


Abstract
Background/Aim: Maspin is known to be a tumor suppressor protein. Its nuclear localization and endogenous inhibition of histone deacetylase 1 (HDAC1) are considered crucial for its tumor suppressor activity. However, it remains unclear whether subcellular localization of maspin correlates with HDAC1 expression level in human breast cancer. Patients and Methods: Immunohistochemical analyses were performed on 164 resected specimens of invasive breast carcinoma using antibodies for maspin and HDAC1. Subcellular localization of maspin protein and HDAC1 mRNA expression level in two human breast cancer cell lines (MCF7, MDA-MB-231) and mammary epithelial cell line (MCF10) were analyzed by immunofluorescence and quantitative polymerase chain reaction, respectively. Results: The frequency of cytoplasmic-only, pancellular (combined nuclear and cytoplasm) and no staining of maspin were 31%, 14.0% and 55%, respectively. The cytoplasmic-only subgroup showed significantly higher histological grade (p=0.004), negative progesterone receptor status (p=0.003) and shorter disease-free survival compared to the pancellular subgroup (p=0.043). High HDAC1 expression was observed in 60% of cases and was significantly correlated with cytoplasmic-only staining compared to pancellular (p<0.001) or no staining (p=0.004). Immunofluorescence analysis revealed that maspin protein was localized mainly in the cytoplasm in MCF7 and MDA-MB-231 cells, while in both the nucleus and cytoplasm in MCF10A cells. HDAC1 mRNA levels were significantly up-regulated in MCF7 and MDA-MB-231 cells compared to MCF10A cells (p<0.001). Conclusion: High HDAC1 expression may contribute to the aggressiveness of human breast cancer with cytoplasmic-only expression of maspin.
Maspin, a unique member of the serine protease inhibitor (serpin) superfamily, was identified originally as a tumor suppressor that is expressed in normal breast epithelial cells but is down-regulated in breast cancer cells (1). Maspin has been shown to inhibit both tumor growth and metastasis in animal models and cancer cell lines, and it has pro-apoptotic, anti-metastatic and anti-angiogenic properties, exerting an inhibitory effect on cancer cell motility, invasiveness and metastatic ability (2). Although the exact biochemical pathways leading to these biological endpoints remain elusive, several studies have characterized the association between maspin expression and clinicopathological factors in breast cancer patients (3-6). We have also reported that cytoplasmic-only expression of maspin could be a poor prognostic indicator in breast cancer patients (7-9). On the other hand, it has been reported that the nuclear localization of maspin in cancer cells was necessary for its inhibition of tumor progression and that the endogenous inhibition of histone deacetylase 1 (HDAC1) could be one of the tumor suppressive mechanisms of maspin (10-12). Taken together, we hypothesized that cytoplasmic-only expression of maspin might be positively correlated with high expression of HDAC1 in human breast cancer. To our knowledge, no study has reported the association between subcellular localization of maspin protein and HDAC1 protein expression in human breast cancer tissue samples. The aim of the present study was to clarify whether subcellular localization of maspin correlates with HDAC1 expression level in human breast cancer in order to gain insight of the tumor suppressive mechanisms of maspin.
Patients and Methods
Patients and tumors specimens. From January 2008 to December 2011 at Tottori University Hospital, Japan, 227 consecutive female patients underwent a surgical resection of breast cancer diagnosed as invasive carcinoma of no special type according to the World Health Organization classification of breast cancer (13). Sixty-three cases were excluded due to a history of neoadjuvant treatment (38 cases), microinvasive carcinoma (9 cases), distant metastasis (5 cases), unavailable tumor specimen (5 cases), bilateral breast cancer (4 cases) and short follow-up period (2 cases). Thus, 164 patients were included in this analysis. The patients' clinicopathological data were obtained from their hospital medical records. The histopathological parameters including lymph node metastasis, estrogen receptor (ER), progesterone receptor (PR) and human epidermal growth factor receptor 2 (HER2) statuses were retrieved from the patients' pathology reports. Histological grades were determined according to the criteria of Elston and Ellis (14), and the grading was independently evaluated by M.W. and Y.U. Tumor stage was determined based on the seventh tumor–node–metastasis (TNM) classification of breast cancer (15). The study was approved by the ethics committee of the Faculty of Medicine, Tottori University (approval number: 1608A083; 15 August 2016).
