


Abstract
Background/Aim: Vascular endothelial growth inhibitor (VEGI) is a multipotential cytokine that plays a role in regulating immunity, anti-angiogenesis, and inhibiting tumor growth. However, the proteins that interact with it are still unknown. In the present study, we examined the proteins that interact with VEGI174 and their expression in renal cell carcinoma (RCC). Materials and Methods: The proteins that interact with VEGI174 were identified using western blot, pull-down assay, and mass spectrometry. The expressions of VEGI174 and the interacting proteins were examined in RCC and were compared to normal renal tissues using immunohistochemical staining and RNA-seq respectively. Results: The results of the mass spectrometric analysis showed that ACLY, ENO1, ZIK1, AKR1C3, and MYC may interact with VEGI174. When compared to the TCGA database, the expression level of VEGI174 in RCC was lower than that in normal kidney using RNAseq (p<0.001). The expression levels of ACLY, ENO1, ZIK1, AKR1C3 and MYC in RCC were higher than those in normal kidney (p<0.05, all of above factors). Moreover, immunochemical staining results also showed that the expression levels of AKR1C3 in RCC were significantly higher those that in normal kidney (p<0.001) and was also positively correlated with higher RCC stage and grade. Conclusion: Taken together, our findings showed that VEGI174 may interact with ACLY, ENO1, ZIK1, AKR1C3, and MYC. The expression of ACLY, ENO1, AKR1C3 and MYC is increased in RCC. AKR1C3 was a new factor that may correlate with the progression of RCC. The results indicated that VEGI174 has more functions than we currently know in the development and progression of RCC.
- Vascular endothelial growth inhibitor (VEGI)
- renal cell carcinoma
- pull-down assay
- mass spectrometric
- AKR1C3
Vascular endothelial growth inhibitor, VEGI (also known as tumor necrosis factor superfamily member 15 (TNFSF15) and TNF ligand related molecule 1 (TL1)), is an anti-angiogenic cytokine that belongs to the TNFSF family of proteins (1-3). Three isoforms of VEGI (VEGI 174, VEGI192, VEGI 251) have been reported, all of them share a common region of 453bp that encodes a domain of 151 amino acids at the C-terminus but they differ in their N-terminal regions (4-6). The biological activity of VEGI mainly depends on the solubilised extracellular domain of the three isoforms. VEGI is a multipotential cytokine that plays a role in inflammation, septic shock, fever, and growth modulation through inducing apoptosis, regulating immunity and anti-angiogenesis. Thus, VEGI may play a role as a target for anti-tumor growth via its inhibitory role on anti-angiogenesis and regulating immunity (7). A great amount of evidence has also shown that overexpression of VEGI could inhibit tumor neovascularisation and progression in cellular and animal models (2, 8).
First, VEGI is able to inhibit the growth of various human tumor cell lines including human histiocytic lymphoma (U-937), human breast carcinoma (MCF-7), human epithelial carcinoma (HeLa) and human myeloid lymphoma ML-1a (9). Second, VEGI is shown to inhibit tumor growth in vivo. It also suppresses the growth of colon carcinoma cells (murine colon cancer cells, MC-38) both in vitro and in vivo (2, 3, 10). Systemic administration of VEGI 192 remarkably inhibited tumor growth and increased survival time of treated animals in a Lewis lung carcinoma (LLC) murine tumor model (11). In 2006, Parr et al. reported that patients with breast tumors expressing reduced levels of VEGI had a higher local recurrence, shorter survival time and an overall poorer prognosis than patients expressing higher levels of VEGI (7). In 2013, Jia et al. reported that the expression level of VEGI was significantly lower in pituitary tumors which invaded the sellafloor, and with suprasellar extension than that of non-invasive tumors (12). They suggested that VEGI was an independent indicator for invasion in human pituitary adenomas. Our previous studies demonstrated that VEGI expression could significantly reduce motility and adhesion of renal, prostate and bladder cancer cells (13, 14). VEGI expression was decreased in renal cell carcinoma (RCC), particularly in high-grade tumors. Furthermore, full length VEGI174 could suppress the growth of RCC in vivo (15-17). In addition, VEGI is also a T-cell co-stimulator and can directly stimulate dendritic cell maturation (8, 18). However, it is still unclear whether activation of tumor specific or non-specific B or T lymphocytes may operate in soluble VEGI-mediated tumor suppression.
