


Abstract
Background/Aim: Hepatocellular carcinoma is a substantial healthcare burden with high prevalence and poor prognosis. As such, efforts are continually made to uncover molecules relevant in cancer biology, that are exploitable as targets for therapy. The mitochondrion is the powerhouse of the cell and exhibits altered functionality in the malignant state, including aberrant regulation of apoptosis and cellular respiration. Augmenter of liver regeneration (ALR) is a multifunctional mitochondrial protein that demonstrates anti-oxidative and anti-apoptotic properties and plays a key role in liver regeneration. Materials and Methods: The present study systematically reviews the available literature on the role of ALR in cancer. Results: Systematic search of PubMed resulted in 12 studies discussing ALR in multiple types of cancer. More specifically, ALR appears to be up-regulated in malignant cells and tissues. Furthermore, treatment of cells with exogenous ALR shows an anti-apoptotic effect while silencing or inhibiting ALR decreases cell and tumor survival. Conclusion: ALR clearly plays a role in cancer biology and demonstrates potential as a therapeutic target.
Hepatocellular carcinoma (HCC) is a common oncologic condition with a poor prognosis that poses as a global burden on medical care (1). Between 2002 and 2012 the mortality rate of HCC rose by 34.8% in the United States and liver cancer, with HCC estimated as 70 to 90% of cases, was considered to be the second leading cause of cancer-related deaths among males on a global scale in 2012 (2, 3). Considering the metabolic aspect of several known risk factors for HCC, such as obesity, diabetes, and non-alcoholic fatty liver disease, a common underlying feature of mitochondrial dysfunction is of growing interest in molecular oncology. The regulation of specific mitochondrial proteins is of particular interest, as these proteins demonstrate potential as therapeutic targets for HCC (4).
Augmenter of liver regeneration (ALR) is one particular protein of interest that localizes in the intermembrane space of the mitochondria of humans and has a prokaryotic homolog, Erv1, sharing 42% identical amino acid residues. ALR/Erv1 has been known to contribute to mitochondrial biogenesis and function as a sulfhydryl oxidase in mitochondrial disulfide relay systems (5). It functions to oxidize the import receptor Mia40, both of which conjunctively import small translocase inner membrane (Tim) proteins through an oxidative-folding pathway (6). In turn, cytochrome c (cyt c), which constitutes an important member of the electron transport chain, oxidizes ALR through locomotion of electrons via FAD, thus connecting this redox regulated import pathway to electron transport (5, 7). During specific unfavorable conditions, cells must reprogram to undergo apoptosis during which cyt c activates a series of activated-caspase pathways that lead to cell suicide (8). It is a trademark of cancer cells to down-regulate apoptotic mechanisms and alter mitochondrial respiratory function, resulting in uncontrolled growth and eventual metastasis of cancerous cells. ALR manifests anti-oxidative and anti-apoptotic properties and is known to support liver regeneration in cases of hepatic injury (9). Considering these properties, early investigations have suggested a role for ALR in cancer (particularly hepatocellular) pathogenesis, growth and progression. In this study, we systematically review the literature discussing ALR in cancer. Clinical implications including oncogenesis and targeted therapy are explored.
Materials and Methods
A systematic review was performed using the National Library of Medicine PubMed database. The search was performed using the following key terms: cancer and augmenter of liver regeneration, growth factor erv1-like, GFER, or hepatic regenerative stimulation substance. Articles were screened by titles and abstracts for possible relevance. Subsequently, full-text articles were retrieved for final inclusion decision. Inclusion criteria were: discussion of ALR in cancer, written in English, use of human subjects/samples. Conference abstracts and literature reviews were excluded from the present study.
Summary of study details investigating ALR in cancer.
Results
Systematic search of PubMed initially returned 41 articles, shown in Figure 1. Screening by titles and abstracts eliminated 28 articles. Full-text review eliminated one article that was an editorial and did not contain original data. Ultimately, 12 articles were included in the present review (10-21). Study characteristics are summarized in Table I.
Hepatocellular carcinoma. Results of studies investigating ALR in cancer are summarized in Table II. The majority of included studies observe ALR to be upregulated in malignant cells. In frozen and paraffin-embedded HCC tissue, ALR expression is consistently altered when compared to normal para-tumoral tissue and/or healthy controls (10, 11, 13, 20, 21). The majority of studies suggest ALR mRNA and protein levels to be higher in HCC tissue relative to normal tissue. In HCC tissue, ALR typically demonstrates cytoplasmic localization with a granular quality on immunostaining (20). Dayoub and colleagues, however, observe the short ALR isoform of 15 kDa to be expressed at lower levels in HCC overall, compared to normal liver tissue (11). Furthermore, this study found ALR expression to be inversely correlated with tumor grade and histologic angioinvasion. No other studies specified the isoform of ALR investigated. Additionally, one study demonstrated serum ALR levels to be unchanged in patients with HCC (21).
