


Abstract
Background: Intronic microRNAs (miRNAs) are considered to be transcribed using their host gene promoter. However, about one third of intronic miRNAs are predicted to have independent promoter elements. Materials and Methods: Human breast cancer cells were cultured under normoxia or hypoxia, and expression levels of intronic miR-106b-25 cluster miRNAs and their host gene minichromosome maintenance complex component 7 (MCM7) transcripts were analyzed by semi-quantitative polymerase chain reaction. The putative promoter element of miR-106b-25 cluster was analyzed by chromatin immunoprecipitation and luciferase assays. Results: Exposure to hypoxia reduced the expression of MCM7 mRNA and a primary transcript of miR-106b-25 cluster, but did not affect that of mature miRNAs. The putative promoter element of miR-106b-25 cluster was not bound by hypoxia-inducible factor 1-alpha (HIF1-α), and was not activated under hypoxia. Conclusion: Maintenance of miR-106b-25 cluster miRNA levels under hypoxia was not caused by the activation of an independent promoter element.
MicroRNAs (miRNAs) are non-coding RNAs with fewer than 25 nucleotides and function as post-transcriptional regulators of gene expression. The miR-106b-25 cluster, consisting of miR-106b, miR-93 and miR-25 miRNAs, has two paralogs in the human genome: miR-17-92 and miR-106a-363 clusters. A total of 15 miRNAs that are located within these clusters form four ‘seed sequence’ families. These miRNA clusters have oncogenic roles in cellular transformation and tumorigenesis by down-regulating several tumor suppressors, such as p21, and phosphatase and tensin homolog (PTEN) (1).
miRNAs are classified as intergenic or intronic based on their genomic locations. Intergenic miRNAs are located between independent transcription units and are transcribed from their own promoters. In contrast, intronic miRNAs are located at the intronic region of their host genes (protein-coding or non-coding genes) and share promoters with their host genes. However, the expression of miRNAs is not always concordant with that of their host genes (2), which can be attributed to the presence of independent upstream regulatory elements, differences of transcript stabilities, and/or post-transcriptional regulation of miRNA maturation processes. It is considered that about 50% of vertebrate miRNAs are intronic (3), and about 35% of intronic miRNAs have upstream regulatory elements regulated by RNA polymerase II or polymerase III (4, 5).
The miR-106b-25 cluster is an intronic miRNA cluster that is located within intron 13 of the minichromosome maintenance complex component 7 (MCM7) gene locus. MCM7 is a component of a DNA helicase that binds to replication origins and plays an essential role in DNA replication and cell proliferation. A high level of MCM7 expression is correlated with cancer progression (6). MCM7 mRNA level is reduced under prolonged hypoxia in a hypoxia-inducible factor 1-alpha (HIF1-α)-dependent manner (7). However, expression levels of miR-106b-25 cluster miRNAs do not always decrease under hypoxia.
Materials and Methods
Cell culture. MDA-MB-231 and human embryo kidney (HEK) 293T cells were purchased from the American Type Culture Collection (Rockville, MD, USA). MDA-MB-231, HEK293T and the dissociated patient-derived tumor xenograft (PDX)-breast cancer (BC)6 cells (8) were maintained in Dulbecco's modified Eagle medium supplemented with 2% fetal bovine serum, 100 U/ml penicillin, and 100 μg/ml streptomycin, and cultured with 5% CO2 at 37°C.
Primers for chromatin immunoprecipitation assays.
Semi-quantitative real-time polymerase chain reaction (PCR) assay. The abundance of each miRNA and mRNA was measured individually as previously described using the Thermal Cycler Dice (Takara, Shiga, Japan) (9). Results were normalized by the amount of small nuclear RNA, SNORD48, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) for miRNAs and mRNAs, respectively.
Chromatin immunoprecipitation assay. MDA-MB-231 cells were seeded at 1×107 cells in 10-cm tissue culture plates. Chromatin immunoprecipitation was performed as described previously using an antibody to HIF1-α (H1alpha67; ThermoFisher, MA, USA) or control normal mouse IgG (Santa Cruz, TX, USA) (10). The primer sets were designed for putative HIF1-α-binding sites within the putative promoter elements of miR-106b-25 cluster (MIR1 and MIR2), together with a negative control (MIR5) and a positive control [pyruvate kinase, muscle 2 (PKM2)] (Table I). The amount of HIF1-α-binding sequence enriched by chromatin immuno-precipitation was evaluated by PCR and subsequent agarose gel electrophoresis.