Immunohistochemistry. All specimens were fixed in 10% neutrally buffered formalin and embedded in paraffin. The sections (4 μm-thick) were deparaffinized and endogenous peroxidase activity was blocked. The specimens were then pretreated in citrate buffer (0.01 M, pH 6.0) in a microwave oven (IMB-T17-5/6; Irisohyama, Sendai, Japan) for 15 min at 700W for maspin, or a water bath at 95°C for 10 min for HDAC1. After the pretreated specimens had cooled to room temperature, non-specific binding was blocked by blocking buffer (Block Ace; Megmilk Snow Brand, Sapporo, Japan). Next, the sections were incubated at 4°C overnight with a monoclonal antibody to maspin (clone EAW24, diluted 1:200; Leica Biosystems, Newcastle, UK) or at room temperature for 30 min with polyclonal antibody to HDAC1 (clone ab19845, diluted 1:3600; Abcam, Cambridge, MA, USA). The sections for maspin were incubated with biotinylated anti-mouse IgG antibody (BA-2000, diluted 1:150; Vector Laboratories, Burlingame, CA) for 20 min, followed by streptavidin biotinylated horseradish peroxidase (HRP) conjugate (diluted 1:150; Invitrogen, Camarillo, CA, USA) for 20 min. The sections for HDAC1 were incubated with the EnVision+ System HRP (Dako, Glostrup, Denmark). Finally, all of the sections were incubated with diaminobenzidine (DAB) solution (liquid DAB+ substrate, imidazole-HCI buffer, pH 7.5, containing hydrogen peroxide and an antimicrobial agent; Dako) for 4 min, and counterstained with hematoxylin.
Evaluation of immunohistochemical findings. The cells were considered positive for maspin expression when strong cytoplasmic-only staining was identified. Strong staining was defined as the staining intensity equal to that of myoepithelial cells in normal breast tissue, and it served as an internal positive control if included. Tumors with >10% positive cells were considered maspin-positive as described previously (8). The subcellular localization of maspin was classified into four categories; cytoplasmic-only, pancellular (combined nuclear and cytoplasmic), nuclear-only, and no staining. The evaluation of HDAC1 expression status was performed according to the methods described by Seo et al. (16). Briefly, the intensity (0, negative; 1, weak; 2, moderate; 3, strong) and the proportion of positive cells (0, negative; 1, <10%; 2, 10-33%; 3, 33-66%; 4, ≥66%) scores were calculated through multiplication and cases were divided into two groups according to their scores as follows: low expression (scores, 0-6) and high expression (scores, 8-12). All slides were evaluated by M.W. and Y.U. independently who were blinded to the patients' clinicopathological data.
Cell culture. The human mammary epithelial cell line MCF10A and the human breast cancer cell line MDA-MB-231 were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). MCF10A cells were maintained in mammary epithelial cell growth medium (MEGM Bullet Kit; Lonza, Walkersville, MD, USA) supplemented with 100 ng/mL cholera toxin at 37°C with 5% CO2. MDA-MB-231 cells were maintained in Leibovitz's L-15 Medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS; Biological Industries Ltd. Kibbutz Beit Haemek, Israel) and L-glutamine at 37°C with 0% CO2. The human breast cancer cell line MCF7 was purchased from the Riken BioResource Center Cell Bank (Riken BRC Cell Bank, Ibaraki, Japan) and cultured in Dulbecco's Modified Eagle Medium (DMEM; Nissui Pharmaceutical, Tokyo, Japan) supplemented with 10% FBS, glucose, and L-glutamine at 37°C with 5% CO2.