It is well known that tumor angiogenesis and immunity are closely related with RCC progression. Our studies showed that VEGI174 might be a putative tumor suppressor and a potential therapeutic target for RCC. However, it is unclear whether VEGI174 acts solely or in collaboration with other proteins as part of a protein complex. In this study, the proteins that interact with VEGI174 were identified by immunoprecipitation and mass spectrometry. The expression of VEGI174 and associated proteins was also examined in RCC tissues using the RNA-seq method.
Materials and Methods
Cell lines. All cell lines used in this study were purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA). Human emborynic kidney 293 T cell line (HEK293T) and Chinese hamster ovarian line (CHO) were used in this study. Cells were routinely cultured with Dubecco's modified Eagle's medium (DMEM) supplemented with 10% fetal calf serum, penicillin and streptomycin (Gibco BRC, Paisley, UK).
Antibodies. Antibodies were purchased from the following sources: The monoclonal anti-Flag M2 Tag was purchased from Sigma-Aldrich Co. LLC. (F1804, Sigma-Aldrich Co. LLC., St. Louis, USA). The monoclonal Rabbit anti-human-ACLY (ATP citrate lyase) and anti-human-ENO1 (Enolase 1) antibodies were purchased from Abcam Inc. (EP704Y and EPR10863, Abcam Inc., Cambridge Science Park, UK). The monoclonal mouse anti-human-VEGI174 antibody was purchased from LSBio Inc. (LS-C76815, LSBio Inc., Seattle, WA, USA). Monoclonal mouse anti-human-AKR1C3 (Aldo-ketoreductase family 1 member C3) antibody was kindly supplied by Professor Qing from the Centre for Cellular & Structural biology, School of Pharmaceutical Sciences, Sun Yat-Sen University, Guangzhou, P.R. China.
Human RCC specimens. A total of 16 pairs of renal cell carcinoma and normal renal tissue samples were snap-frozen in liquid nitrogen immediately after radical nephrectomy. The pathologist verified normal and cancer specimens. All protocols were reviewed and approved by local ethics committee and all patients were given written informed consent. Nine patients were in clinical stage T1 and 7 patients were in T3. In addition, 1 patient was in Fuhrman Grade l, 8 patients were in Fuhrman Grade 2, 6 patients were in Fuhrman Grade 3, and 1 patient was in Fuhrman Grade 4.
RNA isolation and reverse transcription PCR. RNA was isolated using Total RNA Isolation reagent (ABgene, Epsom, UK). First strand cDNA was synthesized from 0.5 μg RNA using a reverse transcription kit (Sigma, Poole, Dorset, UK). The quality of cDNA was verified through amplification and detection of the GAPDH housekeeping gene. Forward and reverse primers of VEGI (Table I) were designed based on human VEGI sequence (GeneBank accession no. BD131562). PCR was performed in a GeneAmp PCR system 2400 thermocycler (Perkin-Elmer, Norwalk, CT, USA). Conditions for PCR were 40 sec at 94°C, 60 sec at 55°C, 60 sec at 72°C (35 cycles). PCR products were separated on a 1.4% agarose gel.
Establishment of Stable Transfectants. CHO and HEK293T cells were transfected with 3 μg of full-length human VEGI174 coding sequence (Table I) cloned into pcDNA3.1+ expression vector (Invitrogen Inc., Paisley, UK) by Lipofectamine 2000. CHO-stable transfectants of full-length human VEGI174 coding sequence were selected for 2 weeks by 2 mg/ml G418 (Invitrogen Inc., Paisley, UK) before being subjected to cloning by limiting dilution.
Western-blotting analysis. CHO or HEK293T cells (1×106) at 48 h after transient transfection with 3 μg of full length VEGI174 using Lipofectamine 2000 (Invitrogen Inc., Paisley, UK), or the same number of the CHO stable cloned VEGI174 were lysed in 200 μl of lysis buffer (50 mm NaCl, 20 mm Tris, pH 7.6, 1% Nonidet P-40, 1× protease inhibitor mixture) for 1 h. The lysates were cleared by centrifugation at 16,000 × g at 4°C for 10 min. Crude protein concentration was measured by DC Protein Assay kit (Bio-Rad) and ELx800 spectrophotometer (Bio-Tek™). Two hundred μg of total protein of each sample was transferred to a new microcentrifuge tube, and immunoprecipitation was performed with 2 μg of the indicated antibodies and 50 μl of protein-G slurry for overnight incubation. Ten μl of the precipitated materials were subjected to western blot analysis with the indicated antibodies. Primary antibody was incubated with the membrane in the dilution recommended by the manufacturer. The bound primary antibody was visualized with HRP-conjugated secondary antibody using Supersignal™ West Dura system (Pierce Biotechnology, Inc., Rockford, IL, USA) and photographed using an UVITech imager (UVITech, Inc., Cambridge, UK).
