


Abstract
Background: Feasibility and efficacy of boric acid (BA)-mediated boron neutron capture therapy (BNCT) was first demonstrated by eliminating hepatocellular carcinoma (HCC) in a rat model. Furthermore, selective uptake of BA by liver tumor cells was shown in a rabbit model. To gain further insight, this study aimed to investigate the mechanisms of transportation and selective uptake of BA in HepG2 liver tumor cells. Materials and Methods: Transportation of BA in HepG2 cells was analyzed by time-course assays and by analyzing the rate of diffusion versus the concentration of BA. The effect of different tumor conditions on BA uptake was studied by treating HepG2 cells with 25 μg 10B/ml BA under different concentrations of glucose, at different pH and in the presence of water-soluble cholesterol. Results: HepG2 cells mainly uptake BA by simple diffusion. Cell membrane permeability may also contribute to tumor-specific uptake of BA. Conclusion: The selective uptake of BA was achieved primarily by diffusion, while other factors, such as low pH and increased membrane fluidity, which are hallmarks of HCC, might further enhance BA uptake.
- Hepatocellular carcinoma (HCC)
- boron neutron capture therapy (BNCT)
- boric acid
- transportation
- cell membrane permeability
- acidic microenvironment
Hepatocellular carcinoma (HCC) is one of the most common types of cancer worldwide (1). While new treatments are continually evolving, HCC remains a significant cause of death, especially in people with cirrhosis.
Current clinical guidelines suggest treating HCC with resection, transplantation, radiofrequency ablation, transcatheter arterial embolization, or administration of the kinase inhibitor sorafenib (2, 3). With these treatment options, the 5-year survival rates of patients who underwent liver resection and transplantation for early-stage HCC are 50-70% and 70-80%, respectively (4). However, more than 80% of patients with HCC are diagnosed with intermediate- or advanced-stage disease and are not surgical candidates due to either multifocal disease occurrences or vascular invasion (5). Such patients will receive palliative care and have a poor 5-year survival rate.
For treating advanced HCC, the multikinase inhibitor sorafenib (also known as Nexavar) is the only United States Food and Drug Administration-approved drug available today. While sorafenib has been shown to improve survival rate in patients with unresectable disease by approximately 3 months (6), it has various adverse effects, including poor liver function and diarrhea. Additionally, some patients with HCC do not respond to the treatment (7). As a result, the clinical challenge for advanced HCC is to find an effective treatment with tolerable side-effects.
Boron neutron capture therapy (BNCT) could be a promising treatment option for HCC. BNCT already has proven its efficacy in various human cancer types, including head and neck cancer, melanoma, and malignant glioma (8-10). In terms of its mechanism of action, BNCT makes use of thermal or epithermal neutrons to irradiate tumors that have been previously loaded with boron drug containing the stable 10B isotope. The resulting reaction generates 4He and 7Li particles within the tumor cells. Because of the short reaction range and high linear energy transfer, nearly all the energy is released inside the tumor. As a result, the tumor cells are eliminated, while the neighboring normal cells remain unharmed.
One challenging aspect of BNCT is to find a boron drug that can be selectively absorbed by the tumor cells. The available boron drugs for BNCT are borocaptate sodium (BSH) and boronophenylalanine (BPA). However, these are not suitable for HCC treatment because of the low tumor-to-normal (T/N) uptake of BSH in liver tumors and the high accumulation of BPA in the pancreas (11-13).
In recent years, the therapeutic efficacy of boric acid [BA; B(OH)3] as the boron drug for BNCT has been re-examined. BA-mediated BNCT has been demonstrated to have promising efficacy and safety in xenograft models of hepatoma in rats and rabbits in studies performed by Chou et al. (14, 15). In these preclinical studies, the research team demonstrated that the selective accumulation of BA in liver tumor cells and tumor blood vessels could lead to rapid reduction in tumor size and destruction of the tumor vasculature after neutron irradiation. However, the mechanism of selective uptake and accumulation of BA in liver tumors remains unclear.
