


Abstract
The 5-year survival rate for patients with oral cancer remains at 50%, in large part due the high rate of post-treatment recurrence. In this study, we transfected epithelial-specific integrin αvβ6 and Fyn-kinase, a member of the Src-family kinases, into embryonic murine fibroblasts. In oral cancer, expression of αvβ6 is neo-expressed. Using a variety of in vitro assays, including cell migration and multicellular spheroid formation, we determined that these embryonic fibroblasts expressing αvβ6 and Fyn-kinase were able to acquire an epithelial phenotype. This is in direct contrast to human oral SCC, where expression of αvβ6 with Fyn-kinase promotes epithelial to mesenchymal transition. This demonstrates that signaling pathways can be species-specific.
Oral cancer is the sixth most common cancer worldwide. In India, oral cancer accounts for more than 50% of all malignancies. However, in the United States, oral cancer accounts for roughly 5% of new cancer cases and 96% of these are squamous cell carcinoma (SCC) (1). This year, 49,670 new cases of oral cancer will be diagnosed in the United States, and more than 9,700 patients will die from this disease (2).
Oral SCC consists of a morphologically-heterogeneous population of cells. This heterogeneity likely contributes to the varied response to traditional treatment (ionizing radiation and chemotherapy). Our laboratory previously showed that neoexpression of the integrin αvβ6 is one of the more notable changes that occur in human oral epithelial transformation (3).
In this study, when epithelial integrin αvβ6 and an activated Fyn were expressed in murine embryonic fibroblasts they emulated the morphology and motility found in epithelial cells, suggesting mesenchymal to epithelial cell transition MET) had occurred.
Materials and Methods
Cell lines. Cells deficient for Src, Yes, and Fyn (SYF) were generated in 1997 from mouse embryos with null mutations in both alleles of genes for the Src family protein tyrosine kinases, Src, Yes, and Fyn (4). The cells were immortalized using a retroviral vector transducing the SV40 large T-antigen and carry neomycin resistance (4).
Transfection of the β6 integrin into SYF cells established the SYFβ6 cell line. Similarly, the transfection of the constitutively active Fyn into SYF cells resulted in formation of the SYFβ6CAFYN cell line (5). The full-length β6 cDNA was a gift from Dr. Dean Sheppard (University of California at San Francisco). The cDNA for the constitutively active Fyn (CA-Fyn) was a gift from Drs. H. Kawakatsu (University of California at San Francisco) and Dean Sheppard. The cDNA was stably expressed in the murine SYF cell line, using the Retro-X system (Takara Bio, Mountain View, CA, USA) (5). Cells were grown in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (5).
Antibodies. Rabbit monoclonal antibodies to phospho-p44/43 mitogen-activated protein kinase (MAPK), extracellular signal–regulated kinase (ERK) (Thr202/Tyr204) (antibody #9101), MAPK (p44/42 MAPK (ERK1/2) #9102), β-actin (clone13E5), and horseradish peroxidase (HRP)-conjugated anti-rabbit (clone7074) and HRP-anti-mouse (CST #7076) antibodies were purchased from Cell Signaling Technology Inc. (Danvers, MA, USA).
Migration assay. Cell migration was evaluated using a 6.5 mm, 8 μM pore-size Transwell® filter migration system (#3422; Corning Life Sciences, Durham, NC, USA). The undersurface of the filter was coated with 10 μg/ml of fibronectin overnight at 37°C. The lower chamber contained 500 μl of fibroblast-conditioned medium as a chemoattractant. The SYF cells (SYF, SYFβ6, and SYFβ6CAFYN; 2×105) were plated onto the upper surface in serum-free medium. The assay was carried out without serum at 37°C for 12 h in the presence of 5% CO2. The filters were then fixed using 2% paraformaldehyde in phosphate-buffered saline (PBS), and the upper surface wiped clean using cotton swabs. The filter was immersed in crystal violet for 20 minutes (1% paraformaldehyde, 0.5% crystal violet, and 20% methanol in PBS). Filters were rinsed twice with PBS, air-dried, evaluated visually and the number of cells crossing the membrane were determined by counting five random fields/filter (in triplicate).
Generation of multicellular spheroids (MCS). Tissue culture plates (Santa Cruz Biotechnology, Dallas, TX, USA) were coated with a thin layer of 0.6% agarose solution and air-dried overnight at room temperature. Cells were harvested from monolayer cultures using 0.25% trypsin-EDTA and rinsed twice to remove serum. The cells were then placed on agarose-coated plates for 48 h at 37°C in 5% CO2 in serum-free media. The size of the spheroids was traced, analyzed and quantified using NIH Image J (6). Data are expressed as relative value units (rvu).