PCR primer sequences.
Affinity purification of flag-tagged VEGI174 and mass spectrometry. Flag tagged VEGI174 was stably expressed in HEK293T cells. Two liters of cell culture was harvested and treated with or without 20 Gy IR. Four hours after IR treatment, cells were lysed with 40 ml NETN buffer on ice for 10 min. Cell lysates were centrifuged at 12,000g at 4°C for 20 min. Soluble fraction was collected and insoluble fraction was washed three times with PBS and then treated with 50 units of micrococcal nuclease on ice for 1 h and centrifuged at 12,000g at 4°C for 20 min. The supernatant was the chromatin fraction combined with the NETN soluble faction. Cell lysates were incubated with 500μl streptavidin conjugated beads (Amersham, Tokyo, Japan) at 4°C for 2 h. The beads were washed three times with NETN buffer, and then bead-bound proteins were eluted with 1 ml PBS containing 2mM biotin (Sigma, Poole, Dorset, UK). The eluted supernatant was incubated with 50 μl S beads (Novagen, California, US) at 4°C for 2 h. The beads were washed three times with NETN buffer and subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The gels were digested, and the peptides were analyzed by liquid chromatography-tandem mass spectrometry.
Mass Spectrometric Analysis. MALDI quadrupole time-of-flight mass spectrometry (Applied Biosystems/MDS Sciex QSTAR Pulsar) was used to identify VEGI174 associated proteins by peptide mass fingerprinting and the peptide sequence information was obtained by MS/MS analysis. The peptide masses and the amino acid sequences identified from the peptide fragmentation data were subjected to MASCOT and PROSPECTOR searching engines.
Genomic analysis of RCC tissues. For transcriptome sequencing (RNA-seq) of renal cell carcinoma tumor tissues, snap frozen samples were used to extract RNA. Its purity and integrity were checked by Agilent Bioanalyzer prior to RNA sequencing. Only RNA samples with RIN >7 and 28S/18S >1 were proceeded for library construction and sequencing. The sequencing was performed at PE125 on an Illumina HiSeq2500 platform by certified Illumina service providers. Transcript expression was estimated by MMSEQ (19) and represented by log2 (FPKM).
Immunohistochemical staining of renal specimens. Frozen specimens of renal cell carcinoma (n=16) and normal renal tissue (n=16) were cut at a thickness of 6μm using a cryostat (Leica CM 1900, Leica Microsystems UK Ltd., Buckinghamshire, UK). The nature of the samples was independently verified by two pathologists. After fixation, the sections were blocked with horse serum and probed with or without VEGI174, ACLY, ENO1 or AKR1C3 antibodies for 1 h. The secondary biotinylated antibody and the avidin-biotin complex were subsequently applied to detect VEGI expression in accordance with the Vectastain Universal Elite ABC kit protocol (Vector Laboratories, Peterborough, UK). After developing color with DAB, the sections were counterstained with Gill's hematoxylin. Staining was independently assessed by the authors.
Statistical analysis. Statistical analysis was performed using the SPSS 19.0 software. The two sample t-test was used for normally distributed data. Fisher's exact test was used for analyzing immunohistochemical staining in renal cancer tissues. Differences were considered to be statistically significant at p<0.05.
Results
Genetic manipulation of VEGI levels in HEK293T and CHO cell lines. HEK293T and CHO cells were transfected with a VEGI expression construct to create sublines showing enhanced levels of VEGI expression. VEGI174 was transfected into CHO as a positive control of HEK293T. As shown by RT-PCR analysis, VEGI mRNA expression was significantly increased in HEK293TVEGI and CHOVEGI cells compared with wild-type (Figure 1A). Western blot demonstrated that VEGI protein expression was significantly increased in HEK293TVEGI and CHOVEGI cells compared to wild-type cells (Figure 1B).
