


Abstract
Background: A long noncoding RNA, p21-associated ncRNA DNA damage-activated (PANDA), associates with nuclear transcription factor Y subunit alpha (NF-YA) and inhibits its binding to promoters of apoptosis-related genes, thereby repressing apoptosis in normal human fibroblasts. Here, we show that PANDA is involved in regulating proliferation in the U2OS human osteosarcoma cell line. Materials and Methods: U2OS cells were transfected with siRNAs against PANDA 72 h later and they were subjected to reverse transcription-polymerase chain reaction (RT-PCR), quantitative RT-PCR and cell-cycle analysis. Results: PANDA was highly expressed in U2OS cells, and its expression was induced by DNA damage. Silencing PANDA caused arrest at the G1 phase of the cell cycle, leading to inhibition of cell proliferation. Quantitative RT-PCR showed that silencing PANDA increased mRNA levels of the cyclin-dependent kinase inhibitor p18, which caused G1 phase arrest. Conclusion: These results suggest that PANDA promotes G1-S transition by repressing p18 transcription, and thus promotes U2OS cell proliferation.
A long noncoding RNA (lncRNA), namely p21-associated ncRNA DNA damage-activated (PANDA), has been identified by ultra-high-density tiling array as a lncRNA transcribed from the nearby cyclin-dependent kinase inhibitor 1A (CDKN1A) locus (1). The PANDA gene is located approximately 5 kb upstream of the CDKN1A locus and is transcribed in the opposite direction to CDKN1A. There is a p53-binding site between PANDA and the CDKN1A locus (2). Although both genes are induced by p53 in response to DNA damage, their products have opposing roles in regulating cell proliferation. Whereas p21 protein, the product of CDKN1A, functions as a CDK inhibitor and causes cell-cycle arrest at the G1 phase (3), PANDA promotes cell survival (1). Recently, we reported that PANDA contributes to stabilization of the p53 protein, indicating that PANDA and p53 form a positive feedback loop (4).
It has been reported that several lncRNA, such as X inactive specific transcript (XIST) (5), antisense IGF2R RNA (AIR) (6), KCNQ1 overlapping transcript 1 (KCNQ1OT1) (7, 8) and antisense non-coding RNA in the INK4 locus (ANRIL) (9-12), are involved in transcriptional regulation of target genes. Hung et al. showed that PANDA associates with nuclear transcription factor Y subunit alpha (NF-YA), a component of the nuclear transcription factor NF-Y (1). NF-Y directly binds to and activates the promoter of the pro-apoptotic gene, FAS cell surface death receptor (FAS) (13). Most lncRNAs involved in transcriptional regulation function as recruiters for transcriptional regulators such as polycomb proteins (14, 15). However, PANDA does not function as a recruiter but as a repressor of NF-YA. PANDA induced by DNA damage signalling binds to NF-YA and inhibits its binding to the promoters of apoptosis activators including apoptotic peptidase activating factor 1 (APAF1), B-celll/lymphoma 2-interacting killer (BIK), FAS and leucine-rich repeat and death domain-containing protein (LRDD), thereby repressing their transcription (1). This results in repression of apoptosis in response to DNA damage in human normal fibroblasts.
Recently, we reported that PANDA is induced by enforced expression of an oncogenic form of the small GTPase rat sarcoma viral oncogene homolog (RAS) (known as oncogenic RAS) (16). Oncogenic RAS signalling protects some cell strains from apoptosis (17), leading to progression of carcinogenesis. Thus, oncogenic RAS-induced PANDA expression might be involved in carcinogenesis via repression of apoptosis. In this study, we investigated the role of PANDA in the proliferation of cancer cells, namely human osteosarcoma U2OS cells.