Previous studies have indicated that BA is not metabolized in animals and is mainly transported by simple diffusion (16, 17). Such findings also imply that the observed preferential accumulation of BA in tumor tissues might be because of the differences between normal and tumor tissues, including differential carbohydrate uptake, energy metabolism, cellular membrane lipid composition, and pH value of the microenvironment (18, 19).
On the other hand, cancer cells are known for their high glucose uptake, in which glucose transporter activity can be 10- to 12-fold higher than that in normal cells (20). BA can form complexes with cis-diols containing carbohydrates, such as glucose, mannose and galactose, and BA–carbohydrate interactions are reversible in aqueous solution (21). Therefore, it is possible that the selective accumulation of BA in tumors might result from BA–carbohydrate interactions, during which sugars might help prevent BA from diffusing out of cancer cells.
Meanwhile, lipid composition was shown to impact BA diffusion and transportation in an artificial membrane by changing the permeability coefficient of BA (22). Interestingly, the lipid composition of the plasma membrane in cancer cells is different from that in normal cells (23). For instance, triglyceride and cholesterol levels are decreased in hepatoma, especially in advanced hepatomas, which might relate to increased plasma membrane fluidity (24). Thus, it is possible that enhanced cell permeability is due to the differential lipid composition of hepatoma cells compared to normal liver cells, and this might play an important role in the selective uptake of BA by tumor cells.
Last but not the least, the immediate tumor microenvironment has been shown to be more acidic than the surrounding region. The pH values in tumor blood vessels and the tumor microenvironment have been found to be between 5.0-6.0 and 6.5-6.8, respectively (25). An acidic microenvironment could affect several membrane channels, including sodium channels and calcium channels (18). Moreover, an acidic microenvironment might contribute to higher cell membrane fluidity, which is positively correlated with cell membrane permeability (26). Consequently, it is possible that an acidic tumor microenvironment might contribute to the selective accumulation of BA inside tumor cells.
In this study, we aimed to clarify the mechanism of BA uptake in hepatoma cells reported in previous in vivo studies (14, 15). We investigated whether glucose concentration, pH value, or cell membrane lipid composition could regulate the selective uptake of BA in HepG2 hepatoma cells. Our findings are critical for planning future BA-BNCT clinical trials for advanced HCC.
Materials and Methods
Cell culture. HepG2 human hepatocellular carcinoma cells (ATCC® HB-8065; Manassas, VA, USA) were cultured in high glucose (4.5 g/l) Dulbecco's modified Eagle's medium (DMEM; Gibco, El Paso, TX, USA) containing 10% heat-inactivated fetal bovine serum, 100 unit/ml streptomycin, 100 μg/ml penicillin, 0.25 μg/ml fungizone and 3.7 g sodium bicarbonate at 37°C in ambient air with 5% CO2.
Preparation of BA solution. 10B-Enriched BA (99% 10B) was used as the boron drug in this study and was purchased from Aldrich Inc. (Darmstadt, Germany). BA powder was dissolved in phosphate-buffered saline (PBS) to yield the requiring 10B concentration, and then processed with a 0.22 μm filter to sterilize the solution. The stock solution of BA (6,000 μg 10B/ml) was stored at 4°C. (Amounts of boron in the solution or sample were expressed as μg B.)
Chemical treatment. For treatment, 2×106 HepG2 cells were seeded and cultured in 10 cm cell culture dishes. Medium with pH 5.4 was prepared by hydrochloric acid titration. For the pH 5.4 treatment group, cells were cultured for 48 h in pH 7.4 medium, rinsed with 2 ml of pH 5.4 medium and then cultured in pH 5.4 medium during 25 μg 10B/ml BA treatment for 5, 30 and 180 min.
For cell membrane fluidity assays, water-soluble cholesterol (Sigma, St. Louis, MO, USA) was used to change the cell membrane composition and to increase lipid rafts in cell membrane as described previously (27). Briefly, cells were cultured in glucose-free DMEM containing 10% heat-inactivated fetal bovine serum for 47 h, treated with different concentrations of water-soluble cholesterol for 1 h and then treated with 25 μg 10B/ml BA for 5 min.
For glucose treatment, cells were cultured in glucose-free DMEM for 24 h. After glucose starvation, the medium was replaced with fresh medium with different glucose concentrations, and cells were then cultured for 48 h. Afterward, cells were treated with 25 μg 10B/ml BA for 180 min.