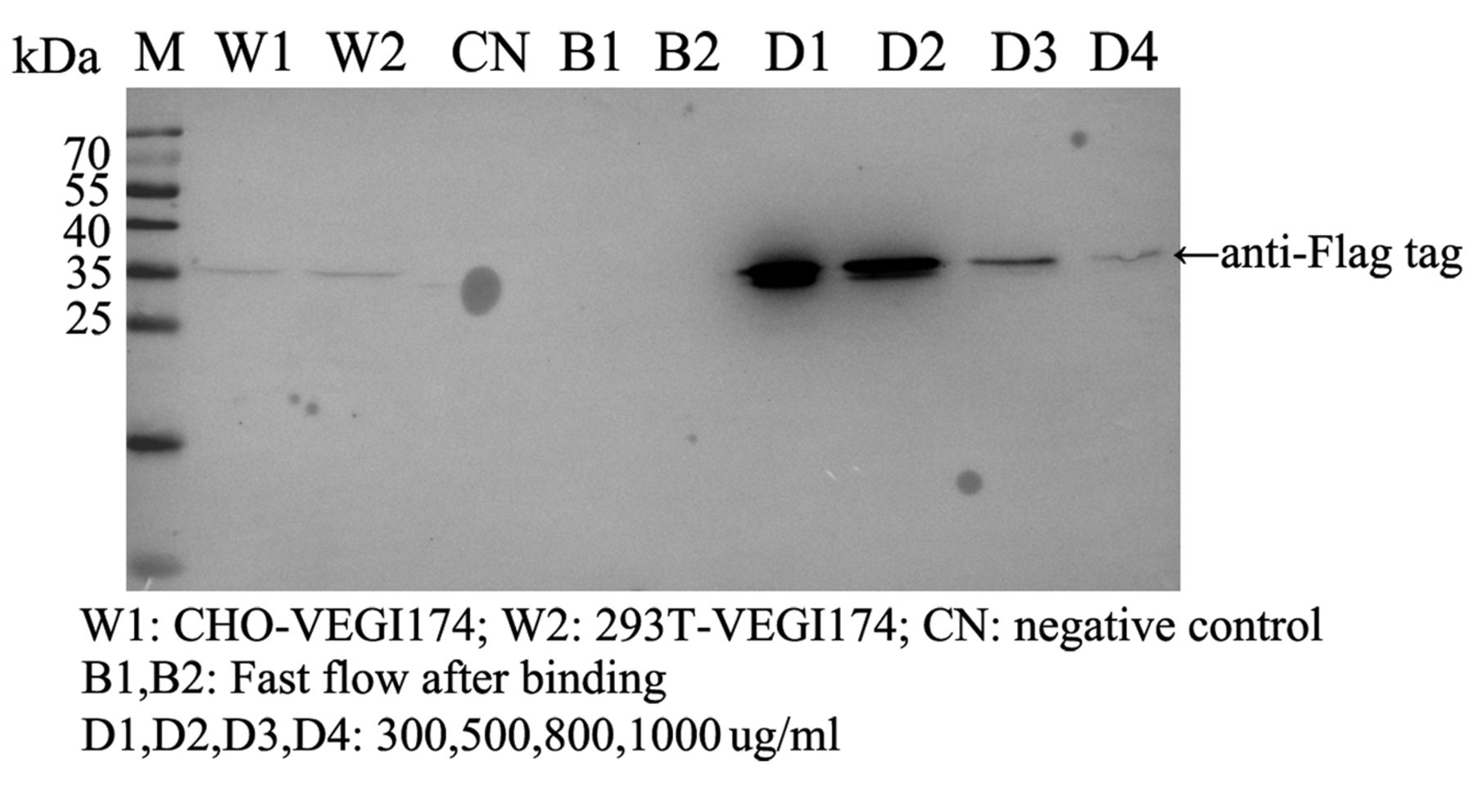
Results of mass spectrometric Analysis. VEGI174 was purified using ANTI-FLAG affinity resin. Protein samples were eluted by gradient FLAG peptide from 300 to 1,000 μg/ml (Figure 2). The gels were digested, and the peptides were analyzed by liquid chromatography-tandem mass spectrometry. The common non-specific associated proteins, including acetyl coenzyme A carboxylase 1, pyruvate carboxylase, methylcrotonoyl coenzyme A carboxylase subunit alpha, heat shock protein superfamily and myosins, are not listed in the mass spectrometry results shown in Table II. There were five proteins that may interact with VEGI174.