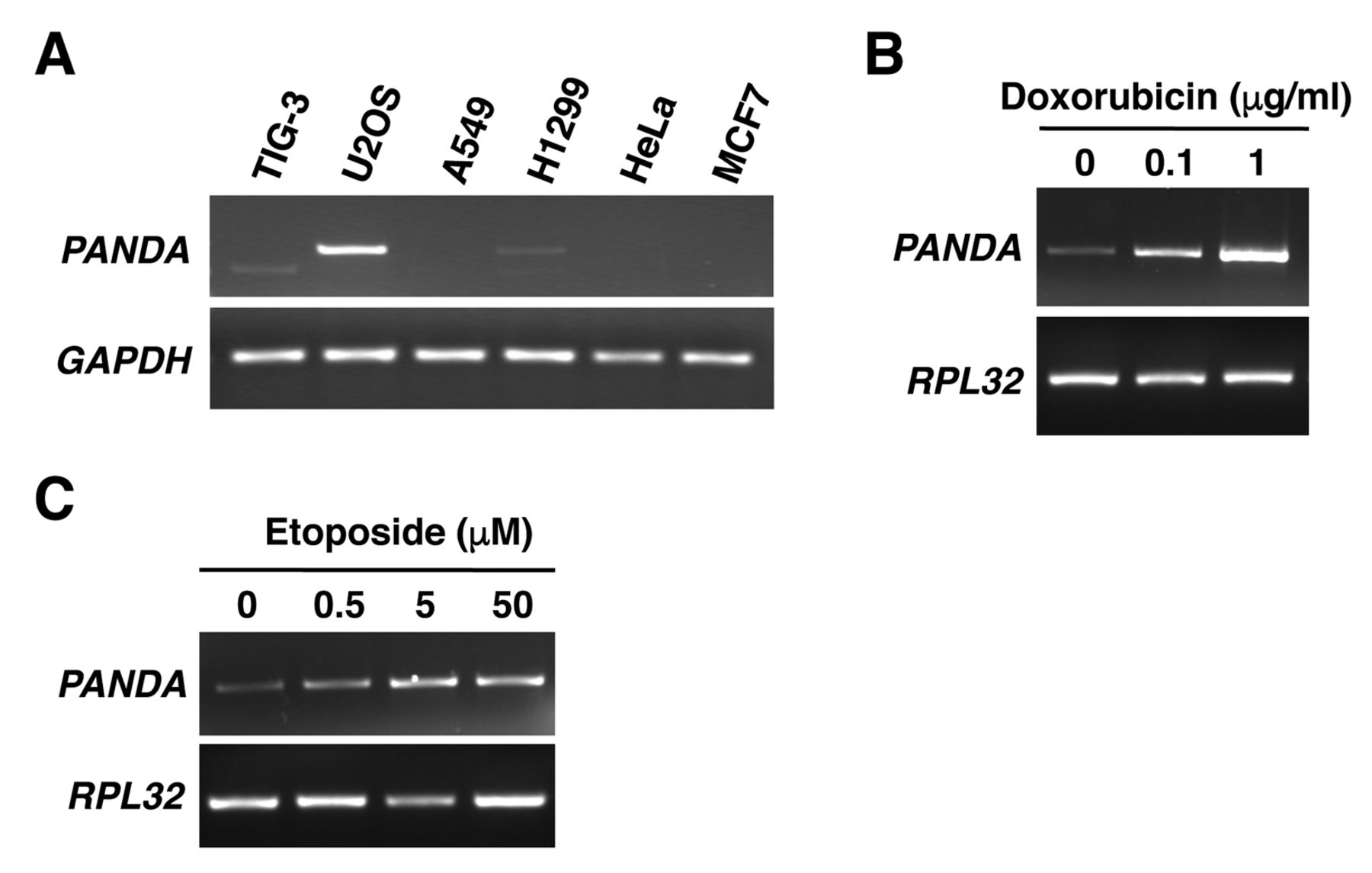
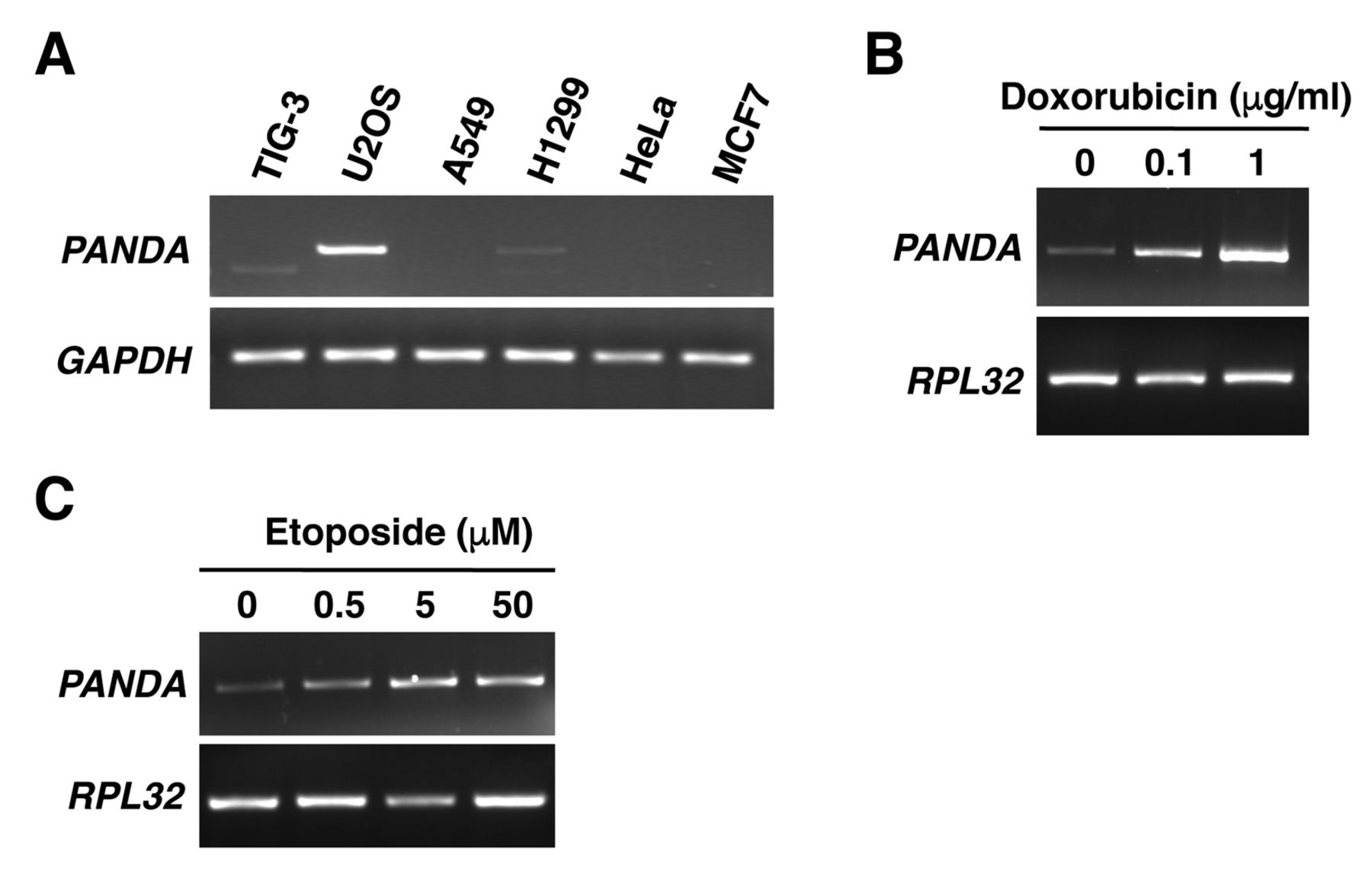
p21-Associated ncRNA DNA damage-activated (PANDA) is highly expressed in U2OS cells and its expression is induced by treatment with doxorubicin and etoposide. A: PANDA expression was detected by reverse transcription-polymerase chain reaction (RT-PCR). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control. U2OS cells were incubated with or without doxorubicin (B) or etoposide (C) for 24 h then harvested. PANDA expression was detected by RT-PCR. Ribosomal protein L32 (RPL32) was used as an internal control.
Materials and Methods
Cell culture and RNA interference. Human normal fibroblasts TIG-3, osteosarcoma cells U2OS, lung cancer cells A549, H1299, cervical cancer cells HeLa, and breast cancer cells MCF7 were cultured in Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (GIBCO, Grand Island, NY, USA) in an incubator at 37°C with 5% CO2. Viable and dead cells were counted by trypan blue staining. The cells were treated with doxorubicin (SIGMA-ALDRICH, Tokyo, Japan) at 0.1 or 1 μg/ml, etoposide (Wako, Osaka, Japan) at 0.5, 5 or 50 μM or vehicle (dimethyl sulfoxide; SIGMA-ALDRICH) for 38 h. U2OS cells were transfected with siRNA oligonucleotides to silence human PANDA using Lipofectamine RNAiMAX (Invitrogen). The nucleotide sequence of the PANDA siRNA oligonucleotide was 5’-AAUGUGUGCACGUAACAGAUU-3’.