Western blot. The cells were grown as a monolayer or MCS in serum-free media for 24 h. The cells were then lysed and total protein was extracted using RIPA lysis buffer [50 mM Tris pH 8, 150 mM NaCl, 0.2% sodium dodecyl sulfate (SDS), 1% deoxycholic acid, 1% Triton X-100. complete protease inhibitor tablet (Sigma-Aldrich, Carlsbad, CA, USA). Proteins were resolved using SDS-polyacrylamide gel electrophoresis (PAGE) (7.5%) and transferred to a polyvinylidene fluoride (PVDF) membrane using a semi-dry apparatus (Bio-Rad, Hercules, CA, USA). PVDF membranes were subsequently incubated with monoclonal antibodies listed above overnight at 4°C. After washing, the blots were incubated with HRP-conjugated anti-rabbit antibodies for 1 h. The blots were treated with enhanced chemiluminescence substrate (Thermo Scientific Pierce; Rockford, IL, USA) and exposed to X-ray film (Santa Cruz Biotechnology, Dallas, TX, USA). Western blot images were quantified using NIH Image J (6). β-Actin or tubulin were used as controls to quantify the bands in each image. The images were digitally captured and assigned rvu based on the intensity of the signal. Image J toolbar was used to draw a rectangle around the control lane (tubulin or actin) and sample lanes. Relative area sizes were evaluated from the generated peaks. Each area was expressed as a percentage of the total size of all of the highlighted peaks. Each subsequent percentage was then divided by the percentage of the assigned control to generate expression as rvu.
Results
β6 Integrin and active Fyn promote epithelial cell morphology in undifferentiated murine fibroblasts. The murine SYF, SYFβ6 and SYFβ6CAFYN cells were grown on tissue culture plastic for 120 h under low serum conditions (1%). Both the SYF and the SYFβ6 cells retained their fibroblastic appearance over the course of the assay and never reached confluence (Figure 1). However, the SYFβ6CAFYN cells contracted and took on a compact/epithelial-like morphology and did reach confluence (Figure 1). In the absence of integrin αvβ6 and its ligand Fyn kinase, the murine cells retained their mesenchymal-like phenotype.
Differential formation of MCS by murine SYF cells. We used an MCS assay to evaluate the ability of the murine SYF cells to form three-dimensional structures in vitro. The SYF, SYFβ6 and the SYFβ6CAFYN cells were grown, serum-free, on agarose-coated tissue-culture plates (7). The cultures were monitored every 12 h and at 48 h the experiment was terminated. SYF cells did not readily form cell–cell clusters, but rather formed the smallest MCS (Figure 2). β6 Integrin co-expressed with Fyn kinase resulted in formation of the largest MCS. MCS formation was as follows: SFYβ6CAFYN>>SYFβ6>>SYF cells (Figure 2).
Cell migration as a response to αvβ6 integrin. In order to examine cell migration, the under surface of Transwell filters was coated with fibronectin. Cell migration occurred at different rates across Transwell filters coated with 10 μg/ml of fibronectin, with the SYF cells migrating the least (Figure 3). The forced expression of β6 integrin increased migration by 40% (Figure 3). When both β6 integrin plus a constitutively active Fyn kinase were expressed, cell migration doubled (Figure 3). These results indicate that expression of β6 integrin with Fyn kinase can promote cell migration in embryonic murine fibroblasts.
Differential expression of pMAPK in SYF MCS. MAPKs are often thought of as biological master switches involved in controlling a wide array of cellular responses including proliferation, gene expression, differentiation, cell survival, and apoptosis (8).
The SYF, SYFβ6 and SYFβ6CAFYN cells were grown on tissue culture plates or on agarose-coated plates for 48 h to evaluate MAPK phosphorylation by western blotting.
The total expression of MAPK was approximately equivalent in all cell lines whether grown as a monolayer or as MCS (Figure 4). However, phosphorylation of MAPK was clearly different between the different cell lines (Figure 4). Surprisingly, when grown as MCS, pMAPK was greatest in SYF cells (Figure 4). Phosphorylation of MAPK was barely detectable in SYFβ6 and SYFβ6 cells (Figure 4). These results suggest that activation of MAPK in murine embryonic fibroblasts occurs independently of β6 and FYN.
Discussion
Our previous work focused on oral cancer cell behavior, specifically on the role of the αvβ6 and its signaling potential (5, 9, 10). In this study, we identified factors that promote the epithelial phenotype in Src-kinase-null murine embryonic fibroblasts. Metaplasia in this murine system requires both β6 integrin and Fyn kinase. Our results demonstrate that the integrin β6 and its Fyn kinase promote mesenchymal to epithelial transition (MET) in embryonic murine fibroblasts.