For co-treatment, 3 mM glucose and BA were added together during 180-min incubation. For 4°C treatment, cells were cultured for 48 h, the medium was removed and the cells were transferred into an incubator at 4°C for 10 min. Afterward, cells were treated with culture medium containing 25 μg 10B/ml BA at 4°C for 5 min.
Cell harvest. After treatment, cells were washed with 5 ml PBS at 4°C three times. The cells were collected in 1 ml PBS after scraping. The resulting cell suspension was transferred into Eppendorf tubes and centrifuged at 200 rcf for 10 min. The supernatant was aspirated and transferred to a new tube, and the cell pellets were weighed. After weighing the cell pellets, the cells and supernatant were mixed for boron quantification.

Time course of the boron uptake of HepG2 cells treated with boric acid (BA). HepG2 cells were cultured for 48 h and then treated with 25 μg 10B/ml BA for different times. Data are means±SD (n=3-6).
Boron quantification by inductively coupled plasma-atomic emission spectrometer (ICP-AES). A dissolved sample was required for atomic emission measurements. A MLS 1200 Milestone microwave digestion system (Milestone s.r.l., Sorisole, Italy) was used to dissolve cells into the aqueous solution and to make samples suitable for ICP-AES analysis. For microwave digestion, a mixture of cells and supernatant, 2 ml of concentrated nitric acid (HNO3; 14N=65%) and 0.5 ml of hydrogen peroxide (30-35%) were placed into a Teflon high-pressure digestion vessel which was then sealed. The vessels were placed into a microwave digestion system for the decomposition process. The decomposition process consisted of three sequential stages, including 3 min at 300 W, 2 min at 600 W and 20 min of cooling and depressurizing. Samples were then diluted with deionized water after which boron concentrations were measured by ICP-AES (OPTIMA 2000 DV; PerkinElmer Instruments, Norwalk, CT, USA). Boron levels were measured at a wavelength of 249.772 nm (28).
Statistical analysis. All data are expressed as means with standard deviations (mean±SD). Student's t-tests were used to analyze the data. A p-value was considered statistically significant when it was less than 0.05.