Luciferase assay. Putative promoter element of miR-106b-25 cluster was amplified by PCR using the genomic DNA of A549 cells as a template, was sequenced, and was cloned into the pMD20 T vector (Takara). The product was then cloned at the 5’ of the luciferase gene of the pGL3-basic vector. HEK293T cells and MDA-MB-231 cells were seeded at 1×105 cells per well in 48-well plates, and luciferase assays were performed as previously described (10).
Statistical analysis. Data from experiments were statically analyzed using the Student's t-test.
Results
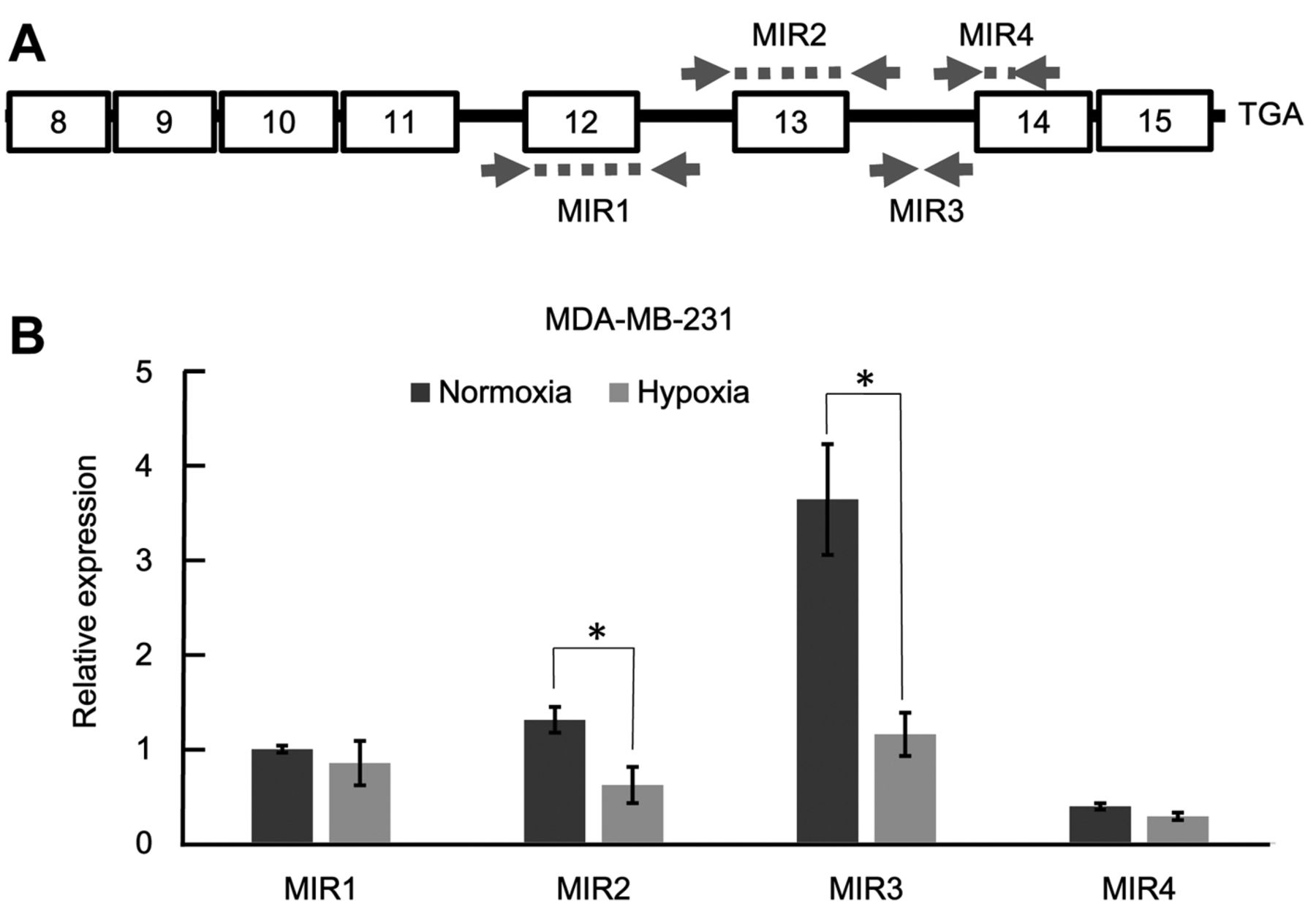
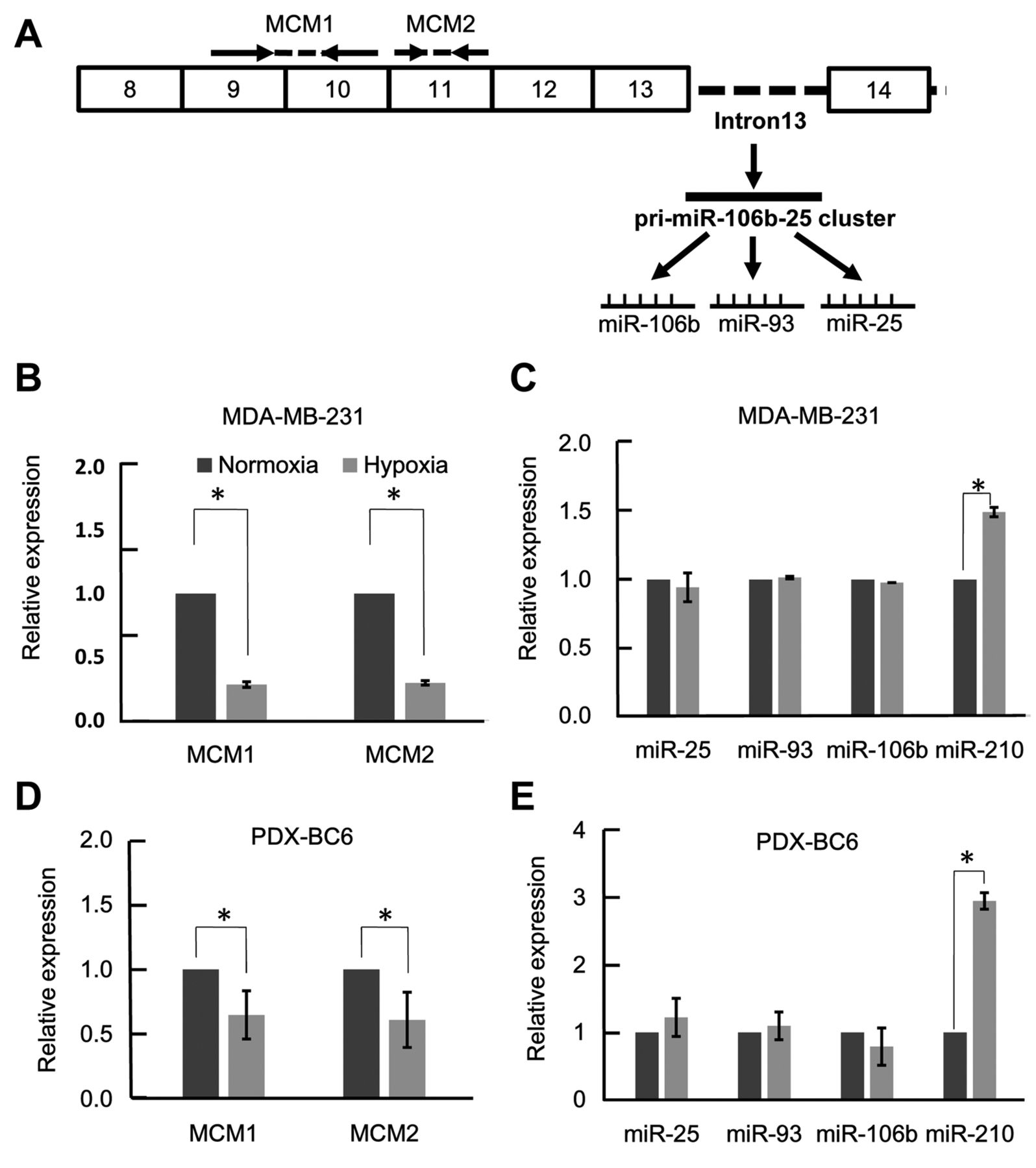
Expression of miR-106b-25 cluster miRNAs and their host gene MCM7 under hypoxia. The precursor for the miR-106b-25 cluster is located within intron 13 of their host gene MCM7 (Figure 1A). Although the presence of an independent promoter element for miR-106b-25 cluster is reported, it remains elusive under what conditions this independent promoter element functions. When MDA-MB-231 human breast cancer cells were cultured under hypoxia, the expression levels of pre-mRNA and mRNA of MCM7 were reduced by about 70% under hypoxia compared with normoxia (Figure 1B). In contrast, the