Integrins are heterodimeric glycoproteins consisting of an alpha- and a beta-chain (11). Each subunit contains a long extracellular domain, a transmembrane region and cytoplasmic domain capable of connecting to the actin cytoskeleton and triggering signal transduction events into the cell (12).
Our group previously demonstrated that cell migration was a direct consequence of β6 integrin expression in oral SCC cells (5,10). We previously showed that the engagement of β6 integrin stimulated secretion of matrix metalloproteinases 2, 3 and 9, and localization and stabilization capable of localized degradation of the extracellular matrix (3). Our previous work also showed that the β6 integrin does not mediate initial cell attachment but rather plays a substantial role in secondary adhesion events such as cell migration (3).
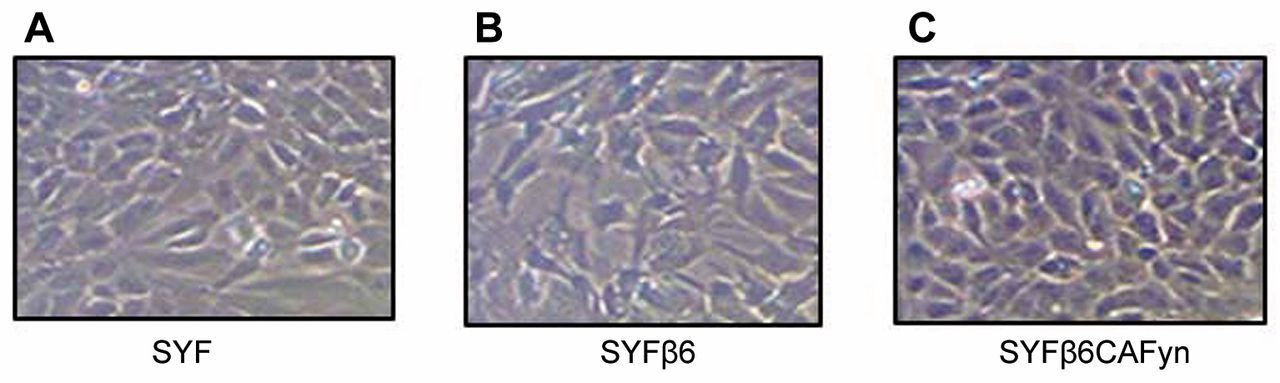
Expression of β6 and Fyn kinase promotes mesenchymal to epithelial transition in SYF cells (with null mutations in both alleles of genes for the Src family protein tyrosine kinases, Src, Yes, and Fyn). SYF cells (A), SYF cells transfected with β6 integrin (SYFβ6) (B) and SYF cells with constitutively active Fyn (SYFβ6CAFyn) (C) were grown on serum-free tissue culture plates for 48 h. SYF and SYFβ6 cell lines retained a-fibroblast shape, while SYFβ6CAFyn cells contracted, becoming more epithelial in morphology. Magnification used: 200×.

Multicellular spheroid (MCS) formation is regulated by β6 and Fyn kinase. SYF cells (with null mutations in both alleles of genes for the Src family protein tyrosine kinases, Src, Yes, and Fyn) (A), SYF cells transfected with β6 integrin (SYFβ6) (B) and SYF cells with constitutively active Fyn (SYFβ6CAFyn) (C) were grown on agarose-coated plates for 120 h. rvu: Relative value units. Magnification used: 200×.
The MAPK/ERK pathway takes extracellular information and communicates this information to the nucleus of the cell. p42 and p44 MAPKs are activated through integrin clustering (13). In this murine SYF system, it appears that pMAPK suppresses MET whether the cells are grown as a monolayer or MCS. In human oral cancer, expression of β6 is associated with tumor aggressiveness and invasion. In the murine system, we found the exact opposite occurs (14).
In the current study, pMAPK was expressed the greatest by the SYF cells. In this system, the expression of β6 with CAFYN suppressed MAPK phosphorylation, which is one of the master switches in cell signaling. This indicates that not all major signaling processes occur in all species.
In this study, murine embryonic fibroblasts were genetically modified to not express any member of the Src family (Src, Yes and Fyn). They retained their normal fibroblast-like morphology except in cells in which integrin β6 and a constitutively active Fyn were co-expressed. This co-transfection resulted in the murine cells taking on an epithelial cell morphology, increasing cell migration and enhancing MCS formation (15). This indicates that cell morphology, migration, and MCS formation are directed by the interaction between β6 and Fyn, but pMAPK is independent of this interaction. In the murine embryonic fibroblast system, the expression of αvβ6 with Fyn-kinase promotes mesenchymal to epithelial transition. This is in direct contrast to human oral SCC, where expression of αvβ6 with Fyn-kinase promotes epithelial to mesenchymal transition. This demonstrates that signaling pathways can be species-specific.

