


Abstract
Background/Aim: It is necessary to select an appropriate strategy that makes cancer cells more sensitive to chemotherapeutic drug(s). The aim of this study was to further explore the mechanism(s) behind drug sensitization in the breast cancer MCF7 cell line using 5-azacytidine (AzaC) pretreatment in doxorubicin (Dox) chemotherapy. Materials and Methods: Cells were treated with either AzaC or Dox alone or in combination (AzaC+Dox) for 24 and 48 h. Cells were also pretreated with AzaC for a week before replacement with Dox at varying concentrations in another group of experiments (AzaC/Dox) for 24 and 48 h. The cells in the groups were subjected to cytochemical analyses, including 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazoliumbromide (MTT), intracellular reactive oxygen species (ROS) and cell death by dual staining with Hoechst33342 (Hoechst) and propidium iodide (PI). Cells were also analyzed for determining specific alterations of possible target proteins by western blot. Results: Compared to a single (AzaC and Dox) or simultaneous drug exposure (AzaC+Dox), cellular activity was significantly reduced in AzaC/Dox group of cells in a dose- and time-dependent manner (p<0.05). The clonogenic potential of cells was also significantly inhibited in AzaC/Dox compared to Dox and AzaC group after 7 days (p<0.05). The rates of ROS-generated and PI-stained cells were significantly higher in the AzaC+Dox and AzaC/Dox groups compared to those of individual drug groups at 48 h (p<0.05). These observations were accompanied by activation of ERK1/2 and p38MAPK, as well as up-regulation of BAX and P53 in the AzaC/Dox group. However, the levels of BCL-2 were down-regulated in Dox and AzaC+Dox among a different group of cells. Elevated levels of activated caspase-3 were observed only in AzaC/Dox group of cells. The levels of phospho-nuclear factor-kappa B (pNF-ĸB)(Thr) decreased with increasing dosage of Dox in different groups, while down-regulation of heat shock protein 70 (HSP70) was observed only in the AzaC+Dox group of cells. Conclusion: AzaC significantly increases the sensitivity of MCF7 cells to Dox via activation of ERK1/2, P53, BAX and caspase-3. Moreover, the inhibition of pNF-ĸB(Thr) and HSP70 might also contribute to an increase in the sensitivity of MCF7 cells to Dox.
Breast cancer shows the highest incidence (25.2%) of the total number of deaths worldwide with considerably higher rate (43.3/100,000) than any other cancer in women (1). The main reason for this high mortality rate is due to failure of clinical treatments that target metastatic breast cancer cells, including chemotherapy and hormonal therapy, as a result of drug resistance or relapses (2). Therefore, it is important to focus on the basic mechanism(s) that lead to metastasis and mediate drug resistance when designing novel therapeutic strategies. A well-known antitumor agent, doxorubicin (Dox), is one of the commonly used drugs to treat a variety of cancers, including breast cancer and lymphoma (3, 4). Dox appears to work via intercalation into the DNA of cancer cells and the subsequent disruption of topoisomerase II-mediated DNA repair (5). Dox also produces free radicals that damage cellular membranes, DNA and proteins (5, 6). The generation of reactive oxygen species (ROS) during the reversible oxidation of Dox to semiquinone produces cytotoxicity (7). It has been reported that Dox stimulates the synthesis of ceramide, which, in turn, switches on the proteolytic cleavage of CREB3L1, followed by nuclear translocation of its NH2-terminal domain and the activation of cell cycle inhibitor genes, including P21 in human hepatoma cells (8). Dox appears to have side-effects, such as cardiotoxicity at high doses and chemoresistance of breast cancer cells at low doses (9). Therefore, alternative therapeutic strategies are sought to overcome the limitations of current cancer treatments using Dox.
Recently, strategies have been suggested to target epigenetic events, such as the de novo methylation of tumor suppressor gene promoters followed by their re-expression in different cancer cell lines (10-12). Two well-known nucleoside analogs and DNA methyltransferase inhibitors (DNMTIs), azacytidine (AzaC) and decitabine (5-AzaDc), have been shown to reverse hypermethylation-induced gene silencing of many tumor suppressor genes, thus up-regulating their expression (12, 13). AzaC, in particular, has demonstrated certain potential to induce anoikis, inhibit mammosphere formation and reduce metalloproteinase-9 activity in human breast cancer MCF7 cells (14).
