


Abstract
Background: Cancer stem cells (CSCs) have a high tumorigenic ability to form patient-derived tumor xenografts (PDXs). PDXs are an attractive pre-clinical model, but gene expression and biological behavior of cancer cells in the tumor will change during establishment and passage of PDXs. Materials and Methods: Human colon cancer PDX was established and passaged either subcutaneously or orthotopically into the murine intestine. Histology and flow cytometric profile of the surgical specimen and the PDX were analyzed. CSCs were then isolated from the tumors and their microRNA (miRNA) expression was analyzed by semi-quantitative polymerase chain reaction. Results: The surgical specimens and PDXs were histologically similar. The size of CSC population increased and expression of miRNAs in CSCs changed in the passaged PDXs. Expression of oncogenic miRNAs was highly up-regulated in the CSCs of the orthotopically passaged PDXs. Conclusion: The xenotransplantation site and the number of tumor passages affect the miRNA expression of human colon CSCs.
Patient-derived tumor xenografts (PDXs) established by transplantation of surgically resected human cancer specimens into immunodeficient mice are able to mirror patients' histopathological and genetic profiles (1), and have been applied to pre-clinical drug testing and biomarker identification (2, 3). Furthermore, the successful tumor engraftment and metastatic progression in the PDX mouse models depend on cancer stem cells (CSCs) within the tumor tissues (4).
MicroRNAs (miRNAs) are non-coding RNAs with fewer than 25 nucleotides and function as regulators of gene expression and signaling pathways that are important for normal physiological functions and disease pathogenesis. miRNAs are differentially expressed between CSCs and other non-tumorigenic cancer cells (NTCs) within the same tumor (4-6). The miRNAs specifically expressed in CSCs, such as miR-200 family miRNAs and miR-142, target signaling pathways critical for the regulation of CSCs.
The xenotransplantation site and number of tumor passages influence the gene-expression profiles, engraftment rate and biological behavior of PDXs (7, 8). Distant metastases from the transplanted site are more frequently observed when the colon PDX tumors are transplanted orthotopically into the intestine rather than being transplanted subcutaneously (9, 10). In lung cancer, 17% of miRNAs were differentially expressed between primary lung tumors and established PDX tumors (11). However, the effect of xenotransplantation site and number of tumor passages on gene expression of the CSCs, the most tumorigenic population of cancer cells, have not been elucidated. In this study, we investigated the miRNA expression of CSCs and NTCs in colon cancer obtained from a patient and the corresponding PDX tumors passaged subcutaneously or orthotopically into murine intestine.
Materials and Methods
Ethics statements. Primary human colon cancer tissue was obtained from a consenting patient at the Kobe University Hospital as approved by the Institutional Review Boards at Kobe University (approval number: 1299). The cancer type and histological grade were obtained from clinical and histopathological reports. All animal experiments were performed under the approval of the Kobe University Animal Care and Use Committee. All surgical procedures were performed under isoflurane anesthesia with care to minimize suffering of mice.