Immunofluorescence analysis. MCF10A (4.0×104 cells/cm2), MCF7 (2.0×104 cells/cm2), and MDA-MB-231 (4.0×104 cells/cm2) cells were separately seeded in 8-well chamber slides (Nalgen Nunc International, Rochester, NY, USA) and incubated for 24 h. After incubation, the cells were fixed with 4% paraformaldehyde at room temperature for 15 min and permeabilized with ice-cold 100% methanol at −20°C for 10 min. These samples were blocked in 10% goat serum/ phosphate-buffered saline (PBS) at room temperature for 60 min and subsequently incubated with the anti-Maspin primary antibody (G167-70, diluted 1:10; BD Pharmingen, Franklin Lakes, NJ, USA) at 4°C overnight, followed by incubation with Alexa Fluor conjugated anti-mouse secondary antibody (Molecular Probes, Leiden, Netherlands). Nuclear counterstaining was performed using DAPI (Molecular Probes, Leiden, Netherlands). Immunofluorescence images were detected using an FV1000 confocal microscope (Olympus, Tokyo, Japan).
RNA extraction and quantitative polymerase chain reaction. Total RNA from MCF10A, MCF7, and MDA-MB-231 cells was extracted using TRIzol Reagent (Thermo Fisher Scientific, Waltham, MA, USA) and reverse-transcribed into complementary DNA (cDNA) using the High-Capacity RNA-to-cDNA Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to manufacturer's protocol. We measured the gene expression levels by performing TaqMan Gene Expression Assays (Thermo Fisher Scientific, Waltham, MA, USA) with the following gene-specific primers: GAPDH, Human GAPDH Endogenous Control (Hs99999905_m1, Thermo Fisher Scientific, Waltham, MA, USA) and HDAC1, TaqMan Gene Expression Assays (Hs00606262_g1, Thermo Fisher Scientific, Waltham, MA, USA).
Statistical analysis. All statistical analyses were performed using the SPSS version 23 software (IBM SPSS Statistics; IBM Corporation, Armonk, NY, USA). The associations between maspin expression status and clinicopathological factors, and the correlation between the subcellular localization of maspin and HDAC1 expression status were evaluated by the Chi-square test. We compared the relative HDAC1 mRNA expression by using a one-way analysis of variance (ANOVA) and the Tukey honest significant difference (HSD) test. For the survival analyses, cancer relapse (local recurrence or distant recurrence) was used to calculate disease-free survival (DFS). DFS was defined as the period from the date of primary surgery to the date of clinical or pathological cancer relapse. In the calculation of DFS, patients were censored at the time of their last cancer-free follow-up or at the time of death due to a reason unrelated to cancer. Survival curves were plotted by the Kaplan-Meier method and differences in DFS were tested using the log-rank test. All tests were two-sided, and differences were considered significant at p-values<0.05.