expression levels of three mature miRNAs within miR-106b-25 cluster were not significantly affected under hypoxia (Figure 1C). Results were basically similar when PDX human breast cancer cells were cultured under hypoxia or normoxia (Figure 1D and E). These findings suggest that, although miR-106b-25 cluster is intronic, the expression levels of the miR-106b-25 cluster miRNAs are not concordant with that of their host gene MCM7 under hypoxia.
Reduction of the expression level of the precursor for miR-106b-25 cluster under hypoxia. The primary transcripts of miRNAs (pri-miRNAs) are first cleaved by microprocessor complex containing a ribonuclease Drosha, and further cleaved by a ribonuclease Dicer to form mature miRNAs. Consistent with the reduction of the expression level of MCM7 mRNA, the expression level of the precursor for miR-106b-25 cluster was reduced under hypoxia (MIR3 in Figure 2). Expression of the MCM7 intron containing the pri-miR-106b-25 cluster was much higher than those of other intronic regions under normoxia, suggesting that the pri-miRNA transcript was protected from degradation (Figure 2B).
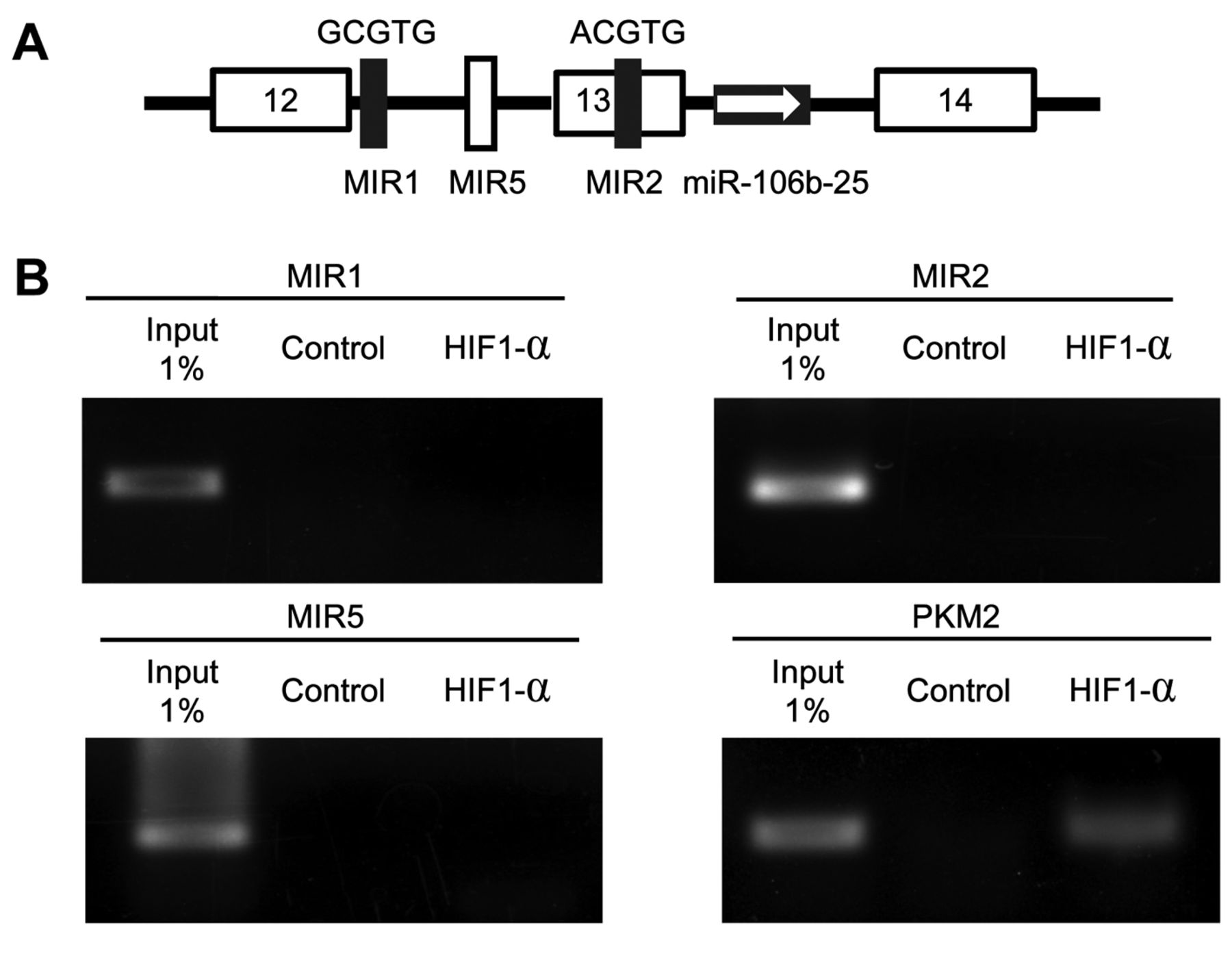