In previous studies, epigenetic modulators have been used in combination with anticancer agents to increase the sensitivity of cancer cells to drug treatment (15-17). The DNMTIs and histone deacetylase inhibitors (HDACIs) alone or in combination can reverse the chemoresistance of epithelial ovarian cancer to cisplatin (18). It has also been demonstrated that DNMTIs and HDACIs with doxorubicin and cyclophosphamide can re-activate pro-apoptotic genes P53 and P21 in breast cancer patients (19). Recently, the sensitizing effects of AzaC on the activity of Dox have been reported at the miRNA expression level in human breast cancer cells (20). However, this sensitizing effect is still not clearly understood at the protein expression level in breast cancer MCF7 cells.
In this study, we evaluated how different combinations (simultaneous and sequential) at varying concentrations of the two drugs (AzaC and Dox) affect survival and proliferation of MCF7 cells by analyzing stress and apoptosis-related gene expression (at the protein level) in addition to cytochemical assays.
Materials and Methods
Cells, drugs and antibiotics. The MCF7 cell line used in this study was obtained from the American Type Culture Collection (ATTC; Manassas, VA, USA). AzaC and Dox were purchased from (Sigma-Aldrich, Seoul, Korea). Dox was dissolved in autoclaved distilled water to make a one mM stock solution, while AzaC was dissolved in 10% dimethyl sulfoxide (DMSO; Sigma) to make 5 and 10 mM stock solutions. All stock solutions were aliquoted in small volumes and stored at −70°C until use.
Cell culture and growth experiments. Cells were maintained as an adherent monolayer culture in high-glucose Dulbecco's modified Eagle's medium (DMEM; Gibco, Grand Island, NY, USA), The medium was supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS; Welgene, Daegu, Korea), 100 μg/ml of penicillin-streptomycin (Sigma), sodium pyruvate (110 mg/l) and sodium bicarbonate (3.7 g/l). Cells were grown on culture dishes (SPL Life Sciences, Seoul, Korea) and incubated under 5% CO2 in air at 37°C. The medium was changed every 48 h. The cells were regularly sub-cultured after 80% confluency by trypsinization and dispersion.
Drug treatment. Cells were treated with different concentrations of AzaC at 5 and 10 μM (AzaC) and Dox at 50-500 nM (Dox) separately or with AzaC at 5 μM and Dox together (AzaC+Dox) for 24 and 48 h. In the sequential treatment (AzaC/Dox), cells were incubated with 5 μM AzaC for 24, 36 and 48 h, as well as one week, and, subsequently, exposed to Dox (50 to 500 nM) for 24 and 48 h. Drugs were used after 24 h of cell seeding in all experiments.
Cell proliferation and drug cytotoxicity assays. Proliferation and viabilities of the cells were determined by using cell proliferation and cytotoxicity assays with 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazoliumbromide (MTT; Sigma) as described elsewhere (21). At specified periods of drug treatments, MTT assays were carried out according to the manufacturer's instructions. Cell survival rate (%) was determined by the ratio of samples' absorbance against that of the control group.
Clonogenic assay. Clonogenic assays were performed as described elsewhere (14). Cells were seeded at a density of 200 cells for each 35-mm culture plate and treated with different concentration of drugs in triplicate. The media, containing the drug(s), was refreshed every 48 h for a week. Then, cell colonies were fixed with an ice-cold methanol-acetic acid solution (3:1) for 30 min at −20°C and stained with 1% crystal violet for 30 min at room temperature. The images were recorded under a low-magnification lens (4×0.13) in the bright field of a microscope equipped with a digital camera (ECLIPSE TE2000-U; Nikon, Tokyo, Japan). The number of colonies per dish was counted and the mean number of colonies scored.
ROS assay. The cells were incubated with 0.5 μM of 2,7-dichloro fluorescein diacetate (DCFDA; Sigma) in a serum-free medium for 20 min at the end of the treatments, washed twice with phosphate buffer saline (PBS), followed by addition of fresh medium; images were recorded using a fluorescent microscope.