RNAseq analysis of renal cell carcinoma tissues. The expression of genes was examined in 16 RCC tissues by RNAseq. In the present study, we analysed more than five genes which may interact with VEGI174. Comparing against the TCGA database, the expression level of VEGI174 (−0.4626) in RCC was lower than that in normal kidney (p=0.006). The expression levels of ACLY (6.6992), AKR1C3 (6.3558), ENO1(4.330), ZIK1(0.5451) and MYC (5.2869) in RCC were higher than that in normal kidney (p-values were 0.001, 0.0045, 0.001, 0.002, and 0.001, respectively).

Confirmation of manipulation of VEGI174 expression in HEK293T and CHO cells. A: Verification of forced expression of the VEGI174 transcript in HEK293T and CHO cells. VEGI174 mRNA expression in the HEK293T and CHO cells was higher than that in wild-type cells. B: Forced expression of VEGI174 at the protein level using western blot analysis of HEK293T and CHO cells. VEGI174 protein level was increased in HEK293T and CHO cells compared with that of wild-type cells.
Five proteins that may interact with VEGI174.
In stage T1 group, the expression level of AKR1C3 was lower than that of stage T3 group, p=0.003. The expression level of ENO1 in stage T1 was higher than that of stage 3 group, p=0.029. However, there were no differences in the expression levels of VEGI, ACLY, MYC and ZIK1, p=0.114, 0.502, 0.826, and 0.822, respectively.
In addition, Fuhrman Grade 1 group and Fuhrman Grade 2 group were merged into a low-grade group. Fuhrman Grade 3 group and Fuhrman Grade 4 group were merged into a high-grade group. The low-grade group expression level of AKR1C3 was lower than that of the high-grade group, p=0.001. The low-grade group expression level of ENO1 was higher than that of the high-grade group, p=0.020. However, there were no differences in the expression levels of VEGI, ACLY, MYC and ZIK1, p=0.261, 0.578, 0.936, and 0.989, respectively.
Detection of VEGI174, ACLY, ENO1 and AKR1C3 protein by immunohistochemistry. There are many researches on MYC and ZIK1, therefore we did not detect them in this study. In immunohistochemical staining, VEGI174 was seen in the cytoplasmic area of normal renal tubular epithelia cells, but the staining was decreased or absent in RCC cells, particularly in specimens with high-grade tissues (Figure 3A). The positive staining of normal tissues (87.5%, 14/16) was significantly higher than that of RCC tissues (31.25%, 5/16), p=0.006.
ACLY was seen in the cytoplasm of normal renal tubular epithelia cells. The staining was seen in the cell membrane and cytoplasm of RCC tissues (Figure 3B). However, there was no significant difference of ACLY expression between normal renal and RCC tissues, p=0.342.

Co-immunoprecipitation of VEGI174 with its associated proteins. Co-immunoprecipitation of VEGI174 was performed by using anti-flag monoclonal antibody conjugated beads from CHO or HEK293T cells stably transfected with flag-labeled VEGI174 expression construct. The eluent was immunoblotted by anti-flag monoclonal antibody.
ENO1 was seen in the cytoplasmic and nucleus area of normal renal tubular epithelia cells. The staining was also seen in the cytoplasmic and nucleus area of RCC tissues (Figure 3C). However, there was no significant difference of ENO1 expression between normal renal and RCC tissues, p=0.117.
AKR1C3 was seen in the cytoplasm of normal renal tubular epithelia cells, but the staining was increased in the cytoplasmic and nucleus area of RCC tissues (Figure 3D). The positive staining of normal tissues (25%, 4/16) was significantly lower than that of RCC tissues (81.25%, 13/16), p=0.002. Moreover, the higher expression level of AKR1C3 was seen in higher stage and grade of RCC.
