


Abstract
The role of cluster of differentiation (CD) 24 in breast cancer remains unclear; previously, we showed that CD24 suppresses malignant phenotypes by inactivating Hedgehog signaling through signal transducer and activator of transcription (STAT) 1 inhibition. In this study, we examined how CD24 affects chemosensitivity in breast cancer cells. The CD44+CD24+ breast cancer cell line MCF-7 was transfected with CD24 with/without STAT1 siRNA, and chemosensitivity to 5-fluorouracil (5-FU) and cis-diamminedichloroplatinum (CDDP) was measured. CD24 inhibition reduced chemosensitivity to 5-FU, while STAT1 inhibition did not affect chemosensitivity to 5-FU in CD24 siRNA-transfected cells. Conversely, CD24 inhibition did not affect chemosensitivity to CDDP, while STAT1 inhibition reduced chemosensitivity to CDDP in CD24 siRNA-transfected cells. STAT1 inhibition, but not CD24 inhibition, reduced expression of the ATP-binding cassette (ABC) transporter genes, ABCB1 and ABCG2. In conclusion, CD24 inhibition may modulate chemosensitivity according to drug type, but ABC transporter expression appears not to contribute to this mechanism. This study contributes to determining the role of CD24 in breast cancer.
Since the identification of cancer stem cells (CSCs) in breast cancer (1), there has been a focus on the biological phenotypes of cluster of differentiation (CD) 24−/lowCD44+ cells. However, the biological role of CD24 remains unclear, and how CD24 status affects chemosensitivity also remains controversial (2-5). We previously showed that CD24 regulates proliferation and invasion via signal transducer and activator of transcription (STAT) 1 inhibition, which inactivates Hedgehog (Hh) signaling in breast cancer, and that STAT1 could be a therapeutic target (6). We also revealed that Hh signaling plays an important role in the maintenance of the CD24−/lowCD44+ population of breast cancer cells (7). Many researchers have also shown a relationship between the CD24−/lowCD44+ population and Hh signaling (8-10). Apart from regulating phenotypic changes, chemosensitivity is also an important factor for cancer treatment. ATP-binding cassette (ABC) transporters, such as ABCB1 and ABCG2, are thought to be activated in CSCs and contribute to multidrug-resistance phenotypes (11, 12). In this study, we examined how CD24 affects chemosensitivity in breast cancer cells.
Materials and Methods
Cell culture and reagents. Human breast cancer MCF-7 cell line (American Type Culture Collection, Manassas, VA, USA) was maintained in RPMI-1640 medium (Nacalai Tesque, Kyoto, Japan) supplemented with 10% fetal calf serum (FCS; Life Technologies Grand Island, NY, USA), and antibiotics (100 units/ml of penicillin and 100 μg/ml of streptomycin; Meijiseika, Tokyo, Japan). 5-Fluorouracil (5-FU) and cis-diamminedichloroplatinum (CDDP) were purchased from Kyowahakkou Kirin (Tokyo, Japan) and Bristol Myers Squibb (New York, NY, USA), respectively. The concentration of 5-FU and CDDP used in experiments was determined according to previous research (13).
RNA interference. Small interfering RNA (SiRNA) for CD24 (ON-TARGETplus™ SMART pool, L-187156), STAT1 (ON-TARGETplus™ SMART pool, L-003543), and negative control (ON-TARGETplus™ Control Non-targeting siRNA, D-001810) were purchased from Dharmacon RNA Technologies (Chicago, IL, USA). Cells (0.2×106 cells/well) seeded in 6-well plates were transfected with 100 nM siRNA using Lipofectamine RNAiMax Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Cells were used for experiments at 2 days after transfection.