Reverse transcription-polymerase chain reaction (RT-PCR) and quantitative RT-PCR (qRT-PCR). RT-PCR and qRT-PCR assays were performed as previously described (11). The sequence of the PCR primers for p15, p16, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and ribosomal protein L32 (RPL32) are described in (11). The sequence of primers for PANDA, p18, p19, p21, p27 and p57 were as follows: PANDA: 5’-AGACCCCAGTGGCACCTGAC-3’ and 5’-GGGCAGAACTTGGCATGATG-3’; p18: 5’-CGTCAATGCACAAAATGGATTTGG-3’ and 5’-GAATGACAGCGAAACCAGTTCGG-3’; p19: 5’-GTGCATCCCGACGCCCTCAAC-3’ and 5’-TGGCACCTTGCTTCAGCAGCTC-3’; p21: 5’-GGCAGACCAGCATGACAGATT-3’ and 5’-GCGGATTAGGGCTTCCTCTT-3’; p27: 5’-ATAAGGAAGCGACCTGCAACCG-3’ and 5’-TTCTTGGGCGTCTGCTCCACAG-3’; p57: 5’-AGATCAGCGCCTGAGAAGTCGT-3’ and 5’-TCGGGGCTCTTTGGGCTCTAAA-3’. qPCR assays were performed using an Mx3000P Real-Time Q-PCR System (Agilent Technologies, Santa Clara, CA, USA). The quantification of data was performed by relative quantitative determination method using calibration curve. Mean values and standard deviations were calculated from triplicates of three independent experiments. The statistical analyses were performed using the paired one-tailed t-test.
Cell-cycle analysis. A total of 1×106 U2OS cells transfected with control siRNA or siRNA oligonucleotides targeting PANDA were fixed with 70% ethanol at −20°C overnight. The fixed cells were then subjected to staining using a Muse Cell Cycle Kit (Merck Millipore, Darmstadt, Germany) according to the manufacturer's protocol. They were then analysed with a Muse Cell Analyzer (Merck Millipore).
Results
We first examined the expression pattern of PANDA in various human cell lines. RT-PCR data showed that PANDA was highly expressed in the human osteosarcoma cell line U2OS, compared with in other cell lines (Figure 1A). It has been reported that PANDA expression was increased in normal human fibroblasts by treatment with the anticancer drug doxorubicin, which causes DNA damage (1). We, therefore, examined whether PANDA expression was also induced by DNA damage in U2OS cells. RT-PCR data showed that treatment with doxorubicin (Figure 1B), and with etoposide (Figure 1C), another DNA-damaging drug, increased PANDA expression. These results indicate that PANDA is indeed induced in response to DNA damage in U2OS cells.