Discussion
RCC is a wide-spread oncourological disease with a tendency towards morbidity progression. At the time of RCC diagnosis, metastatic lesions are present in 20-30% of patients (20, 21). Also, in 20-30% of patients with preoperative absence of metastases, after nephrectomies metastatic lesions appear or local recurrence of disease take places (22, 23). Surgical treatment of kidney tumors is the only effective treatment modality. Until the 21st century, the efficacy of drug therapy for disseminated RCC (interferon–α, interleukins, hormonal therapy and chemotherapy) was insignificant or absent, and the median overall survival (OS) in metastatic disease rarely exceeded 8 months (24). It was only in the recent decade, after deeper understanding of the peculiarities of initiation and development of tumor progression in RCC, that it became possible to approve more efficacious drugs targeting key molecules involved in the RCC carcinogenesis.
VEGI is a multipotential cytokine that plays a role in regulating immunity, anti-angiogenesis, and inhibiting tumor growth (16, 25). Studies have shown that overexpression of VEGI could inhibit tumor neovascularization and progression in cellular and animal models. Our pre-study also showed the VEGI174 inhibitory effect on cellular motility, adhesion, vascular endothelial tube formation and tumor growth in vivo (15). It was suggested that VEGI174 functions mainly through inhibition of angiogenesis and is a negative regulator of aggressiveness during the development and progression of RCC (17).
A few mechanisms underlying the inhibitory effect of VEGI on angiogenesis were revealed. First, VEGI prevents G0/G1 endothelial cells from re-entering the cell cycle in response to growth stimuli. Second, there are two members of the TNF receptor superfamily which VEGI interacts with, namely death receptor 3 (DR3) and decoy receptor 3 (DcR3). DR3 is the functional receptor of VEGI251. It contains a death domain in its cytoplasmic tail and induces apoptosis in death receptor-3-expressing cell lines, such as in human umbilical vein endothelial cells (HUVECs). On the contrary, previous studies indicated that DcR3 is overexpressed in malignant tumors arising from the oesophagus, stomach, glioma, lung, colon and rectum (10, 18, 26-28). DcR3 enhances angiogenesis by blocking the autocrine-angiostatic function of VEGI in human umbilical vein endothelium (18). Meanwhile, anti-VEGI and -DR3 antibodies lead to increases of both cell proliferation and motility, and an induction of the formation of tube network (29). The angiogenic effect of VEGI antibody and DR3 antibody is similar to that induced by DcR3. Furthermore, VEGI induces apoptosis in actively proliferating cells, via activation of the stress protein kinases, SAPK/JNK and p38 MAPK (SAPK, stress-activated protein kinase, JNK, c-Jun N-terminal protein kinase, p38 MAPK, p38 mitogenactivated protein kinase), and the caspases, mainly caspase-3-like protease. Additionally, VEGI-induced apoptosis is attenuated by a caspase inhibitor.

Immunohistochemical staining of human renal specimens. A1, B1, C1 and D1 were normal renal tissues. The VEGI, ACLY, Eno1 and AKR1C3 protein was found to be stained in the cytoplasmic area of normal renal tubular epithelia cells. ENO1 was also seen in some nucleus areas of normal renal tubular epithelia cells. A2, B2, C2 and D2 were renal cell carcinoma tissue. A2, the staining of VEGI174 was decreased or absent in RCC cells, particularly in specimens with high-grade tissue. The positive staining of normal tissue was significantly higher than that of RCC tissues, p=0.006. B2, the staining of ACLY was seen in cell membrane and cytoplasmic in RCC tissues. C2, the staining of ENO1 was seen in cytoplasmic and nucleus area of RCC tissues. There was no significant difference of ACLY and ENO1 expression between normal renal and RCC tissues (p=0.342, p=0.117, respectively). D2, the staining of AKR1C3 was increasing in cytoplasmic and nucleus area of RCC tissues. The positive staining of normal tissue was significantly lower than that of RCC tissues, p=0.002.
However, it is still unclear whether VEGI174 acts solely or in collaboration with binding proteins in the form of a protein complex. In this study, we showed that there were five proteins that may interact with VEGI174. For MYC, it is well known that its pathway is activated in most cases of human RCC. The MYC oncogene contributes to tumorigenesis of many types of cancer through various mechanisms (30-33), including the regulation of proliferation and growth, protein and ribosomal biogenesis, changes in metabolism, lipid synthesis, and induction of angiogenesis (34-37). For Zinc finger protein interacting with K protein 1 (ZIK1), was first described in 1996 by Denisenko et al. (38). In addition to the nine contiguous zinc fingers in the C-terminus, ZIK1 contains a KRAB-A domain thought to be involved in transcriptional repression. ZIK1 demonstrated aberrant DNA methylation, it might function on tumor development (39). It was unknown whether there was a relationship between decreasing VEGI174 expression level and increasing MYC or ZIK1 expression levels in RCC tissues. There are many studies that have reported MYC and ZIK1 expression and function, thus we did not examine them with IHC.