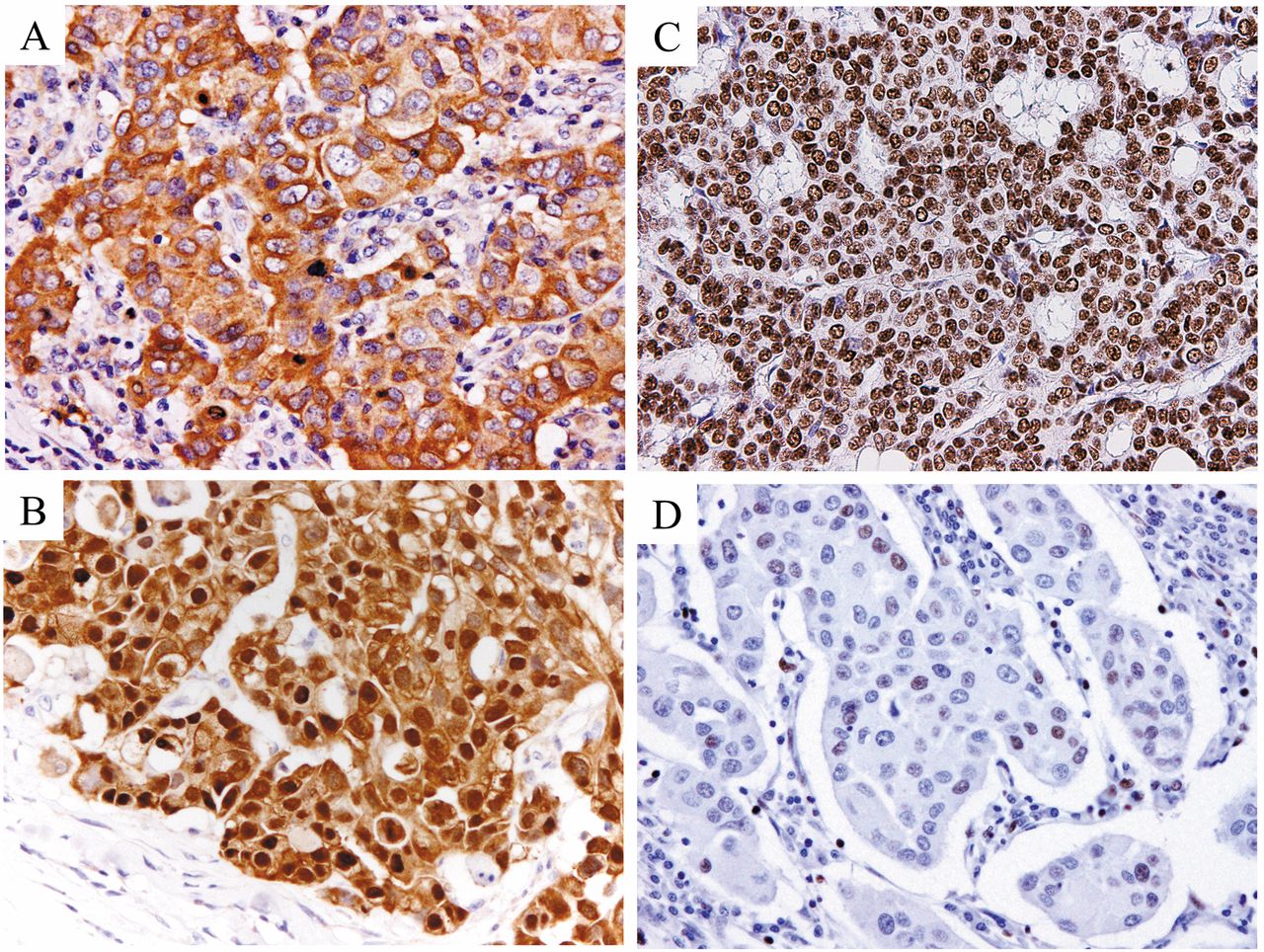
Immunohistchemical staining patterns of maspin in invasive carcinoma of the breast. A: Cytoplasmic-only expression of maspin. B: Pancellular (combined nuclear and cytoplasmic) expression of maspin. Immunohistchemical staining of HDAC1 in invasive carcinoma of the breast. C: High expression of HDAC1 (intensity score: 3, proportion score: 4). D: Low expression of HDAC1 (intensity score: 2, proportion score: 2).
Results
Clinicopathological characteristics. The median age of the 164 patients at the time of operation was 60 years (range=32-90 years), and the median follow-up time was 68.5 months (range=9-104 months). The positive rates of ER, PgR and HER2 were 82.3%, 64.6% and 16.5%, respectively. Twenty-one patients (12.8%) experienced recurrence, and three patients (1.8%) deceased of breast cancer progression.
Subcellular localization of maspin. Representative immunohistochemical staining patterns of maspin expression are shown in Figure 1A and B. Subcellular localization of maspin expression was as follows: cytoplasmic-only staining (50 cases, 30.5%), pancellular (combined nuclear and cytoplasm) staining (23 cases, 14.0%), and no staining (91 cases, 55.5%). Nuclear-only staining was not observed.