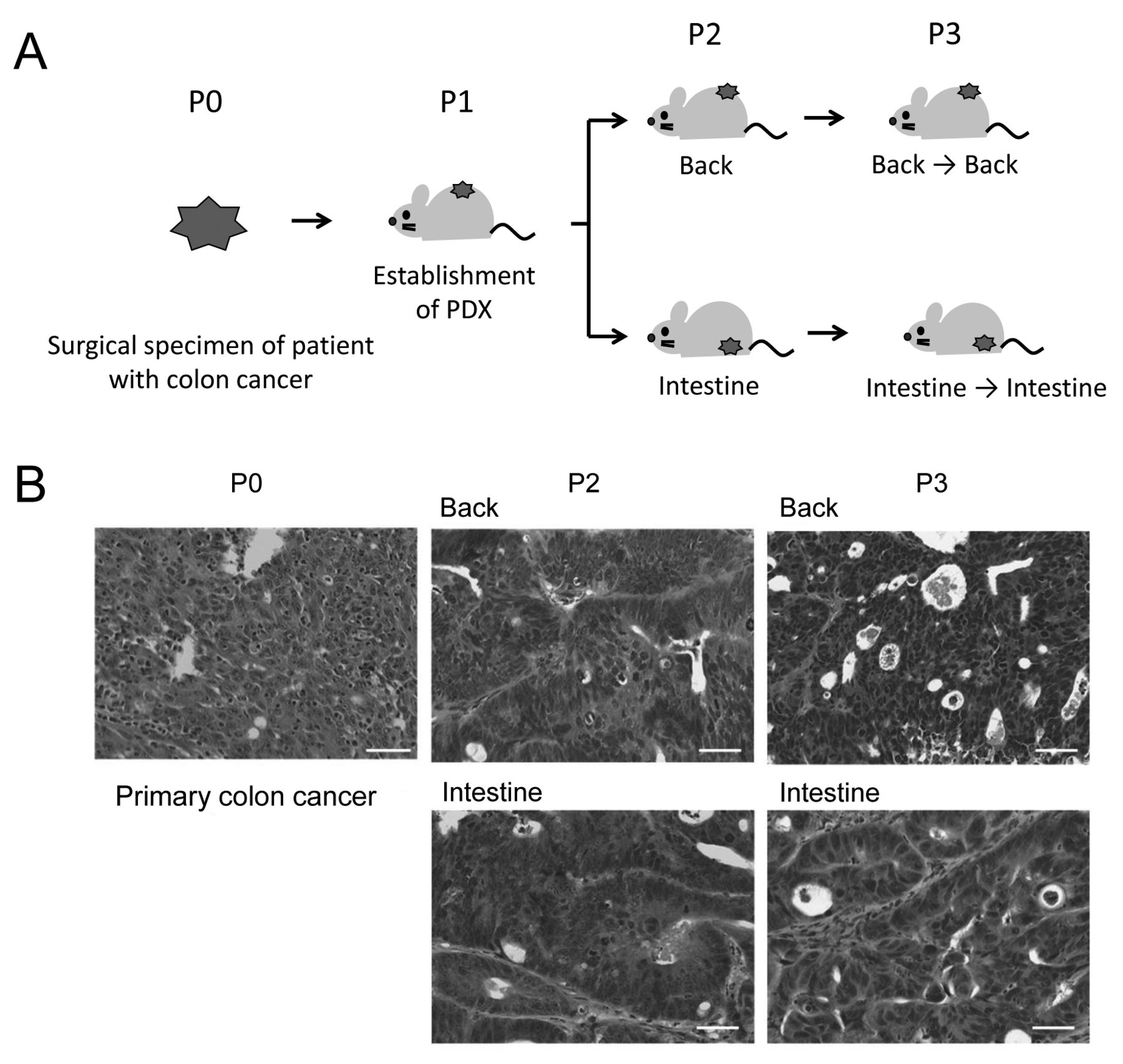
Establishment and passage of the human colon cancer patient-derived tumor xenografts (PDXs). A: Schematic illustration of the study design. Human colon cancer PDX was established by subcutaneous transplantation of the surgical specimen of a patient with colon cancer into the immunodeficient mice. The established PDX was then passaged either subcutaneously or orthotopically into the intestine. Back: Subcutaneous xenotransplantation; Intestine: orthotopic transplantation into the intestine, P0-3: number of tumor passages. B: Histological appearance of the patient colon cancer and PDX tumors. The PDX tumors retained the histological appearance of the patient cancer tissue, and were similar between the subcutaneously and the orthotopically passaged PDXs. Representative HE sections are presented. Bars: 50 μm.
PDX. The surgical specimen obtained from the patient with colon cancer was minced, suspended in Matrigel (BD Biosciences, San Jose, CA, USA), and then subcutaneously transplanted on the back of a female non-obese diabetic/severe combined immunodeficiency/Il2rgnull (NSG) mouse (Oriental Kobo, Japan) as previously described (5, 12) (Figure 1A). The established PDXs were passaged either subcutaneously on the back or orthotopically into the intestine of the mice. When the tumors reached approximately 1-2 cm in size, they were harvested for further analyses.