Invasion is modulated by β6 and Fyn kinase. SYF cells (with null mutations in both alleles of genes for the Src family protein tyrosine kinases, Src, Yes, and Fyn), SYF cells transfected with β6 integrin (SYFβ6) and SYF cells with constitutively active Fyn (SYFβ6CAFyn) were plated onto Transwell filters in which the bottom surface was coated with 3 μg/ml of fibronectin. The cells were allowed to migrate onto fibronectin substrate for 24 h, after which the number of cells traversing the filter was determined.
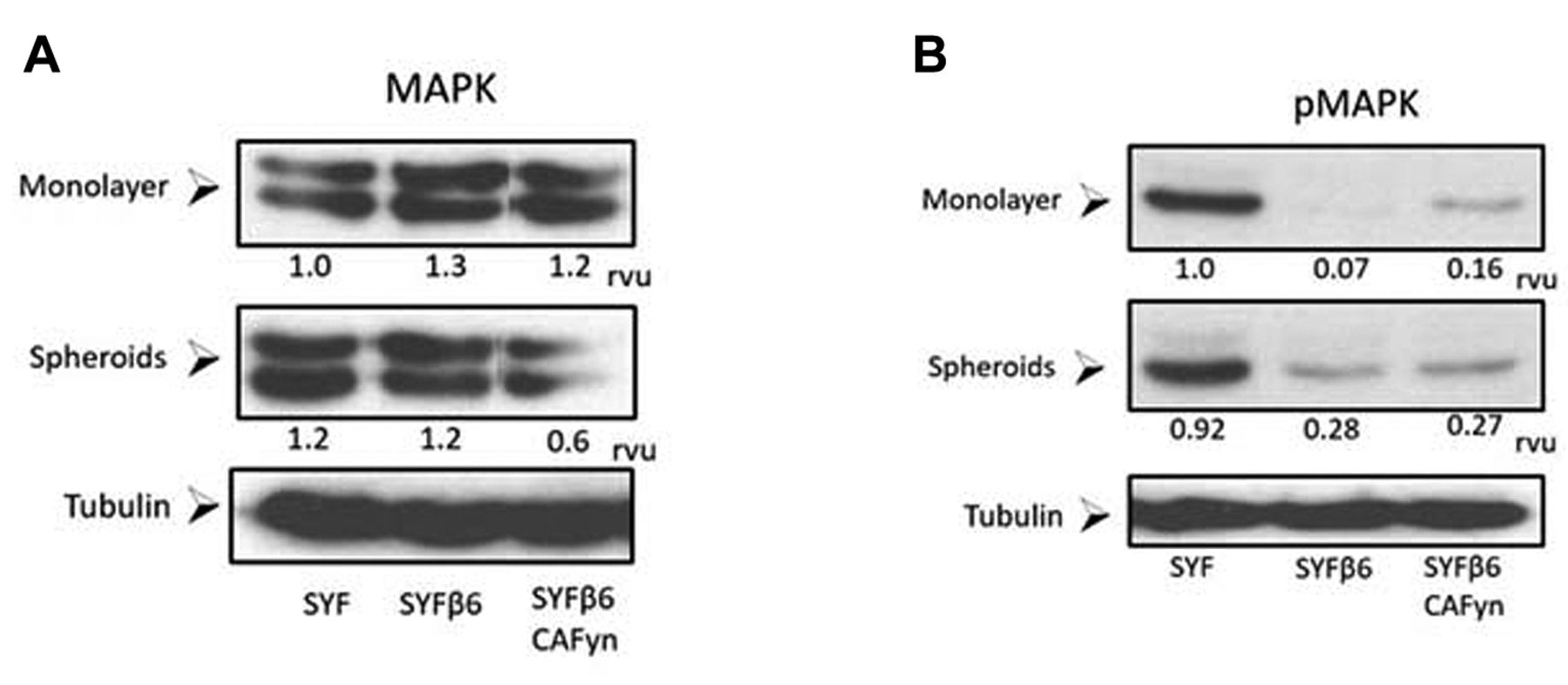
Activation of mitogen-activated protein kinase (MAPK) occurs independently of β6 or activation of FYN. SYF cells (with null mutations in both alleles of genes for the Src family protein tyrosine kinases, Src, Yes, and Fyn), SYF cells transfected with β6 integrin (SYFβ6) and SYF cells with constitutively active Fyn (SYFβ6CAFyn) were grown as monolayers or multicellular spheroid for 24 h and evaluated by western blotting for activation of MAPK (pMAPK). The blots were quantified and assigned relative value units (rvu) using NIH Image J.
- Received October 11, 2017.
- Revision received November 13, 2017.
- Accepted November 16, 2017.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.