Hoechst and propidium iodide (PI) staining. To evaluate the effects of different concentrations on cell death, the Hoechst dye, that stains both living and dead cells, was used in combination with PI, which stains dying or dead cells. Drug-treated cells were incubated with 10 μg/ml of Hoechst (Sigma) for 20 min, washed twice with PBS followed by the addition of PI (2 μg/ml) containing fresh medium. After 5 min, PI-stained cells were observed under a Nikon fluorescent microscope (Nikon) connected to a FLEX digital camera (Diagnostic Instruments, Inc., Sterling Heights, MI, USA) using SPOT software ver. 4.6 (Diagnostic Instruments, Inc.) and all images were saved for further analysis.
Western blot analysis. Western blot analysis was performed according to a previous report (22). Briefly, cells were harvested at the end of each treatment period, washed with PBS and lysed with protease inhibitor containing lysis buffer. Protein concentration was quantified by the Bradford assay. Thirty to fifty μg of protein were separated on an 8-10% sodium dodecyl sulfate-polyacrylamide gel. After transferring the proteins to nitrocellulose membranes (Hybond ECL; GE Life Sciences, Newtown, CT, USA), they were probed with primary antibodies against BAX, B-2, caspase-3, ERK1/2, p-ERK1/2, HSP70, MAPK, P53, NF-ĸB and GAPDH (Abcam Ltd., Cambridge, UK) at a dilution of 1:1000 in 5% skim milk solution in PBS. Goat anti-mouse and goat anti-rabbit secondary antibodies (1:2000; Abcam) conjugated with horseradish peroxidase (HRP) or alkaline phosphate (AP) were used. The enzyme activities were visualized with an enhanced chemiluminescence detection reagent (Daeillab Service Co, Ltd., Seoul, Korea) or AP-substrate, nitro blue tetrazolium (NBT; Acros, Geel, Belgium) and 5-bromo-4-chloro-3-indolylphosphate (BCIP; Acros). Immunoreactivity was semi-quantified for each protein as the relative ratio normalized against GAPDH by using ImageJ ver.1.43u (NIH, Bethesda, MD, USA).
Statistical analysis. Data collected from the experiments were analyzed by using SPSS ver. 12.0 (SPSS Inc., Chicago, IL, USA) and the results expressed as mean±standard deviation (±SD). One-way analysis of variance and Duncan's multiple range tests were used for significance between the different concentrations and time intervals used. Student's t-test was used to analyze the effects of AzaC in combination with Dox on cell cycle, ROS production and nuclear fragmentation compared to the effects of Dox alone. A value of p<0.05 was considered statistically significant.
Results
Cytotoxic effects of Dox and AzaC in MCF7 cells. Despite its efficacy, a major limitation of Dox –as a cancer treatment– is the dose-dependent development of progressive dilated cardiomyopathy, which can cause heart failure (9). Therefore, it was necessary to establish the known cytotoxic effects of Dox on cells at varying concentrations and treatment times for the system employed herein. We used MTT assays to measure cellular activity at the end of the Dox treatments. In the 24 h group, a significant reduction in cell activity was observed at Dox levels of 100 nM and higher. Also, compared to the 24 h treatment, reduced cell activity was much more prominent in the 48 h groups (Figure 1A). For example, cell activity fell to 84.6% at the highest Dox level of 500 nM after 24 h, a reduction that dropped even more to 44.1% after 48 h (p<0.01). The toxicity of AzaC was also evaluated in cells at 5 and 10 μM for 24 and 48 h. In the 24 h group, no significant reduction in cell activity was recorded for both AzaC concentrations. However, cell activity dropped to 82.8% at 5 μM AzaC after 48 h of drug treatment (p<0.05, Figure 1B). After considering the direct relationship between dosage and antineoplastic effect, in conjunction with the cardiopathic effects, we designed a strategy to enhance the cell growth inhibition of Dox in cells at lower concentrations through simultaneous treatment with previously fixed dosages of AzaC. To evaluate the toxicity of drugs, we treated the AzaC+Dox group of cells for 24 and 48 h. In the 24 h group, cellular activity was found slightly lower in the AzaC+Dox-treated group compared to Dox group; however, it was not statistically significant. Cellular activity was similar after 48 h in the AzaC+Dox group (Figure 1C). After evaluating the cytotoxic activity for selected concentrations in the AzaC+Dox group of cells, cellular activity in the AzaC/Dox-treated group was checked. Compared to Dox group, significant changes in cellular activity were recorded as there was a dose- and time-dependent decrease in the AzaC/Dox group at one week (p<0.01). Cellular activities markedly decreased to 54.7% and 25.7% at 24 and 48 h at 500 nM Dox, respectively. Interestingly, significant reduction in cellular activity was found in the AzaC/Dox- compared to Dox-treated group at all concentrations used (p<0.05).
