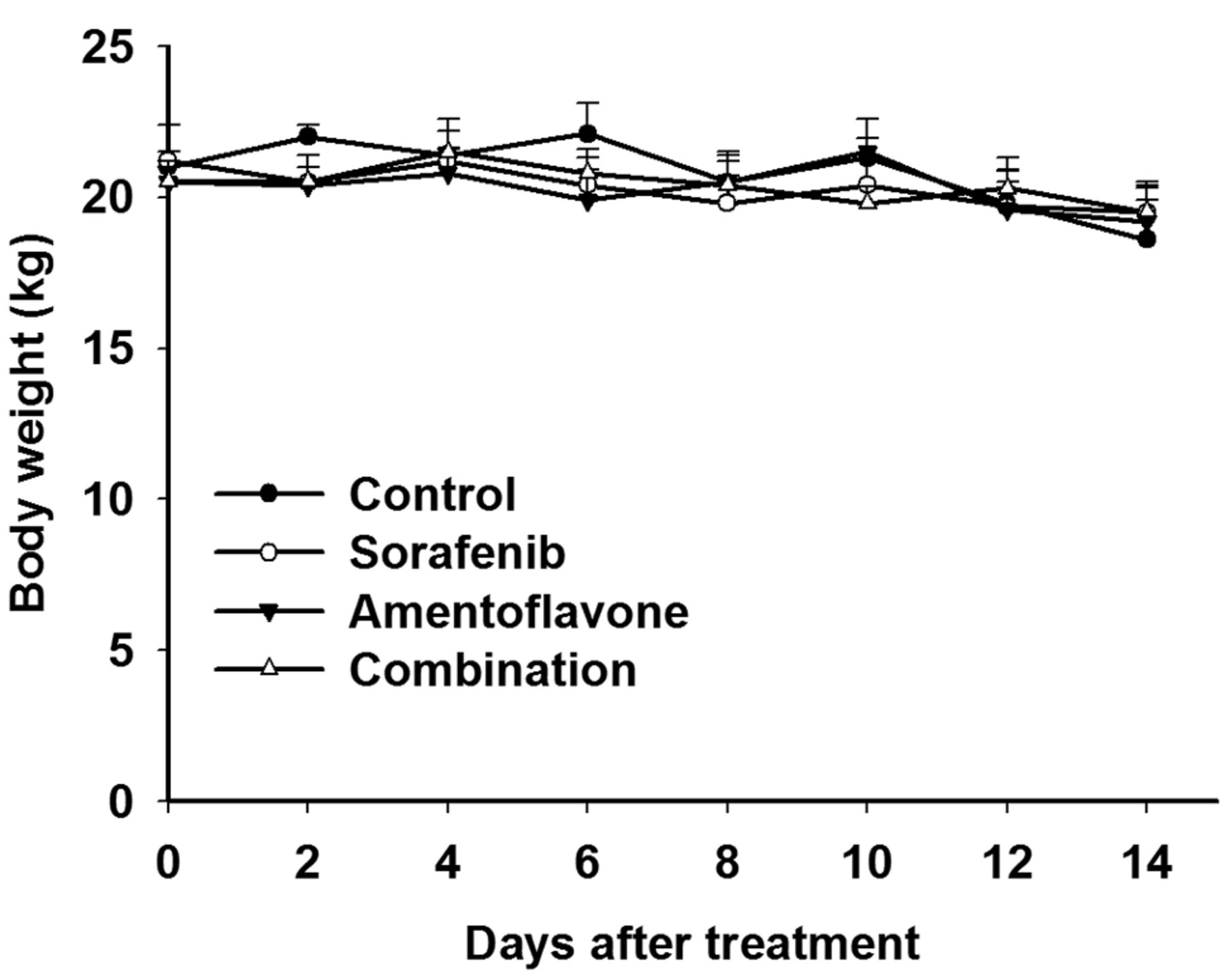
Effect of sorafenib, amentoflavone, or combination on body weight in SK-Hep1 HCC bearing mice. Toxicity of treatment was monitored with body weight.
Toxicity analysis of treatments in SK-Hep1 tumor bearing mice. We used body weight measurement to evaluate the general toxicity of treatments in SK-Hep1 HCC bearing mice. Figure 4 presented body weights that were not significantly changed by sorafenib, amentoflavone, or combination treatment in SK-Hep1 HCC bearing mice.