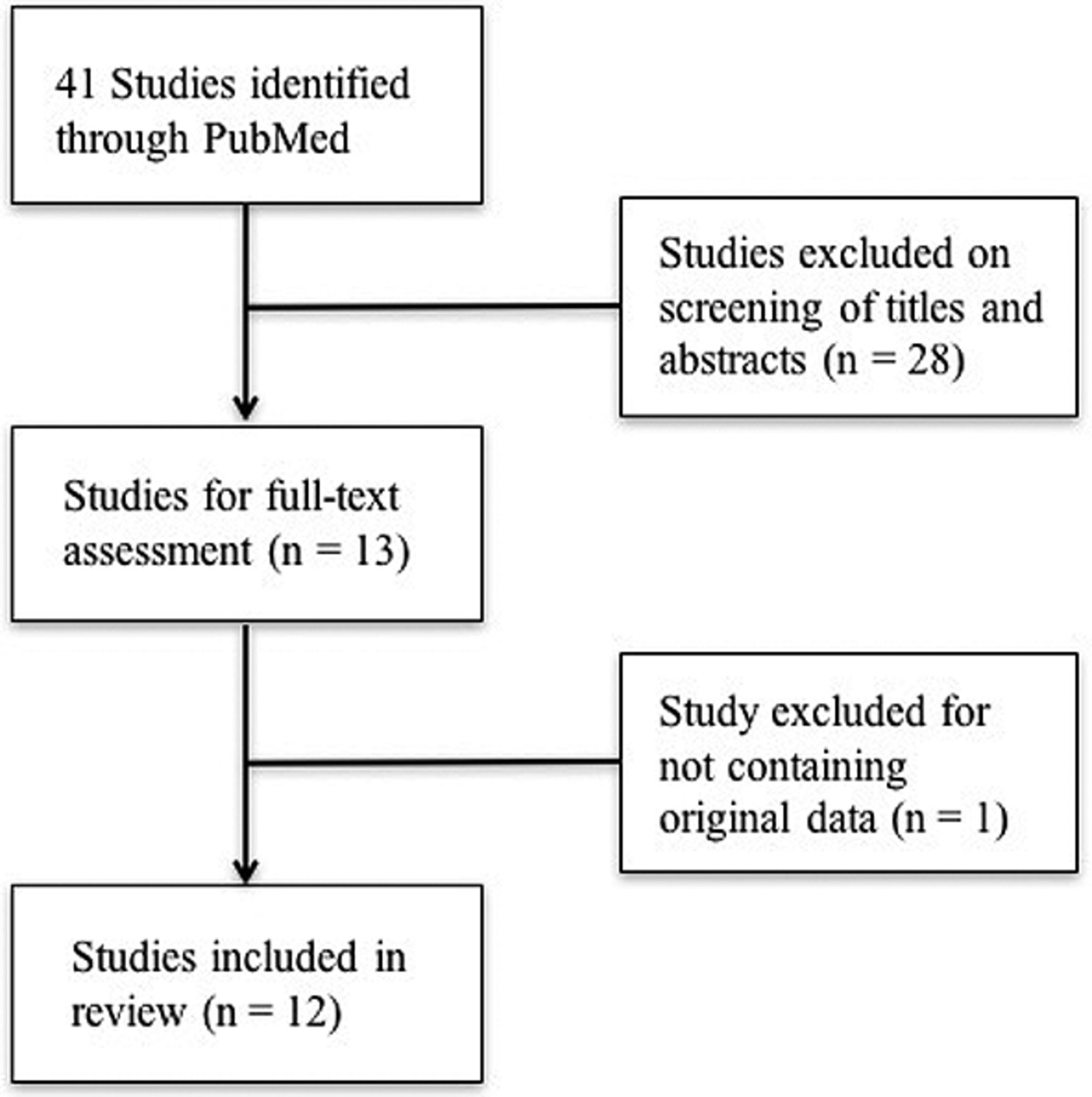
Systematic review of PubMed for articles discussing the clinical value of ALR in cancer. Initial search returned 41 articles. Screening by titles, abstracts, and full texts resulted in 12 articles remaining for final inclusion in this review.