Effects of Dox and AzaC on the clonogenic potential of MCF7 cells. After observing a decrease in cellular activities in different groups at high drug concentrations, it was important to know the antiproliferative effects of each drug on the clonogenic potential of cells at lower concentrations (23). Therefore, we tested the single-cell-derived colony formation potential of the drugs under investigation in a monolayer culture for a week followed by crystal violet staining and counting of the colonies. Compared to the control group, there was a significant reduction in the size and number of colonies in the AzaC-treated group after 7 days (p<0.05). Also, the colonies in AzaC groups were scattered with no sharp clonal boundaries. The mean rates of colony formation were 87.3% in control cells, which dropped to 73.5% and 64.8% at 5 and 10 μM of AzaC group levels, respectively. No statistically significant differences were observed between the AzaC concentrations (Figure 2A and B). After evaluating the antiproliferative efficiency for AzaC group, we further evaluated the the clonogenic potential of the drugs (Dox and AzaC/Dox) at various concentrations. Marked differences were found in the morphological features of colonies between control and Dox group at different levels after a week. The cell number and size of colonies gradually decreased in the Dox group in a dose-dependent manner. The colony forming efficiency at varying levels of Dox was 54.8% at 50 nM, 58.4% at 100 nM, 49.6% at 200 nM and 3.8% at 500 nM, respectively, after 7 days. No significant differences were found at lower concentrations, while a significant difference was found at 500 nM, the highest concentration used (p<0.05). We found significant growth inhibition and highly reduced colony forming efficiency in the AzaC/Dox group of cells. The colony