Results
Mechanistic analysis of BA transportation in HepG2 cells. To analyze HepG2 cell uptake of BA over time, HepG2 cells were incubated with 25 μg 10B/ml and BA was assayed at different time points. As shown in Figure 1, the boron concentration in HepG2 cells rapidly increased within the first 5 min and remained at a stable level from 10 to 12 μg/g for 240 min. These data suggest that BA likely enters the tumor cells via simple diffusion. For further confirmation of the simple diffusion mechanism, we repeated the assay at two different temperatures: 37°C and 4°C. HepG2 cells were treated with 6.25 to 1,000 μg 10B/ml BA at either 37°C or 4°C for 5 min before harvesting.
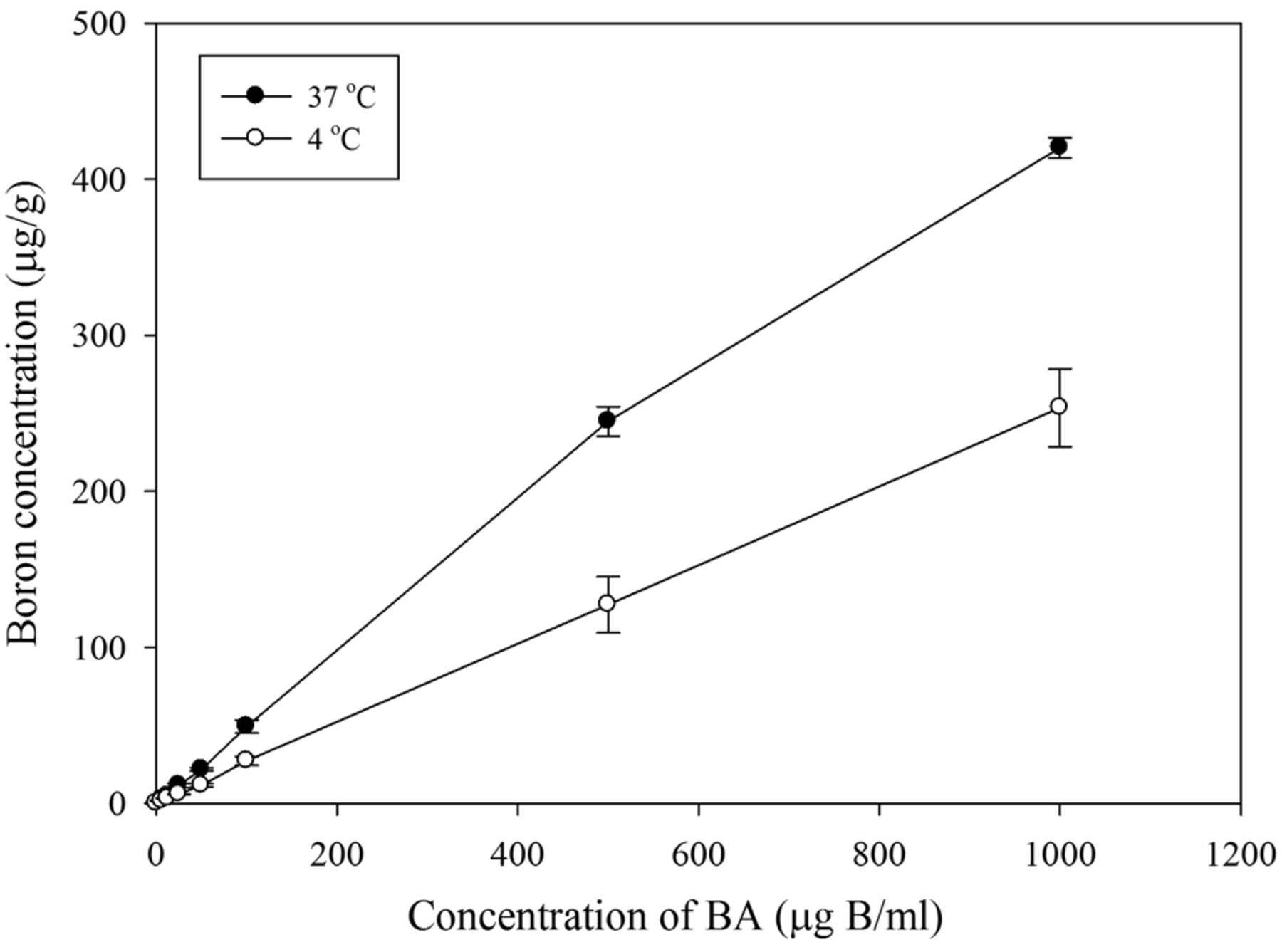
Boron uptake of HepG2 under different boric acid (BA) concentrations and temperature conditions. HepG2 cells were cultured for 48 h and treated with different concentrations of 10B/ml BA for 5 min at 37°C or 4°C. The slope of the trend line for the 37°C group was 0.4346 (R2=0.994). The slope of the trend line for the 4°C group was 0.254 (R2=0.9999). Data are means±SD (n=3).
As shown in Figure 2, we found a linear correlation between the boron concentrations at both temperatures. The slope at 37°C and 4°C was determined to be 0.43 (R2=0.99) and 0.25 (R2=1.00), respectively. The diffusion rate of BA at 4°C appears to be lower than at 37°C. Based on Fick's law of diffusion, these data further support our belief that simple diffusion is the primary mechanism of BA uptake in HepG2 cells.
Effect of glucose–BA interaction on uptake of BA in HepG2 cells. To examine whether altered glucose level could influence the uptake of BA, after 24 h glucose starvation, HepG2 cells were cultured in medium with different glucose levels for 48 h. The cells were then treated with 25 μg 10B/ml BA for 3 h to ensure sufficient uptake. We also included a co-treatment group (3 mM glucose plus 25 μg 10B/ml BA) for comparison. As shown in Figure 3, we found no significant difference in boron uptake with glucose pretreatment or co-treatment. These data indicate that glucose level and glucose influx appear not influence the accumulation of BA in HepG2 cells.