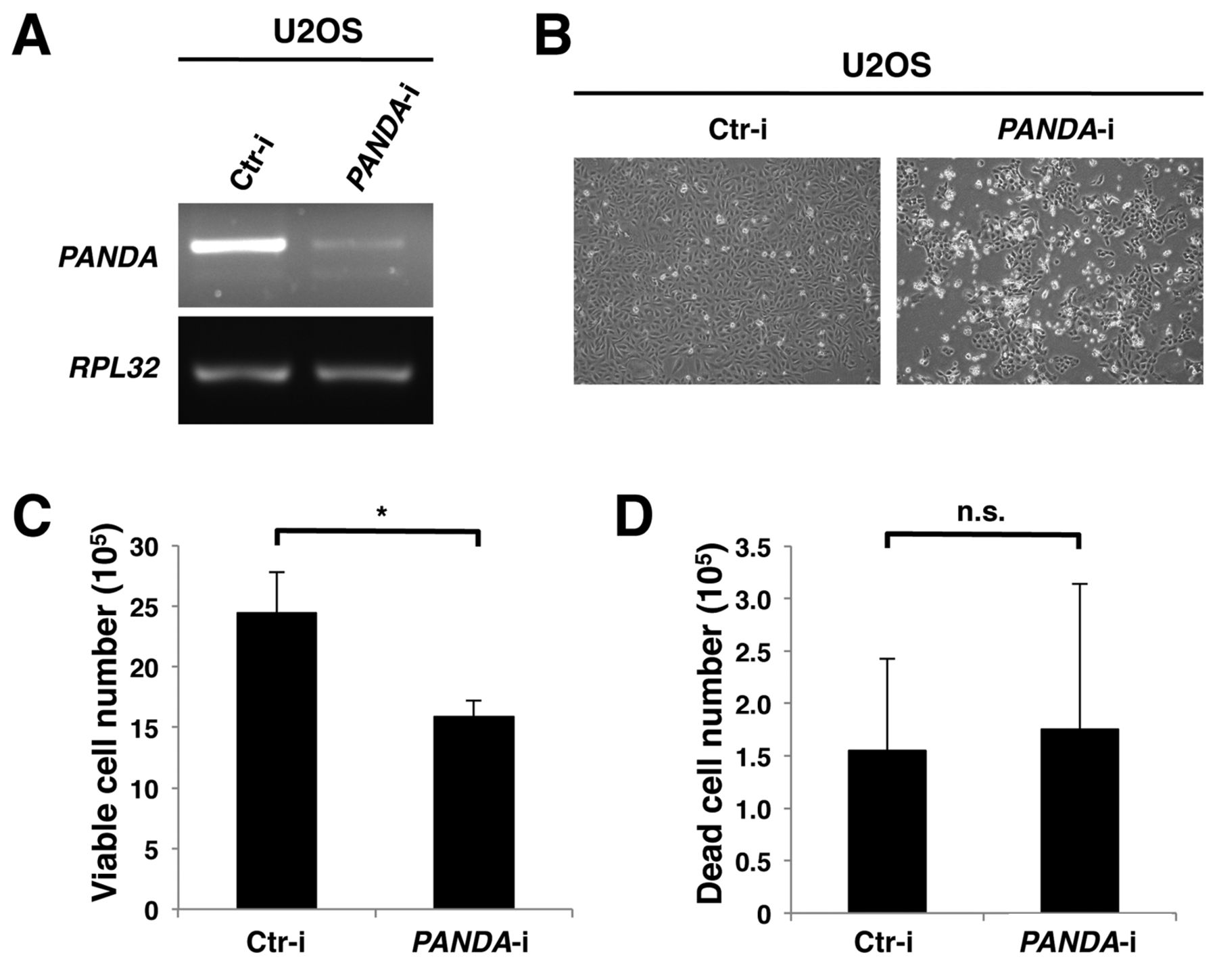
Silencing p21-associated ncRNA DNA damage-activated (PANDA) reduces the number of viable cells but does not affect the number of dead cells. A: U2OS cells were transfected with control siRNA (Ctr-i) or siRNA oligonucleotides targeting PANDA (PANDA-i). At 72 h after transfection, cells were harvested and subjected to reverse transcription-polymerase chain reaction. B: Phase-contrast microscopic images of cells transfected with control or PANDA siRNA oligonucleotides. Viable (C) and dead (D) cells were counted by trypan blue staining at 72 h after transfection. Data were analyzed using the Student's t-test. *p<0.05; n.s., not significant.
We next examined the function of PANDA in U2OS cells. We depleted PANDA expression by transfecting siRNA oligonucleotides targeting PANDA into U2OS cells. The siRNA oligonucleotides reduced the PANDA level such that it was almost undetectable (Figure 2A). We investigated the effects of PANDA depletion on U2OS cell proliferation. Silencing PANDA also resulted in a decrease in the number of viable cells (Figure 2C), but did not affect the number of dead cells (Figure 2D). We further investigated the effects of PANDA depletion on the cell cycle in U2OS cells. Cell-cycle analysis revealed that silencing PANDA resulted in an increase in the proportion of cells in the G1 phase and a decrease of the proportion of cells in the S and G2/M phases (Figure 3). These results suggest that PANDA positively regulates cell proliferation by promoting G1 phase progression.