forming efficiency of each concentration on the AzaC/Dox group was 7.7% at 50 nM, 4.3% at 100 nM, 3.2% at 200 nM and 0.2% at 500 nM. Statistically significant differences were found in colony forming efficiency among the different concentrations in the AzaC/Dox group of cells (p<0.05). Also, the colony forming efficiency of AzaC/Dox-treated cells was significantly lower than in the Dox group of cells (Figure 2C and D; p<0.05).
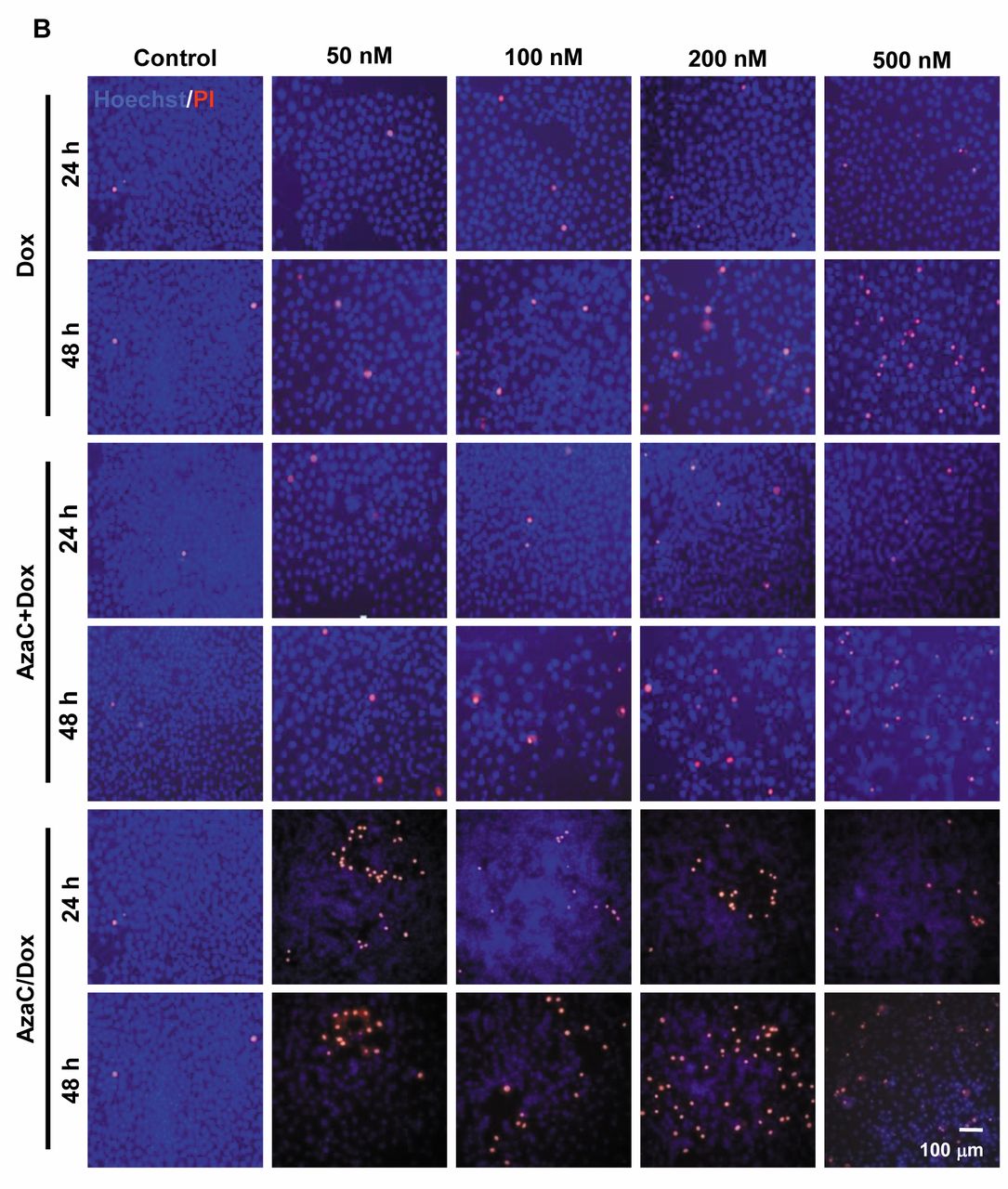
Association of oxidative stress with cytotoxicity in MCF7 cells. Next, we evaluated free radical generation in concentration- and time-dependent manner in the different groups tested by using DCFDA cell staining. ROS staining was found in cells at varying AzaC concentrations at 48 h. However, these observations were not significant. Statistically significant and dose-dependent differences were observed in Dox-treated cells. ROS production was significantly increased at different concentrations of AzaC+Dox- and AzaC/Dox-treated cells compared to Dox and AzaC groups in a dose- and time-dependent manner (p<0.05; Figure 3A). To further verify the effects of the different drug concentrations observed in a time-dependent manner on cell death, double staining with Hoechst and PI was used. The rates of PI-stained cells increased in a dose-dependent manner at different concentrations of Dox at 48 h (p<0.05; Figure 3B). No significant differences were at the two AzaC concentrations used at 24 and 48 h. Compared to Dox and AzaC groups, the rates of PI-stained cells were significantly increased in a dose- and time-dependent manner at the different concentrations of AzaC+Dox and AzaC/Dox used (p<0.05; Figure 3B). Also, in the group of cells that received high levels of AzaC/Dox, floating PI-stained cells were observed in addition to adherent stained cells after 48 h.