Discussion
Both rapidly accelerated fibrosarcoma (RAF)/mitogen-activated protein kinase kinase (MEK)/ERK and phosphoinositide 3-kinase (PI3K)/AKT signaling pathways contribute to tumor growth, metastasis, angiogenesis, and anti-apoptotic potential in HCC (19). Sorafenib blocks RAF/MEK/ERK signaling transduction, but does not affect (PI3K)/AKT signaling pathway in HCC (5). Akt phosphorylation modulates acquired resistance to sorafenib and correlates to poor prognosis in HCC (20-21). Inhibition of AKT phosphorylation has been indicated to sensitize HCC to sorafenib. PI-103, a potent inhibitor of AKT (Ser473) phosphorylation, combined with sorafenib significantly suppressed epidermal growth factor (EGF)-stimulated HCC cell proliferation by inhibiting both Raf/MEK/ERK and PI3K/AKT signaling transduction (22). Combination of PI-103 and sorafenib significantly inhibited tumor growth as compared to single-agent treatment in HCC in vivo (23). We demonstrated that combination of amentoflavone and sorafenib significantly reduces tumor growth, protein levels of pERK and AKT (Ser473) in HCC in vivo and ex vivo.
Anti-apoptotic proteins such as X linked inhibitor of apoptosis protein (XIAP), cellular FADD-like IL-1β-converting enzyme (FLICE)-inhibitory protein (C-FLIP), and myeloid cell leukemia 1 are overexpressed in HCC and linked to poor survival of HCC patients. C-FLIP, MCL-1, and XIAP diminish anticancer agents-induced apoptosis through inhibition of extrinsic, intrinsic apoptosis, and caspase-3 activation, respectively (24). Lee et al., performed phase II study of AEG35156, antisense oligonucleotide to XIAP, combined with sorafenib in patients with advanced HCC. The combination of AEG35156 and sorafenib has additional activity in terms of objective response rates (ORR) and moderate benefit on progression-free survival (PFS) as compared to sorafenib alone (25). Hsu et al. presented overexpression of cyclin E1 and myeloid cell leukemia 1 (Mcl 1) reduces sorafenib-triggered apoptosis, whereas inhibition of cyclin E1 and Mcl 1 promotes induction of apoptosis in HCC (26). We showed that amentoflavone significantly enhances sorafenib-inhibited expression of anti-apoptotic proteins MCL-1, C-FLIP, and XIAP in HCC ex vivo.
Many flavonoids augment sorafenib-inhibited cell growth by inducing apoptosis in cancers. Anti-cancer agents activate apoptosis signaling through initiation of extrinsic and intrinsic pathways (16). Wogonin, a flavonoid extracted from Scutellaria baicalensis, combined with sorafenib effectively kills HCC cells through apoptosis potentiation (11). Quercetin, a natural flavonoid derived from fruits and vegetables, enhances sorafenib-induced apoptosis through induction of intrinsic apoptotic pathway in human gliomas. Combination of quercetin and sorafenib significantly increase expression of active-caspase-3 and -9 (13). Fisetin, a dietary flavonoid existed in many fruits and vegetables, has been shown to potentiate sorafenib-induced apoptosis via activation of death receptor-5 mediated caspase-8/caspase-3 and the mitochondria-dependent apoptotic pathway in cervical cancer HeLa cells (27). We demonstrated the combination of amentoflavone and sorafenib significantly trigger expression of activecaspase-9, -8, and -3 as compared to single-agent treatment in HCC ex vivo.
In conclusion, this study revealed amentoflavone as a sorafenib sensitizer to enhance sorafenib-inhibited tumor growth through inhibition of the anti-apoptotic potential and induction of extrinsic and intrinsic apoptotic pathways in HCC in vivo. We suggest that the combination of amentoflavone and sorafenib may be a potential strategy for treatment of patients with HCC.
Acknowledgements
This study was supported by Grants RD2017-021 and RD106039 from the National Yang-Ming University Hospital, I-Lan, Taiwan and Show Chwan Memorial Hospital, Taiwan. The Authors acknowledge the technical services provided by Clinical Medicine Research Laboratory of National Yang-Ming University Hospital, I-Lan, Taiwan and Translational Laboratory, Department of Medical Research, Taipei Medical University Hospital, Taipei, Taiwan.
Footnotes
Conflicts of Interest
The Authors declare no competing financial interests.
- Received December 29, 2017.
- Revision received February 14, 2018.
- Accepted February 15, 2018.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Inactivation of AKT/ERK Signaling and Induction of Apoptosis Are Associated With Amentoflavone Sensitization of Hepatocellular Carcinoma to Lenvatinib
- Reinforcement of Sorafenib Anti-osteosarcoma Effect by Amentoflavone Is Associated With the Induction of Apoptosis and Inactivation of ERK/NF-{kappa}B
- The In Vivo Radiosensitizing Effect of Magnolol on Tumor Growth of Hepatocellular Carcinoma
- Amentoflavone Induces Apoptosis and Reduces Expression of Anti-apoptotic and Metastasis-associated Proteins in Bladder Cancer
- In Vitro and In Vivo Antitumor Effects of Pyrimethamine on Non-small Cell Lung Cancers