Real-time reverse-transcription (RT)- polymerase chain reaction (PCR). Total RNA was extracted using High Pure RNA Isolation kit (Roche Diagnostics Gmbh, Mannheim, Germany). For real-time RT-PCR, 1 μg of RNA was treated with DNase and was reverse transcribed to cDNA using the Quantitect Reverse Transcription Kit (Qiagen, Valencia, CA, USA). Reactions were run with iQ™ SYBR Green Supermix (Bio-Rad, Hercules, CA, USA), on a DNA Engine Option 2 System (MJ Research, Waltham, MA). The primer sequences used were ABCB1, forward, 5’-TGA CAT TTA TTC AAA GTT AAA AGC A-3’, reverse, 5’-TAG ACA CTT TAT GCA AAC ATT TCA A-3’; ABCG2, forward, 5’-GCC GTG GAA CTC TTT GTG GTA G-3’, reverse, 5’-ACA GC AAG ATG CAA TGG TTG T-3’, and β-actin (ACTB), forward, 5’-TTGCCGACAGGATGCAGAAGGA-3’, reverse, 5’-AGGTGGACAGCGAGGCCAGG AT-3’. The amount of each target gene in a given sample was normalized to the level of ACTB.
Fluorescence-activated cell sorting (FACS) analysis. To analyze the expression of the surface molecules on MCF-7 cells, cells were incubated for 30 min with fluorescein isothiocyanate-conjugated monoclonal antibody to CD24 (BD Pharmingen, San Diego, CA, USA) and phycoerythrin (PE)-conjugated antibody to CD44 (BD Pharmingen). Mouse IgG was used as isotype controls (BD Pharmingen). The fluorescence intensity was measured using a FACSCalibur flow cytometer (BD Pharmingen) and analyzed with CELLQuest software (BD Pharmingen).
Statistical analysis. Data are presented as the means±standard deviation (SD). Student's t-test was used to compare continuous variables between two groups. A p-value of less than 0.05 was considered statistically significant.
Results
CD24 inhibition reduced chemosensitivity of MCF-7 cells to 5-FU. Since the MCF-7 cell line primarily consists of CD24+ cells, MCF-7 cells were selected for this study. We first confirmed that transfection of MCF-7 cells with CD24 siRNA significantly reduced CD24 expression (Figure 1). Next, chemosensitivity to 5-FU was measured in control MCF-7 cells, CD24 siRNA-transfected MCF-7 cells and CD24 and STAT1 siRNA-co-transfected MCF-7 cells. 5-FU chemosensitivity of CD24 siRNA-transfected MCF-7 cells was significantly lower compared to control cells on both day 1 and 2 (Figure 2). STAT1 inhibition did not significantly affect chemosensitivity to 5-FU of CD24 siRNA-transfected MCF-7 cells (Figure 2).
CD24 inhibition did not affect chemosensitivity to CDDP in MCF-7 cells. Next, the chemosensitivity to CDDP was investigated in the same cells. While chemosensitivity to CDDP in CD24 siRNA-transfected MCF-7 cells increased compared with controls on day 1, there was no significant difference in chemosensitivity on day 2 (Figure 3). STAT1 inhibition significantly reduced chemosensitivity to CDDP in CD24 siRNA-transfected MCF-7 cells (Figure 3).
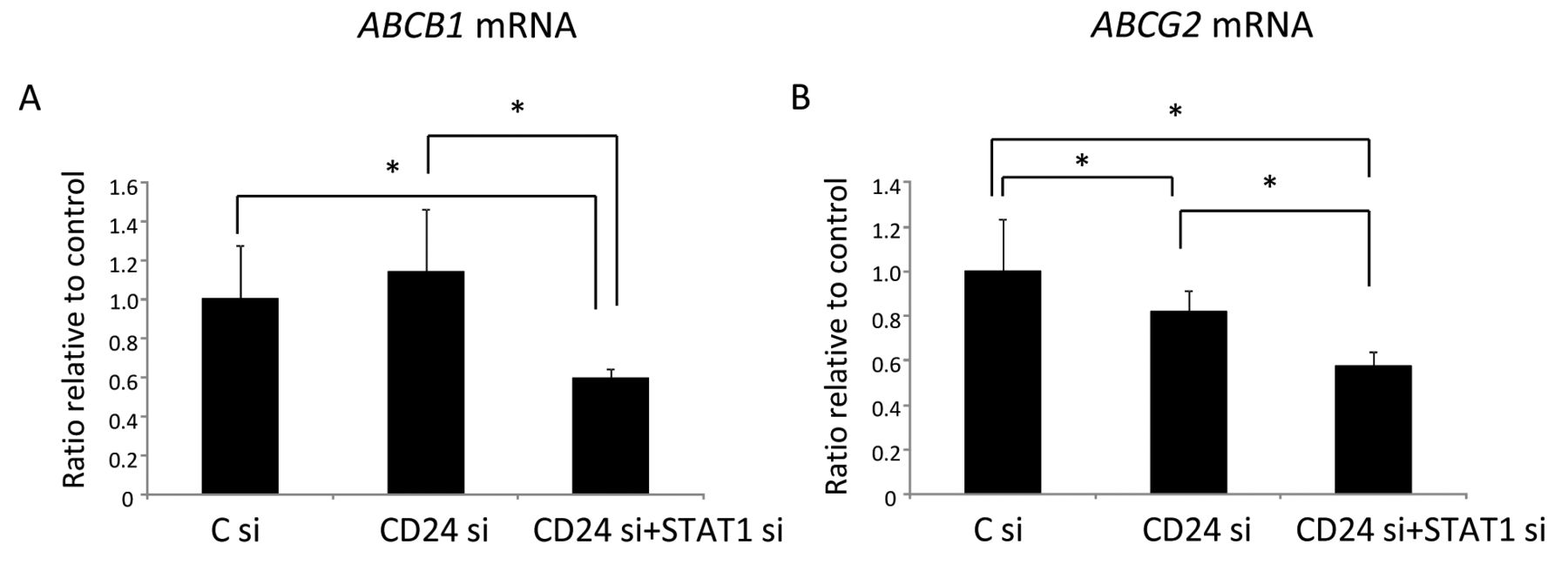
STAT1 inhibition but not CD24 inhibition decreased ABCB1 and ABCG2 expression. ABCB1 and ABCG2 are involved in drug resistance, playing pivotal roles in efflux of chemotherapeutic drugs (14-16). Therefore, expression of ABCB1 and ABCG2 in control siRNA, CD24 siRNA-transfected and CD24 and STAT1 siRNA-co-transfected MCF-7 cells was measured. There was no significant change in ABCB1 and ABCG2 mRNA expression in CD24 siRNA-transfected MCF-7 cells compared with control cells. However, expression of ABCB1 and ABCG2 in CD24 and STAT1 siRNA-co-transfected MCF-7 cells significantly decreased compared with CD24 siRNA-transfected MCF-7cells (Figure 4).