Boron uptake of HepG2 cells under different glucose concentrations. HepG2 cells were glucose starved by culture in glucose-free medium for 24 h. After starvation, cells were treated with glucose-free, 3 mM glucose or 15 mM glucose for 48 h and then treated with 25 μg 10B/ml boric acid (BA) for 3 h. For one of the glucose-free groups, 3 mM glucose was added during the BA treatment. Data are means±SD (n=3).
Effect of an acidic microenvironment on uptake of BA in HepG2 cells. To explore whether cancer cells could uptake more BA under acidic conditions, HepG2 cells were treated with 25 μg 10B/ml BA for 5, 30, and 180 min in either pH 7.4 or pH 5.4 medium to mimic the reported acidity levels of the tumor microenvironment. As shown in Figure 4, we found no statistically significant increase in BA uptake within the same pH group; however, there was a slight increase in boron level at pH 5.4 with a longer incubation time. However, due to large standard error values for the pH 5.4 group, we concluded that the data only suggest the possibility that an acidic environment might weakly influence the uptake of BA in cancer cells.
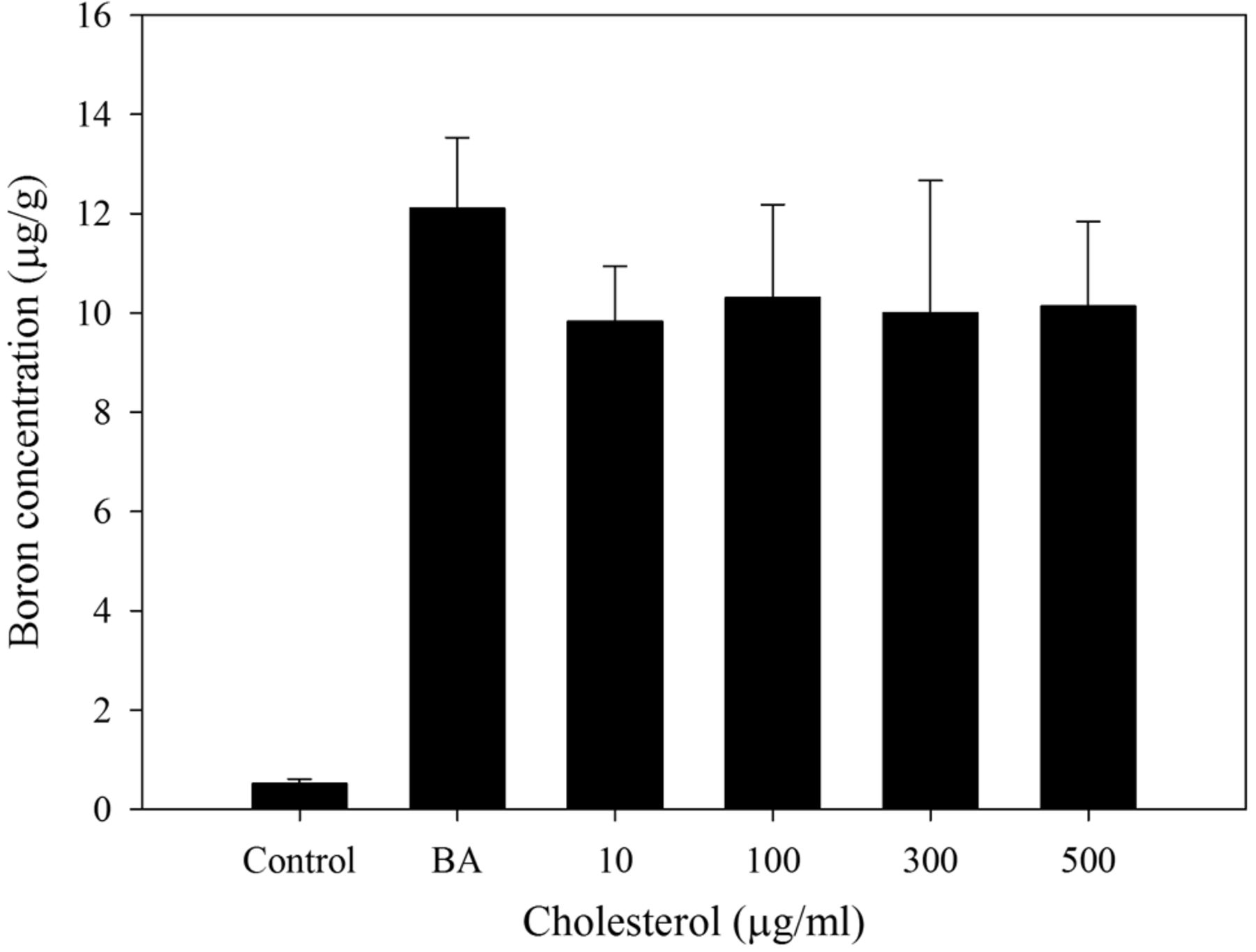
Effect of cell membrane fluidity on uptake of BA in HepG2 cells. To further investigate whether cell membrane fluidity can affect BA uptake in cancer cells, HepG2 cells were pretreated with 10, 100, 300, or 500 μg/ml water-soluble cholesterol to alter their cell membrane fluidity. The cells were then treated with 25 μg 10B/ml BA for 5 min before cell harvesting and boron quantification. As shown in Figure 5, we observed no statistically significant differences in BA uptake. However, the data suggest that cell membrane fluidity might weakly impact the uptake of BA in HepG2 cells.