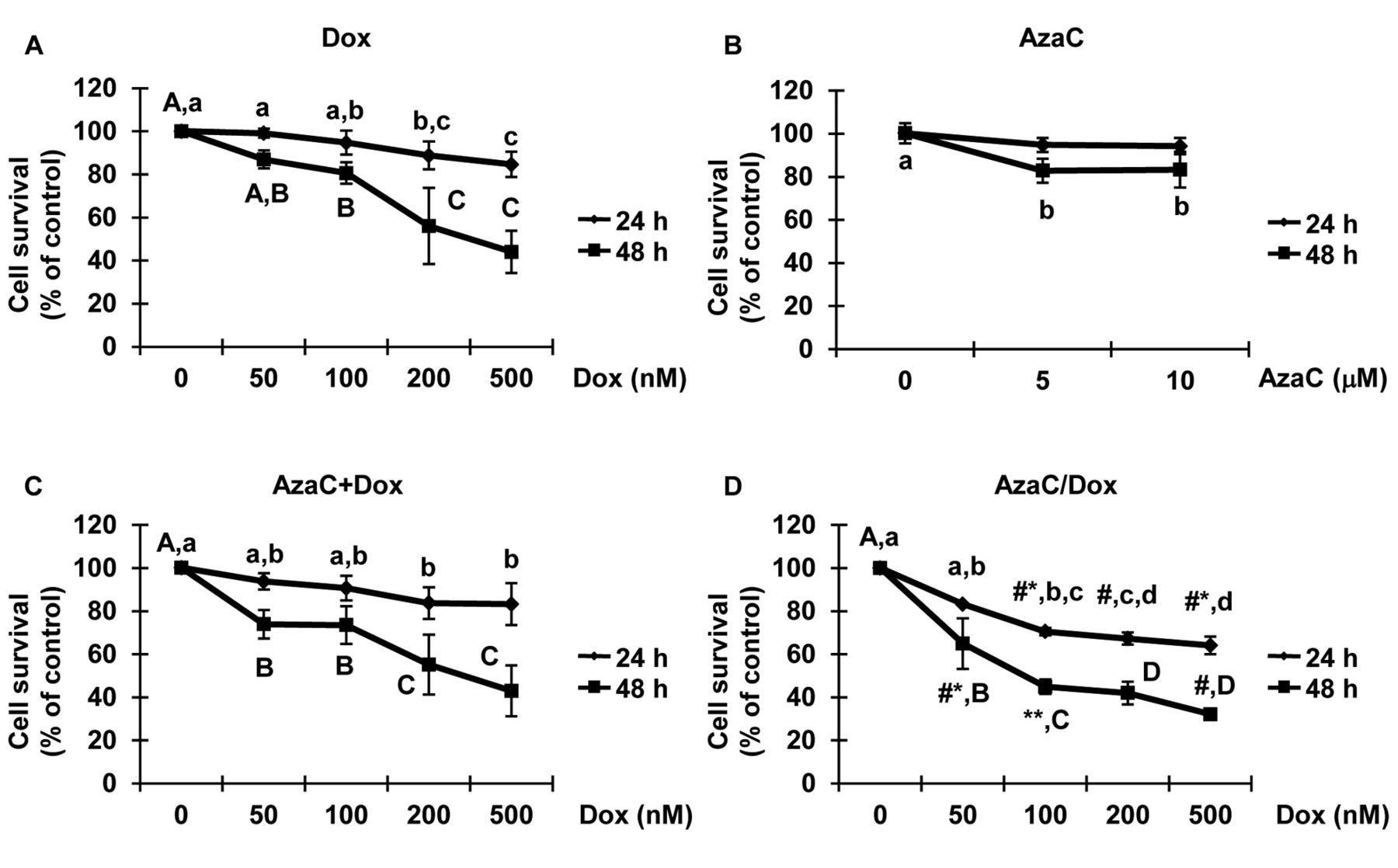
MTT assay evaluating the effects of the drugs under study on MCF7 cells. A: Cells were treated with 50-500 nM of doxorubicin (Dox), B: 5-10 μM of 5-azacytidine (AzaC), C: 5 μM of AzaC and Dox together and D: AzaC/Dox for 24 and 48 h. The data is presented as % cell viability of the control group and shown as means±SD of three independent experiments in triplicates (n=9). The #p<0.05, #*p<0.01, **p<0.001 values show statistical differences between Dox- and AzaC/Dox-treated groups when used at the same concentrations.