Contrary to the decreasing expression level of VEGI174, three other factors increased in RCC tissues. The first, ACLY was an important enzyme involved in lipid biogenesis that is linked with glucose metabolism. Studies have shown a role for ACLY in tumorigenesis whereby increased levels of the enzyme leads to increased metabolic activity via activation of Akt signaling. Increasing lines of evidence suggest that enzymes involved in lipid biogenesis play a significant role in cancer cell proliferation and progression. In many cancer types such as glioblastoma, colorectal cancer, breast cancer, non-small cell lung cancer, hepatocellular carcinoma etc., the level of ACLY has been found to be quite high as compared to normal cells (40-43). In this study, mRNA levels of ACLY in RCC were higher than that in normal tissues. The IHC did not show the same result. Normal renal and RCC tissues all had positive staining. However, in RCC tissues, the staining was seen in all areas of the cell, including cell membrane, cytoplasm and nucleus. In normal renal tissues, ACLY was only seen in the cytoplasmic area of renal tubular epithelia cells. We could not explain this kind of difference.
Secondly, ENO1 generates ATP during glycolysis. Several studies have shown that ENO1 is a multifunctional protein participating in several physiological processes, including growth control, hypoxia tolerance and autoimmune activities (44, 45). Remarkably, accumulating evidence have suggested that ENO1 can function as an oncogenic protein by promoting cell proliferation, invasion and metastasis. ENO1 expression is frequently increased in diverse tumors, including head and neck, thyroid, breast, lung, prostate, colon and gastric cancer, glioma and cholangio carcinoma (46-49). Moreover, overexpression of ENO1 is positively associated with progression and poor prognosis in several tumors (49). Up to now, the role of ENO1 in RCC remains unknown. In this study, ENO1 expression was higher in RCC than that in normal renal according to the results of RNAseq and IHC. However, contrary to other tumors, the ENO1 expression level in the low stage and grade group was higher than that in the high stage and grade group.
Thirdly, AKR1C3 was originally cloned from human liver and prostate cDNA libraries (50, 51). It is a monomeric cytoplasmic protein of 323 amino acids and a molecular weight of 37 kDa (52, 53). AKR1C3 is overexpressed in numerous cancers, including cancers of the prostate, breast, uterine, blood, lung, brain and kidney, whereas its up-regulation frequently correlates with tumor invasiveness and aggressiveness (54-61). In this study, mRNA level of AKR1C3 in RCC was higher than that in normal tissues. Moreover, the higher expression level of AKR1C3 was positively correlated with higher RCC stage and grade. This is contrary to the reduction of VEGI174 in higher RCC stage and grade. IHC results also suggested this kind of correlation. So, AKR1C3 may correlate with the progression of RCC.
In conclusion, in this study, five proteins were found to interact with VEGI174. It is still unclear whether VEGI174 acts in collaboration with other binding proteins in form of a protein complex. In contrast to the decrease in VEGI174 expression in RCC tissues, the expression of the above five proteins was increased in RCC tissues. Based on other studies, overexpression of the above five proteins is associated with progression in several tumors. The exact function of those five proteins is unknown in RCC. We did not know the relationships between VEGI174 and above five proteins too. However, the results indicated that VEGI174 has more functions than we now know in the supression of the development and progression of RCC. Therefore, further studies should be performed to clarify this matter. Moreover, the expression of AKR1C3 was correlated with high-grade and high-stage RCC. It is suggested that AKR1C3 not only interacts with VEGI174, but also closely relates with the progression of RCC.
Acknowledgements
This study was supported by the National Natural Science Foundation of China, grant No. 81372138 and Cancer Research Wales (W.G.J.).
Footnotes
↵* These Authors contributed equally to this study.
Conflicts of Interest
The Authors declare no potential conflicts of interest.
- Received May 22, 2017.
- Revision received June 8, 2017.
- Accepted June 9, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}