Undetectable activity of the putative promoter element of miR-106b-25 cluster under hypoxia. The presence of an independent upstream regulatory element for intronic miR-106b-25 cluster has been reported (11). There are two putative HIF1-α-binding sites within 1.4 kb of the 5’ region of pri-miR-106b-25 cluster (MIR1 and MIR2 in Figure 3A). The chromatin immunoprecipitation assays revealed that neither of these sites was bound by HIF1-α under hypoxia (Figure 3B). In addition, another two putative HIF1α-binding sites within 3 kb of the 5’ region of pri-miR-106b-25 cluster were not bound by HIF1-α under hypoxia (data not shown).
Promoter activity was then evaluated using the PGL3 luciferase plasmid in which this putative promoter element was cloned at the 5’ of the luciferase gene (Figure 4A). Activity of the putative promoter element was not significantly affected under hypoxia, and mutations in the HIF1-α-binding sequences did not affect the activity of the putative promoter element under hypoxia (Figure 4). These results suggest that the discordance of the expression levels of miR-106b-25 cluster miRNAs and their host gene MCM7 under hypoxia is unlikely to be caused by the activation of an independent regulatory element for miR-106b-25 cluster.

Expression of miR-106b-25 cluster miRNAs and their host gene minichromosome maintenance complex component 7 (MCM7) under hypoxia. A: Schematic illustration of the MCM7 mRNA and its intronic miR-106b-25 cluster. Positions of the primers for semi-quantitative real-time polymerase chain reaction assays are presented. B: Reduction of the expression level of MCM7 mRNA under hypoxia. MDA-MB-231 cells were cultured under hypoxia or normoxia for 48 h. n=3, *p<0.05. C: Maintenance of the expression levels of miR-106b-25 cluster miRNAs in MDA-MB-231 cells under hypoxia. miR-210 served as a positive control. n=3, *p<0.05. Expression levels of MCM7 mRNA (D) and miR-106b-25 cluster miRNAs (E) were measured using PDX-derived breast cancer cells. Data are the mean±standard deviation, n=3, *p<0.05.
Discussion
Hypoxia affects the transcriptional regulation of mRNA, mRNA stability, translation, and protein stability. In addition, hypoxia changes the expression levels of various miRNAs, that are represented by the up-regulation of miR-210. Expression of some intronic miRNAs are not concordant with those of their host genes and overall correlation between intronic miRNAs and host gene expression is low under hypoxia (12).