Given that silencing PANDA causes cell-cycle arrest at the G1 phase, we next examined the effects of PANDA depletion on the expression of all CDK inhibitors that cause G1 phase arrest by inhibiting CDK enzymatic activity (3). qRT-PCR data showed that silencing PANDA significantly increased the mRNA level of p18, but did not increase those of other CDK inhibitors (Figure 4). Expression of p16 was not detected in U2OS cells, because the p16 promoter is methylated and transcriptionally repressed in U2OS cells. These data suggest that PANDA is involved in the repression of p18 transcription.
Discussion
In this study, we revealed that PANDA is involved in the positive regulation of proliferation in U2OS cells via regulation of the G1 phase of the cell cycle. We observed that PANDA was highly expressed in U2OS cells compared to other human cell lines, and its expression was induced in response to DNA damage. Hung et al. reported that PANDA expression was induced in a p53-dependent manner in normal human fibroblasts (1), and showed that silencing PANDA enhanced DNA damage-induced apoptosis. In this study, we showed that in the absence of stress from DNA damage, silencing PANDA reduced the number of viable cells, but did not affect the number of dead cells. We, furthermore, showed that silencing PANDA caused cell cycle arrest at the G1 phase. Taken together, these results suggest that in the absence of stress from DNA damage, PANDA may positively regulate cell proliferation by promoting G1 phase progression, and that once cells undergo DNA damage, PANDA expression increases, which may function to repress apoptosis and thus promote cell survival.

Silencing p21-associated ncRNA DNA damage-activated (PANDA) results in cell-cycle arrest at the G1 phase in U2OS cells. A: U2OS cells were transfected with control siRNA (Ctr-i) or siRNA oligonucleotides targeting PANDA (PANDA-i). At 72 h after transfection, the cells were fixed overnight with 70% ethanol. After propidium iodide staining, the fixed cells were subjected to cell-cycle analysis. The x axes indicate PI staining intensity, reflecting the DNA index, and the y axes indicate cell counts. B: The proportions of cells in the G1, S and G2/M phases of the cell cycle are shown.
We also showed that silencing PANDA increased the expression level of p18 mRNA, suggesting that PANDA represses p18 transcription. The molecular mechanism by which PANDA represses p18 expression is undetermined. It has been reported that PANDA binds to NF-YA, a transcription factor, and inhibits its binding to the promoters of target genes such as FAS, phorbol-12-myristate-13-acetate-induced protein 1 (PMAIP1) and BCL2-binding component 3 (BBC3), leading to the repression of their transcription (1). However, silencing NF-YA did not reduce the level of p18 mRNA (data not shown), suggesting that NF-YA is not involved in the regulation of p18 transcription. It is, therefore, an important issue to elucidate the NF-YA-independent mechanism of p18 repression by PANDA.

Effects of p21-associated ncRNA DNA damage-activated (PANDA) silencing on the expression of cyclin-dependent kinase (CDK) inhibitors in U2OS cells. U2OS cells were transfected with control siRNA (Ctr-i) or siRNA oligonucleotides targeting PANDA (PANDA-i) and harvested at 72 h after transfection. The mRNA levels of the CDK inhibitors p15, p18, p19, p21, p27 and p57 were determined by quantitative reverse transcription-polymerase chain reaction. Results are expressed relative to the corresponding values in U2OS cells transfected with control siRNA oligonucleotides (Ctr-i). The mean values and standard deviations were calculated from three representative experiments. Data were analyzed using the paired t-test. *p<0.05; n.s., not significant.
Hung et al. showed that PANDA is highly expressed in metastatic ductal carcinomas compared to normal breast tissue (1). We recently reported that PANDA expression is increased by oncogenic RAS signaling (16). We, therefore, postulate that aberrant expression of PANDA promotes cell proliferation by repressing p18, and represses apoptosis, leading to oncogenic transformation.
Acknowledgements
We thank Chihiro Murasaki and Masashige Hirata for their technical assistance and our laboratory members for helpful discussion. This work was supported in part by grants from the Ministry of Education, Culture, Sports, Science and Technology of Japan 26430127 (to Y. Kotake) and the Takeda Science Foundation (to Y. Kotake).
Footnotes
This article is freely accessible online.
- Received October 31, 2016.
- Revision received December 6, 2016.
- Accepted December 7, 2016.
- Copyright© 2017 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}