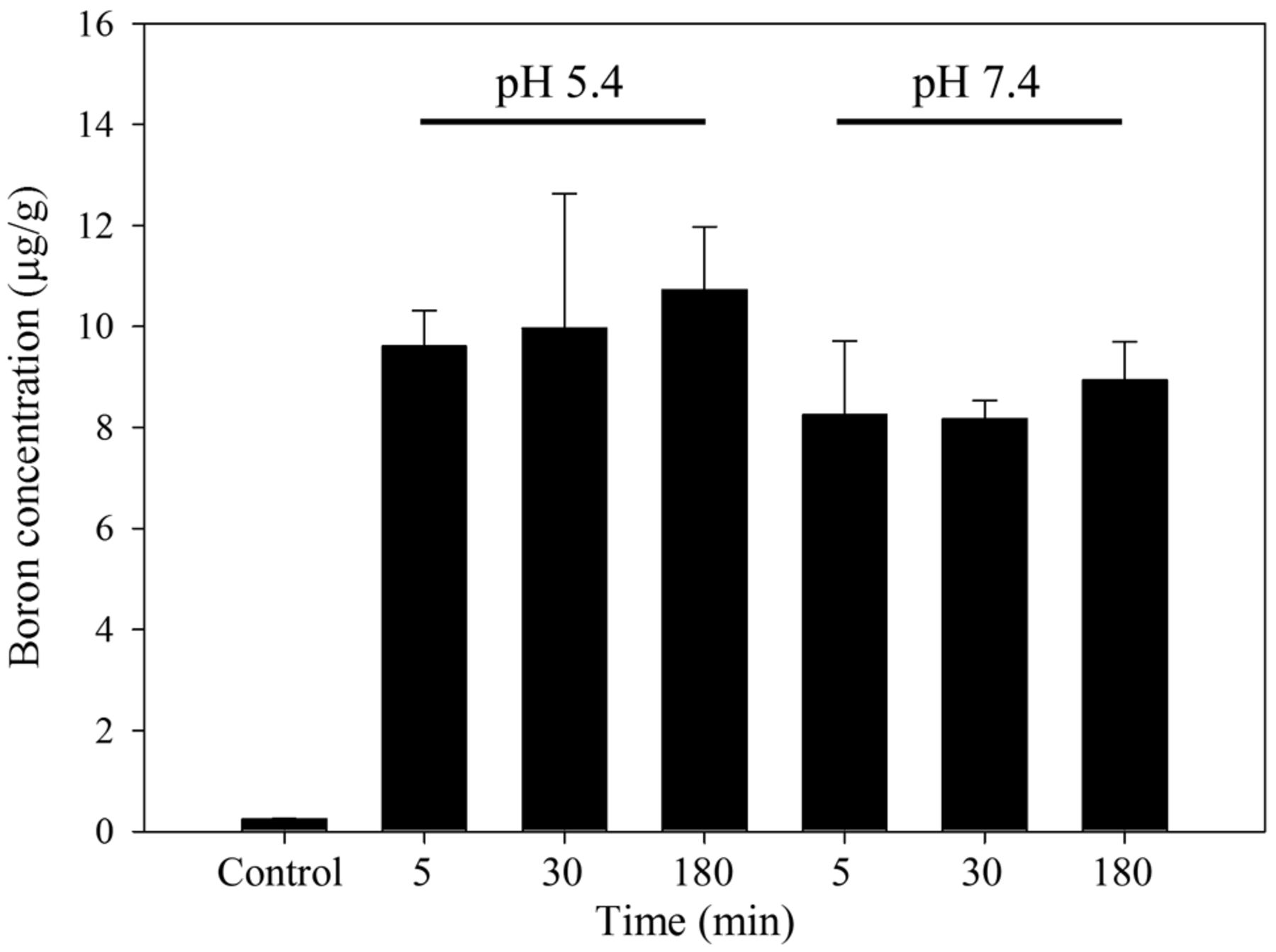
Boron uptake of HepG2 cells under different pH conditions. HepG2 cells were cultured for 48 h and then treated with 25 μg 10B/ml boric acid in culture medium which had been titrated to either pH 5.4 or pH 7.4 for 5, 30, or 180 min. Data are means±SD (n=3). Uptake did not differ significantly between treatments (p-values of 0.2208, 0.3109 and 0.1019 at 5, 30 and 180 min, respectively).

Boron uptake of HepG2 cells cultured with cholesterol. HepG2 cells were cultured for 47 h and treated with different concentrations of cholesterol for 1 h and then treated with 25 μg 10B/ml boric acid for 5 min (n=3). Data are means±SD. Uptake did not differ significantly between treatments (p-values of 0.0954, 0.2574, 0.2952 and 0.2001 for cells treated with 10, 100, 300 and 500 μg/ml cholesterol, respectively).
Discussion
In this study, we first postulated that glucose influx and glucose level might affect the uptake of BA in HepG2 cells, which could depend on the interaction between BA and glucose. However, we found no significant change in BA uptake under different glucose conditions (Figure 3).
The mechanisms of BA transport can be classified into simple diffusion and facilitated transport. Under simple diffusion, BA is directly transported through the cell membrane, with no carrier proteins involved. In this case, the direction of BA transport is determined by the gradient of BA inside and outside of the cell. On the other hand, the facilitated diffusion of BA has been extensively studied in plants. The transportation of BA in plants is assisted by specific carrier proteins, such as nodulin-26-like intrinsic protein NIP5;1, which is a well-known BA channel protein identified in plants (29).