The presence of independent transcriptional elements for intronic miRNAs is one of the mechanisms that account for the discordance of the expression levels between miRNAs and their host genes. The transcription start site for the pri-miR-106b-25 is located at 79 bp upstream of miR-106b (11). However, no putative internal RNA polymerase-binding sites are found near miR-106b-25 cluster in the University of California, Santa Cruz Genome Browser. Moreover, binding of RNA polymerase to the putative promoter element is not presented. Our results showed that a putative promoter element was not functional under hypoxia (Figures 3 and 4). Therefore, hypoxia is not the factor which activates the independent transcription element for miR-106b-25 cluster.
Reduction of the expression level of the precursor for miR-106b-25 cluster under hypoxia. A: Schematic illustration of the exon-intron structure of minichromosome maintenance complex component 7 (MCM7). Intron-spanning primers were destined to measure the amounts of primary transcript of MCM7 pre-mRNA. MIR3 primers are located within the pri-miR-106b-25 cluster. B: Reduction of the expression level of miR-106b-25 precursor under hypoxia. MDA-MB-231 cells were cultured under hypoxia or normoxia for 48 h. The amount of primary transcript of MCM7 pre-mRNA was measured by semi-quantitative real-time PCR assays. Data are the mean±standard deviation, n=3, *p<0.05.

No binding of hypoxia-inducible factor 1-alpha (HIF1-α) to the putative promoter element of miR-106b-25 cluster under hypoxia. A: Schematic illustration of the putative promoter element of miR-106b-25 cluster. Positions of the putative HIF1-α-binding sites are presented. Black box: putative HIF1-α-binding site [(A/G)CGTG]. White box: negative control region. B: Chromatin immunoprecipitation for putative HIF1-α-binding sites. Lysate of cross-linked MDA-MB-231 cells cultured under hypoxia for 48 h was immunoprecipitated using antibody to HIF1-α or control mouse IgG. The template for input was purified from the 1% of total cell lysate. The HIF1-α-binding site for pyruvate kinase, muscle 2 (PKM2) was amplified as a positive control.

Activity of the putative promoter element of miR-106b-25 cluster was not up-regulated under hypoxia. A: Schematic illustration of the putative promoter element of pri-miR-106b-25 cluster. Upstream 1.4 kb region of the pri-miR-106b-25 was cloned into the 5’ of luciferase gene in pGL3 luciferase plasmid. HIF1-α-binding sequences were mutated in the pGL3-miR-Pro-Luc mutant plasmids. Black box: putative HIF1-α-binding site. Wild-type and mutated sequences are presented. B: Relative activity of luciferase gene under hypoxia. MDA-MB-231 cells were cultured under hypoxia or normoxia for 48 h. The mean of the results from the cells cultured under normoxia was set at 1.0. Data are the mean±standard deviation, n=3.
An increase of the nonsense-mediated decay variants of MCM7 that contain the intron encoding pri-miR-106b-25 is another mechanism that can cause uncoupling of the expression of MCM7 and miR-106b-25 (13). However, because the expression level of pri-miR-106b-25 cluster itself was reduced under hypoxia, contribution of the increase of nonsense mediated decay variants of MCM7 will be low under hypoxia.
Every step of miRNA production is subject to regulation. Expression levels of miRNA-processing proteins such as Drosha and Dicer are reduced under hypoxia, leading to the decrease of the expression of mature miRNAs (14). In addition, the activities of microprocessor complex and Dicer are regulated by many factors, such as adenosine deaminases, RNA-helicases, and RNA-binding proteins (15). Because hypoxia affects the transcriptional regulation of mRNA, mRNA stability, translation, and protein stability, it is possible that hypoxia maintains the expression levels of miR-106b-25 cluster miRNAs by regulating the maturation steps of miR-106b-25 cluster miRNAs.
Acknowledgements
This study was supported by Grants-in-Aid for Scientific Research from the Japan Society of the Promotion of Science, by Extramural Collaborative Research Grant of Cancer Research Institute, Kanazawa University and by grants from the Japan Foundation for Applied Enzymology and the Itoh-Chubei Foundation.
- Received May 2, 2017.
- Revision received May 26, 2017.
- Accepted June 29, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.