The bulk of studies investigated the role of ALR in cancer biology using HCC cell lines. In vitro, ALR is consistently associated with a protective effect on cancer cell survival and apoptosis. Cao et al. observed overexpression of ALR in HCC cells to have a protective effect against radiation-induced mitochondrial dysfunction and subsequent apoptosis (10). Additionally, treatment of HCC cells with ALR has a protective effect against apoptosis induced by ethanol, TRAIL, anti-Apo, TGF-β, and actinomycin D (15).
Summary of clinical implications of ALR in cancer.
Silencing of ALR consistently demonstrated an anti-tumoral effect in HCC. Upon ALR silencing, overall growth and survival of HCC cells was inhibited, while a minimal effect was noted in normal hepatocyte cells (10, 19). In a murine HCC xenograft model, silencing of ALR and treatment with anti-ALR monoclonal antibodies both similarly inhibited tumor growth (19). Only one study reported that lowered ALR expression has been linked to HCC oncogenesis. In this study, mice with a liver-specific ALR knock-out were observed to have increased hepatosteatosis, hepatocyte apoptosis, and elevated HCC incidence of 60% at one year (13).
Other cancers. Five studies discussed cancers other than HCC (14, 15, 16, 17, 18). Gatzidou and colleagues observed elevated ALR levels in colorectal cancer (CRC) tissue and cell lines when compared to matched non-neoplastic tissue or controls, respectively (14). Similar to HCC, ALR expression in malignant tissue was inversely correlated with tumor grade. Namely, ALR was elevated in well to moderately differentiated tumors when compared to poorly differentiated ones. In another study, in vitro treatment with ALR does not appear to reduce apoptosis in bronchial, colonic, gastric, and pancreatic malignant cell lines (15). In T cell leukemia, cultured cells were observed to upregulate ALR compared to normal primary T lymphocytes (18). Furthermore, treatment with ALR reduces leukemic T cell sensitivity to vincristine.
ALR is of interest in neuronal tissue as well. Both neuroblastoma and glioma cells cultured with ALR exhibit a dose-dependent improvement in survival in H2O2-induced apoptosis (16, 17). Furthermore, silencing of ALR in glioma cells induces a pro-apoptotic state with increased reactive oxygen species generation and mitochondrial degeneration.
Discussion
Expression of ALR as assessed by varied qualitative and quantitative assays is clearly altered in malignancy. In light of the multifunctional nature of mitochondria, ALR likely plays a central role in cancer biology, in apoptotic regulation and cell respiration. In HCC tissues, ALR was consistently increased compared to healthy controls with the exception of one study which specifically investigated the 15 kDa isoform. Via alternative splicing, ALR may be found as a 21/23-kDa isoform predominantly in the mitochondrial inter membrane space or as a 15-kDa isoform at extracellular, nuclear, and cytoplasmic localizations (11). This likely accounts for the inconsistencies in ALR levels found across the included studies. In HCC, the malignant state may be associated with an altered expression profile of ALR that is preferential for the larger isoform. This would also explain the oncogenic effect of ALR knock-out in mice (13). If the 15-kDa isoform is associated with healthy hepatocytes while the longer isoform is upregulated in malignancy, a pan-ALR knock-out may be oncogenic. Further investigation dissecting the specific functions of the ALR isoforms in healthy cell and cancer biology is necessary.
ALR demonstrates potential as a therapeutic target in cancer treatment. Sufficient in vitro evidence suggests ALR silencing to be anti-tumoral. Silencing and/or inhibiting ALR in an HCC xenograft nude mouse model showed promising anti-tumor effect (19). However, considering oncogenic effect and increased hepatosteatosis in the murine liver-specific knock-out of ALR (13), further elucidation of the roles of the ALR isoforms must be performed. Perhaps specific targeting of the 21/23-kDa isoform may minimize adverse outcome.
Overall, the currently available literature clearly suggests an oncogenic role of ALR in cancer biology. However, the complete function of ALR in cancer is not fully understood. Further investigations need to be performed on ALR function in tumor progression and as a targeted therapy. This early evidence, however, is promising for clinical use of this molecule.
Acknowledgements
The Authors received financial support from Kadner-Pitts Research Grant, Loyola Marymount University, Los Angeles, CA, USA.
Footnotes
This article is freely accessible online.
Conflicts of Interest
None to declare.
- Received May 9, 2017.
- Revision received May 17, 2017.
- Accepted May 18, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.