Relationship between clinicopathological features and subcellular localization of maspin. The associations between the clinicopathological features and subcellular localization of maspin expression are summarized in Table I. Cytoplasmic-only expression of maspin significantly correlated with higher histological grade (p=0.004) and negative PR status (p=0.003). Pancellular expression of maspin significantly correlated with lower pathological tumor classification (p=0.005).
Association between subcellular localization of maspin and clinicopathological characteristics.
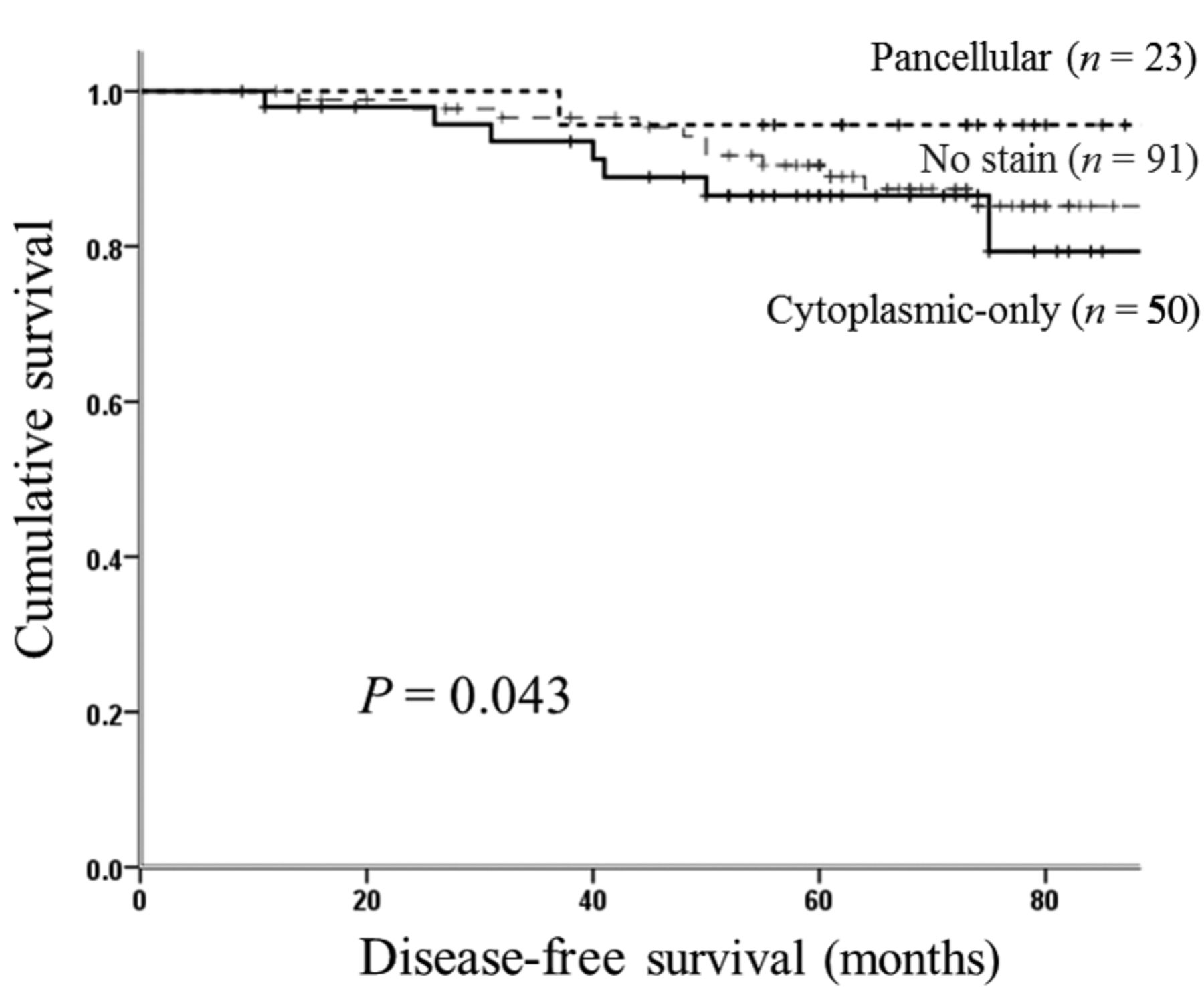
Survival analysis according to maspin expression status. For the survival analysis, we classified the subcellular localization of maspin expression into three groups. DFS curve for the patients is shown in Figure 2. The 5-year DFS rates of the cytoplasmic-only staining group, pancellular staining group and no staining group were 86.5% (95% confidence interval [CI]=72.4-93.7%), 95.7% (95% CI=72.9-99.4%) and 90.5% (95% CI=81.8-95.1%), respectively. According to the log-rank test, the cytoplasmic-only staining group showed significantly shorter DFS compared to the pancellular staining group (p=0.043).