Effects of Dox and AzaC on protein expression levels in MCF7 cells. To investigate whether the drugs affect protein level expression, we studied the expression of stress- and cell death-related proteins in different groups by western blotting. No significant differences were found in ERK1/2 phosphorylation among different concentrations of Dox at 24 and 48 h. The activation of p38MAPK, compared to the control group, was seen at the level of 50 nM at 24 h, while, at the same time, no significant activation was observed at 48 h in the Dox-treated group of cells. P53 activation was shown in a dose-dependent manner at 48 h. The BCL-2 and phospho-nuclear factor kappa B (pNF-ĸB)(Thr) were down-regulated at different concentrations of Dox. No significant differences were recorded in HSP-70 and caspase-3 expressions; however, a gradual increase in BAX was observed in association with dosage at 48 h (Figure 4A). Next, we evaluated the effect of AzaC on protein expression. Treatment with 5 μM AzaC increased p-ERK1/2 levels in a time-dependent manner. However, no further increase was observed with 10 μM. We did not observe activation of p38MAPK at different AzaC concentrations. P53 was up-regulated at 5 μM with no further effect at 10 μM. No significant differences were found in the HSP70, pNF-ĸB(Thr) and caspase-3 expressions at the two AzaC concentrations used. Small differences in BAX and BCL-2 levels were found in a dose- and time-dependent manner (Figure 4B). Subsequently, we investigated the effect(s) of AzaC+Dox when used at different concentrations. p-ERK1/2 and p38MAPK were observed with 500 nM at 24 h, while a dose-dependent increase was found at 48 h. No significant differences were found in P53, caspase-3 and BAX expression in different levels. BCL-2 was down-regulated significantly in a dose-dependent manner at 24 h. However, no BCL-2 expression was found at 48 h. The pNF-ĸB(Thr) and HSP70 were reduced in dose- and time-dependent manner in the AzaC+Dox group of cells (Figure 4C).

Effects of the drugs under study on the clonogenic potential of MCF7 cells by crystal violet staining. A: Single-cell-derived colonies were treated with 5 and 10 μM of 5-azacytidine (AzaC) for a week, B: % colony-forming efficiency (CFE) of AzaC-treated cells, C: cells treated with 50-500 nM of doxorubicin (Dox) and D: % CFE of Dox- and AzaC/Dox-treated cells (n=3).
One of the most important parts of this study was to determine how the drugs investigated affected protein expression levels in AzaC/Dox-treated cells. The activation level of ERK1/2 and p38MAPK significantly increased in a dose- and time-dependent manner in the AzaC/Dox group compared to Dox- and AzaC- or AzaC+Dox-treated group of cells. Moreover, expression of P53, BAX and caspase-3 proteins also increased in the AzaC/Dox group of cells. No significant effects of the drugs were observed on BCL-2. Activation of NF-ĸB was reduced in a dose- and time-dependent manner. However, no significant changes were found in HSP70 levels in the AzaC/Dox-treated group of cells (Figure 4D).
Discussion
In breast cancer cells, the sensitizing role of AzaC to Dox via controlling the level of miRNA, which is responsible for cell proliferation, has been recently reported (20). AzaC also sensitizes platinum-resistant ovarian cancer cells to carboplatin via inducing caspase-3 expression (21). Unfortunately, 5-aza-2’deoxycytidine (decitabine) co-treatment with all-trans-retinoic-acid (ATRA) increases the aggressiveness of tumor in a glioma xenograft model (24). The detailed mechanism(s) of AzaC to improve the efficacy of anticancer drugs is not clear. It is important to know, however, that the anticancer role of AzaC is either dependent on cell type or coordinating cytotoxic agents in combinatorial therapies. Thus, efforts were made to elucidate the mechanism(s) of AzaC sensitization to Dox in human breast cancer MCF7 cells in this study.
The basic characteristic of an anticancer drug is to control the cellular activity of cancer cells. Therefore, we evaluated the cytotoxic effects for both Dox and AzaC alone or in combination. The differences in cytotoxicity were comparable among different groups. Interestingly, the highest cytotoxicity was found in the sequential group, i.e. when AzaC-treated cells were incubated with different concentrations of Dox for 24 and 48 h. Single-cell clonogenicity assay further confirmed the antiproliferative efficiencies of drugs at the same concentrations. Once again, compared to other groups, the percent colony forming efficiency was highly reduced in the AzaC/Dox-treated group. These results indicate that AzaC plays an important role to induce the efficacy of Dox in MCF7 cells.
The sensitizing role of AzaC to Dox was shown by evaluating the level of intracellular ROS generation in different groups. As expected, Dox generated a high level of ROS in a dose- and time-dependent way compared to AzaC group of cells. Surprisingly, the levels of ROS observed in AzaC+Dox- and AzaC/Dox-treated cells were higher compared to those of single drug groups. The Dox-induced ROS generation and cell death in various cell types through mitochondria have been previously reported (25, 26). However, the mechanism of AzaC-induced ROS production is not very obvious (27). In this study, elevated levels of ROS in combined and sequential groups further support the possible sensitizing role of AzaC to Dox in MCF7 cells.