Simple diffusion and facilitated diffusion are both concentration-dependent and bi-directional. In a classic rate of diffusion versus concentration plot, simple diffusion is represented by a linear line (Figure 2), which is the type of diffusion observed in this study. While BA is mainly transported via simple diffusion, we cannot rule-out that the possibility that the diffusion rate might be affected by other factors.
Based on the current understanding of glucose metabolism in HCC, glucose is quickly converted into pyruvate, which is then converted into lactate and acetyl-CoA to enable high aerobic glycolysis. As a result, the increased lactate level would acidify the tumor microenvironment, while acetyl-CoA acts to regulate fatty acid synthesis (30, 31). Based on this information, we investigated whether an acidic microenvironment could affect BA uptake in HepG2 cells. We found that an decrease in pH value might affect BA uptake by hepatoma cells to a small extent (Figure 4). Therefore, glucose metabolism might still play a role in affecting BA uptake in hepatoma cells due to its contribution to the acidification of the tumor microenvironment.
A microenvironment of lower pH results in higher cell membrane fluidity (26). This phenomenon occurs when the cancer cell membrane composition is modified by charged lipid head groups (32). The ability of the pH level to instantly affect cell membrane fluidity and permeability might explain why the uptake of BA would be accelerated at lower pH (Figure 4).