Immunohistochemical findings of HDAC1. HDAC1 was observed in the nucleus and was expressed in both tumor cells and normal epithelium (Figure 1C and D). Regarding the staining intensity of HDAC1, the proportion of strong, moderate and weak staining was 35.4%, 43.3% and 18.9%, respectively. The proportion of HDAC1-positive cells was as follows: 58.5% (96 cases) at ≥66%, 21.3% (35 cases) at 33-66%, 12.8% (21 cases) at 10-33%, and 4.9% (8 cases) at <10%. Four (2.4%) cases showed no expression of HDAC1. Thus, high expression of HDAC1 was observed in 98 cases (59.8%) and it was correlated only with lower histological grade (p= 0.027). There was no significant difference in DFS between the patients showing high and low expression of HDAC1.

Kaplan-Meier survival curve for disease-free survival of the 164 patients according to subcellular localization of maspin expression (cytoplasmic-only, pancellular, and no staining groups).

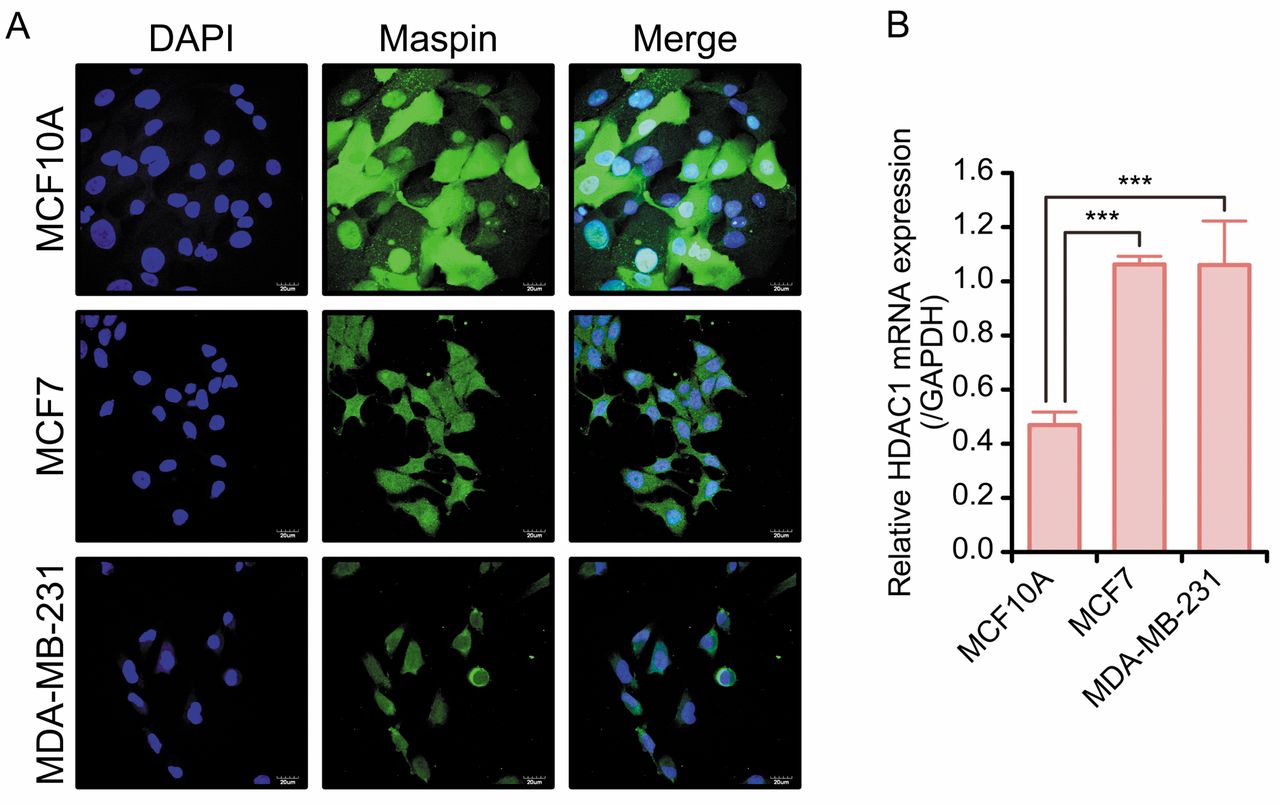
Association between HDAC1 mRNA expression and subcellular localization of maspin in normal breast epithelial cell line and breast cancer cell lines. A: Subcellular localization of maspin in MCF10A, MCF7, and MDA-MB-231 cells was analyzed by immunofluorescence studies with maspin-specific antibody. Maspin (Alexa 488, green) and nuclei (DAPI, blue) signals were detected by laser confocal microscopy. B: Expression of HDAC1 mRNA in