Expressions of cluster of differentiation (CD) 24 and CD44 on control (Cont) siRNA- and CD24 siRNA-transfected MCF-7 cells were investigated by fluorescence-activated cell sorting analysis.
Discussion
In this study, STAT1 inhibition led to a decrease in ABCB1 and ABCG2 expression in CD24-inhibited MCF-7 cells. An augmentation of chemosensitivity is anticipated in this situation; however, chemosensitivity to 5-FU did not increase, and conversely, chemosensitivity to CDDP decreased (Figures 2 and 3). One potential mechanism for these results is that the percentage of cells in S phase might be decreased by Hh signaling inactivation, which results from STAT1 inhibition; 5-FU is an S phase-sensitive drug, and therefore may not have been as active, as was previously shown (13). Our results are consistent with the findings of Kaewpiboon et al. who showed that STAT1 up-regulation affects etoposide resistance through ABCB1 expression (17). The activity of ABC transporters may differ depending on drug type. CDDP is a cell cycle-independent drug, and we have no clear hypothesis regarding why chemosensitivity to CDDP did not increase concomitantly with decreasing ABC transporter expression. There may be other genes or molecules that were affected by STAT1 inhibition. Furthermore, STAT1 inhibition has been shown to reduce proliferation of and invasion by MCF-7 cells (6). Therefore, STAT1 may be a therapeutic target in breast cancer, but we caution should be exercised in cases of combination chemotherapy.

MCF-7 cells transfected with control siRNA (C si), CD24 siRNA, and both CD24 and STAT1 siRNA were treated with 5-fluorouracil (5-FU) at the indicated concentration. After 1 and 2 days' culture, viable cells were counted under light microscopJ. *Significantly different at p<0.05.

MCF-7 cells transfected with control siRNA (C si), CD24 siRNA, and both CD24 and STAT1 siRNA were treated with cis-diamminedichloroplatinum (CDDP) at the indicated concentration. After 1 and 2 days' culture, viable cells were counted under light microscopy. *Significantly different at p<0.05.
Importantly, the CD24−/lowCD44+ population itself are not breast CSCs; breast CSCs cannot be induced by CD24 inhibition alone. Indeed, not all Hs578T cells, which are mainly CD24−, have stem cell-like features (6). Whether the relationship between CD24 expression and various pathological features or cell phenotypic changes is causal or the result of indirect interactions should be carefully studied.

ATP-binding cassette sub-family B member (ABCB)1 (A) and ABCG2 (B) mRNA expression in MCF-7 cells transfected with control siRNA (C si), CD24 siRNA, and both CD24 and STAT1 siRNA were estimated by real-time polymerase chain reaction. *Significantly different at p<0.05.
In conclusion, we examined the role of CD24 in breast cancer cells, especially focusing on chemosensitivity. CD24 inhibition may modulate chemosensitivity according to drug type, and ABC transporter expression might not contribute to the molecular mechanism involved in this. This study helps determine the cellular function of CD24 in breast cancer and breast CSCs.
Acknowledgements
This study was supported by the Japan Society for the Promotion of Science Kakenhi Grant Number JP26293289. The Authors thank Ms Kaori Nomiyama for skillful technical assistance.
Footnotes
Conflicts of Interest
The Authors declare no conflict of interest in regard to this work.
- Received November 18, 2016.
- Revision received December 13, 2016.
- Accepted December 20, 2016.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.