ROS generation and cell death in doxorubicin (Dox)-, 5-azacytidine (AzaC)+Dox- or AzaC/Dox-treated MCF7 cells. A: Reactive oxygen species (ROS) production in Dox-, AzaC+Dox- and AzaC/Dox-treated cells at 24 and 48 h. B: Apoptotic cells stained with propidium iodide (PI) in Dox, AzaC+Dox and AzaC/Dox treatment groups at 24 and 48 h (n=3). DCFDA, 2,7-dichloro fluorescein diacetate.
Hoechst/PI staining further evaluated the cytotoxic and pro-apoptotic effects of the drugs under study. Similar to our preliminary results, the ratio of PI-stained cells was higher in Dox- compared to the AzaC-treated group in a dose- and time-dependent manner. Notably, the rates of PI-stained cells were higher in AzaC+Dox and AzaC/Dox groups compared to other groups. Similar to previous studies, both AzaC and Dox caused high membrane damage in AzaC+Dox- and AzaC/Dox-treated groups followed by cell death (28). The increased rates of dead or dying cells in combined and sequential groups is possibly due to AzaC sensitization to Dox in MCF7 cells.
Cytochemical observations at the protein level of molecules involved in cellular stress response and apoptosis further confirmed the above-described results. Western blot was used to evaluate the activation level of ERK1/2 and MAPK, P53 and caspase-3, BAX and BCL-2, NF-ĸB and HSP70 in the different groups studied. The majority of pro-apoptotic factors, such as ERK1/2, MAPK, P53, caspase-3 and BAX, were up-regulated in AzaC/Dox compared to other groups. The association of ERK with cell death via extrinsic and intrinsic pathways has been reported in different cell types (29). Also, the activation of p38MAPK in response to the cytotoxic drugs genistein and taxol has been studied in MCF7 cells (30). Previous reports showed that elevated level of P53 is responsible for drug-induced cell death (31). Activated caspase-3 and BAX induces chemosensitivity via release of cytochrome c, followed by cell death (29, 32). These results suggest that AzaC sensitization to Dox possibly follows the ERK/p38 and P53 signaling pathway.

Effects of drug treatment on apoptosis and stress-related factors in MCF7 cells. A: Cells were treated with either doxorubicin (Dox) or B: 5-azacytidine (AzaC) and C: AzaC+Dox or D: AzaC/Dox at the indicated concentration for 24 and 48 h. Western blot demonstrates the effects of drugs on target protein expressions.
Unexpectedly, the level of BCL-2 did not change in AzaC/Dox- as it was shown down-regulated in Dox- and AzaC+Dox-treated groups when used at different concentrations. Down-regulation of p-NF-ĸB(Thr) was equally observed in all groups except in the AzaC group. Moreover, the levels of HSP70 were reduced in a dose- and time-dependent way in AzaC+Dox and AzaC/Dox groups, respectively. Inhibition of NF-ĸB activity can sensitize the cells to drugs and induce apoptosis (33). Moreover, the cytoprotective effects of HSP70 against drugs have also been reported (34, 35). Thus, our results demonstrate that down-regulation of p-NF-ĸB(Thr) and HSP70 is possibily involved in AzaC sensitization to Dox.
In conclusion, the sensitizing effect of AzaC to Dox, as investigated by cytochermistry and western blotting, shows that (i) highest cytotoxicity and antiproliferative effects were seen in the AzaC/Dox-treated group; (ii) intracellular ROS production was significantly increased in the AzaC/Dox group; (iii) highest rates of PI-stained cells were observed in the AzaC/Dox-treated group compared to other groups; (iv) EKR1/2, MAPK, P53, caspase-3 and BAX expressions were markedly increased; and (v) NF-ĸB and HSP70 markedly decreased in the AzaC/Dox-treated group. In conclusion, we propose that AzaC has significant sensitizing effects to Dox in human breast cancer MCF7 cells.
- Received March 16, 2017.
- Revision received April 1, 2017.
- Accepted April 3, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.