MCF10A, MCF7 and MDA-MB-231 cells was examined by qPCR. HDAC1 mRNA expression level was normalized to the expression of GAPDH. Data are shown as mean±standard deviation (SD) (n=3). ***p<0.001 by one-way ANOVA and Tukey's HSD test.
Association between subcellular localization of maspin and HDAC1 expression. The correlation between subcellular localization of maspin and HDAC1 expression is summarized in Table II. Cytoplasmic-only subgroup was significantly positively correlated with high expression of HDAC1 compared to the pancellular subgroup (p<0.001) and compared to the no staining subgroups (p=0.004). The pancellular subgroup showed a significant positive correlation with the low expression of HDAC1 compared to the no staining subgroup (p=0.028).
Association between subcellular localization of maspin and HDAC1 mRNA expression in breast cancer cells. Immunofluorescence analysis showed that maspin protein was localized mainly in the cytoplasm in MCF7 and MDA-MB-231 cells, whereas it was localized in both the nucleus and cytoplasm in MCF10A cells (Figure 3A). The protein levels of maspin in MCF7 and MDA-MB-231 cells were lower than those in MCF10A cells (data not shown). The HDAC1 mRNA levels were significantly up-regulated in MCF7 and MDA-MB-231 cells compared to those in MCF10A cells (p<0.001) (Figure 3B).
Correlation between subcellular localization of maspin and HDAC1 expression.