In addition, tumor blood vessels are known to be more acidic (pH 5.0-6.0) than the tumor microenvironment (pH 6.5-6.8) (25), which implies that the cell membrane fluidity of endothelial cells in tumor blood vessels might be higher than that of the surrounding cells. Consequently, this might explain why BA-treated liver tumor sections have a higher alpha-track signal density in tumor blood vessels (135,748 tracks/mm2) than in the tumor region (125,096 tracks/mm2) and in normal liver tissue (45,022 tracks/mm2) after neutron irradiation, which could lead to the selective destruction of tumor blood vessels as reported (15).
The pH could also regulate the balance of BA and borate levels, since the BA form exists at more than 95% when the pH value is lower than 7 (33). It should be noted that BA is a water-soluble chemical with a solubility of 4.72, 6.23 and 8.08 wt% at 20°C, 30°C and 40°C, respectively (34). Boric acid is a Lewis acid and usually exists as borate ions [i.e. B(OH)4−] in water with a pH value higher than 9 (33).
Considering the complex interactions of BA with other biomolecules via diol bonds, it is interesting to note that trans-diols do not react with BA at pH 5.4, while 10% of trans-diols can complex with BA at pH 7.4 (33). On the other hand, the formation of cis-diols decreases by 30% when the pH is dropped from 7.4 to 5.4 (33). This phenomenon might suggest that approximately 30% of BA is released from BA–diol complexes when they move into the acidic tumor microenvironment (pH 5.0-6.0). However, further experiments are needed to analyze the ratio of free BA to complexed BA in the blood.
In addition to pH value, the cholesterol level of hepatoma cells is decreased in HCC, especially for cases with advanced hepatomas (24). This altered lipid synthesis results in abnormal lipid composition in cancer cells, including a higher ratio of unsaturated to saturated fatty acids, and a lower ratio of cholesterol to phospholipid, both of which are important determinants of cell membrane fluidity (35). In this study, we found that decreased cell membrane fluidity by cholesterol treatment reduced BA uptake by HepG2 cells (Figure 5), which may partially contribute to the enhanced BA uptake seen in liver tumors compared to the normal liver tissues.
Liver tumors have disorganized, leaky vasculature and lack a lymphatic system. Specifically, tumor-derived vascular endothelial growth factor (VEGF) has been shown to enhance tumor angiogenesis, which not only promotes HCC progression but also increases tumor cell hyperpermeability (36). Since the concentration gradient of BA might affect the passive diffusion rate, it is plausible that BA accumulates in HCC because of the enhanced permeability of tumor blood vessels, which enable BA to enter tumor cells faster than normal neighboring liver tissues. The absence of a lymphatic system in liver tumors might reduce the clearance rate of BA in the tumor, restricting BA in the tumor interstitial fluid and causing selective accumulation of BA in the tumor.
In previous studies, BA has been shown to transiently accumulate in liver tumors in rat and rabbit studies. Our data further reveal that BA is transported into cancer cells by simple diffusion. This new understanding could be used to modify the current treatment protocol to increase efficacy and reduce adverse effects in patients, since BA is not metabolized in animals. Rapid transfusion of saline after BA-BNCT might accelerate the excretion of BA into urine (37).
In current BPA-BNCT clinical studies, patients with cancer undergo positron-emission tomography with 18F-labeled BPA for prognosis and treatment management (12). However, there is no comparable imaging technique for BA-BNCT. Given the roles of increased liver tumor vasculature and cell permeability in BA uptake and retention, we propose that dynamic contrast-enhanced magnetic resonance imaging (DCE-MRI) could be used as a potential medical imaging method for the prognosis and management of BA-BNCT treatment in HCC. During DCE-MRI measurements, contrast agent wash-in and extravasation changes the T1 relaxation time of tumor tissues (38). This can enable the computation of the volume transfer constant by associated signal change, which reflects the tumor vascular permeability and perfusion. We believe such properties could correlate positively to BA uptake in HCC. Therefore, DCE-MRI might have the potential to evaluate which patients with HCC are candidates for receiving BA-BNCT.
As our understanding of BA transportation and selective accumulation in tumor tissues improves, we hope that BA-BNCT will be able to significantly accelerate the development of better and safer HCC therapy.
Acknowledgements
This work was supported by the National Tsing Hua University preclinical BNCT boron drug development program; Grant No. 106N314CE1.
- Received August 16, 2017.
- Revision received September 8, 2017.
- Accepted September 14, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}