Discussion
We previously reported that cytoplasmic-only expression of maspin was an independent poor prognostic indicator in breast cancer patients (6) and that its expression was down-regulated through the progression from ductal carcinoma in situ to invasive ductal carcinoma using immunohistochemical analysis (7). In the largest series of breast cancer patients by Mohsin et al., it has been reported that cytoplasmic staining of maspin was related to ER and PgR negativity, high S-phase fraction and aneuploidy, and that the nuclear staining of maspin was associated with good prognostic factors (4). The same study also described that cytoplasmic staining showed some nuclear staining in all but nine of the cases, and therefore a very few cases of cytoplasmic-only staining were observed (4). This discrepancy of subcellular localization of maspin may be attributable to the difference of the antibody used. In the present study, cytoplasmic-only expression of maspin was observed in 30.5% of the 164 samples, which is a frequency similar to our previous investigation's finding (27.4%) (6). Our present study also confirmed that cytoplasmic-only expression of maspin was significantly correlated with shorter DFS despite the relatively short median follow-up period, higher histological grade and PgR negativity, as we observed in the previous investigation (6). In addition, we revealed for the first time that pancellular expression of maspin was not correlated with DFS or a marker for an aggressive phenotype (i.e., with a higher histological grade, larger tumor size, ER/PgR negativity, and lymph node metastasis). Goulet et al. reported that the nuclear localization of maspin was required for its tumor and metastasis suppressor function and that tumor cells expressing nucleus-excluded, cytoplasmic-only maspin were more metastatic than the controls in an in vivo model system (10). They speculated that cytoplasmic-only – not pancellular – expression of maspin might be correlated with an aggressive phenotype, which supports our present and previous findings using clinical samples. As the tumor suppressor activity of maspin requires its nuclear localization, it is important to elucidate how maspin gets recruited to chromatin and regulates gene expression. HDAC1, a member of the class I HDACs, is the most extensively characterized protein of this family. An increased deacetylation of histones leads to an increase of cell proliferation and migration, angiogenesis and invasion by reducing the transcription of tumor suppressor genes (17). Maspin is the only endogenous polypeptide HDAC1 inhibitor identified thus far and the endogenous inhibition of HDAC1 has been considered one of the tumor-suppressive mechanisms of maspin (11). We therefore speculated that cases with cytoplasmic-only expression of maspin would frequently show high expression of HDAC1.
To our knowledge, the present findings demonstrated for the first time that cytoplasmic-only expression of maspin positively correlated with high HDAC1 expression, and that pancellular expression of maspin is positively associated with low HDAC1 expression compared to no expression in clinical samples. Our results also demonstrated that two human breast cancer cell lines with cytoplasmic-predominant expression of maspin showed higher HDAC1 expression compared to human mammary epithelial cell lines with pancellular expression of maspin. This observation in the in vitro experiment supports our data from the clinical samples, suggesting that subcellular localization of maspin could play an important role in the regulation of HDAC1. However, the molecular interaction between maspin and HDAC1 may not be mutually exclusive. In fact, HDAC1 target genes include p27KIP1, p21WAF/CIP1 and Bax, whereas maspin interact with glutathione S-transferase (GST), interferon-responsive factor 6 (IRF6) and pro-urokinase type plasminogen activator (pro-uPA) in an HDAC1-independent manner (2). Further investigation is necessary to clarify the precise mechanism and to determine the significance of cytoplasmic-only expression of maspin.
In conclusion, our present results demonstrated that cytoplasmic-only (not pancelluar expression) expression of maspin was an unfavorable prognostic indicator and was positively correlated with high HDAC1 expression in human breast cancer tissue samples as well as human breast cancer cell lines. These results support the hypothesis that the nuclear localization of maspin in cancer cells is necessary for its inhibition of tumor progression and that an endogenous inhibition of HDAC1 could be one of the tumor-suppressive mechanisms of maspin.
Acknowledgements
The Authors are grateful to Kazuo Yashima and Kazuko Fukushima for their excellent technical assistance with the processing of the pathological specimens. This work was supported by JPSS KAKENHI GRANT Number JP16K10458.
Footnotes
Conflicts of Interest
The Authors confirm that there are no conflicts of interest.
- Received June 28, 2017.
- Revision received July 5, 2017.
- Accepted July 6, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Cytoplasmic-only Expression of Maspin Predicts Poor Prognosis in Patients With Oral Squamous Cell Carcinoma
- Cytoplasmic-only Expression of Maspin Predicts Unfavorable Prognosis in Patients With Pancreatic Ductal Adenocarcinoma
- Clinical Significance of Subcellular Localization of Maspin in Patients with Pathological Stage IA Lung Adenocarcinoma