Histopathological evaluation. A portion of the patient colon cancer specimen and PDX tumors were fixed in 4% paraformaldehyde/phosphate-buffered saline (PBS) for 12 hours at 4°C. After paraffin-embedding of specimens, they were cut and stained with hematoxylin and eosin (H.E.).
Flow cytometric analysis. The patient cancer specimen and PDX tumors were dissociated and analyzed as previously described (13). Briefly, the tumor tissues were minced into small pieces with a razor blade, resuspended in RPMI-1640 medium with 10% fetal bovine serum, 2 mM glutamine, 120 μg/ml penicillin, 100 μg/ml streptomycin, 20 mM HEPES, 1 mM sodium pyruvate, 100 units/ml DNase-I (Sigma, Saint Louis, MO, USA) and 200 units/ml Collagenase-III (Worthington, Lakewood, NJ, USA), and incubated at 37°C for 2 h. The cells were then filtered through a 40-μm strainer and were blocked with normal mouse IgG (1:100; Wako, Osaka, Japan). Then the cells were stained with anti-mouse MHC class I H2kD (1:40; eBioscience, San Diego, CA, USA), fluorescein isothiocyanate (FITC)-conjugated anti-epithelial cell adhesion molecule (EpCAM) (1:10; Biolegend, San Diego, CA, USA), and allophycocyanin (APC)-conjugated anti-human cluster of differentiation (CD) 44 (1:20; BD Biosciences, San Jose, CA, USA) antibodies. The cells were analyzed and collected using FACS Aria III (BD Biosciences).

Increase of the size of a cancer stem cell (CSC) population. The profile of the cancer cells within the patient colon cancer (P0) and patient-derived tumor xenograft tumors were analyzed using flow cytometry. The percentage of the cells within the CSC population [epithelial cell adhesion molecule (EpCAM)+/cluster of differentiation (CD) 44+lineage−/ 4’,6-diamidino-2-phenylindole (DAPI)−]of the patient colon cancer and orthotopically passaged DX tumors increased with serial transplantation. CD44+/EpCAM+ CSCs (solid line) and CD44−/EpCAM+ non-tumorigenic cancer cells (NTCs) (dashed line) were collected for miRNA expression analyses.
Semi-quantitative real-time polymerase chain reaction. Reverse transcription, pre-polymerase chain reaction and real-time polymerase chain reaction for the expression analyses of five miRNAs (miR-25, miR-210, miR-221, miR-203 and miR-423) were performed using the RNA extracted from the sorted cells of the patient carcinoma and PDX tissues as templates (5, 14). The abundance of each miRNA was measured individually using a Thermal Cycler Dice (TaKaRa, Shiga, Japan). Results were normalized by the amount of small nuclear RNA expression, SNORD48, or U6 snRNA.
Results
Similarity of the histological appearance of the patient cancer specimen and PDXs. The PDX is an attractive model for analyzing CSCs, a highly tumorigenic population of cancer cells, by enabling recapitulation of the cellular heterogeneity of patient cancer tissues. Patient specimens of primary colon cancer can be engrafted subcutaneously or orthotopically into the intestine of the immunodeficient mice. To investigate the effect of transplantation site and tumor passage on molecular characteristics of CSCs and NTCs, we first engrafted patient colon cancer tissue subcutaneously into the NSG mice, and then the established PDX was passaged either subcutaneously or orthotopically in the intestine (Figure 1A). Histologically, the patient cancer specimen was moderately differentiated tubular adenocarcinoma (Figure 1B). The cancer tissue was mostly filled with cancer cells, a proportion of which exhibited a glandular structure, and contained a few stromal tissues. Histological examination showed that the established PDX tumors we analyzed reflected histopathological appearances of the patient colon cancer tissue well, irrespective of the transplantation site and the number of tumor passages (Figure 1B).
Increase of the CSC population by tumor passaging. The flow cytometric profile of the patient colon cancer and the PDX tumors were analyzed. CD44 is an established marker used to isolate CSCs from primary colon cancer specimens (15). The population of CD44+ CSCs within the EpCAM+ colon cancer epithelial cells was 0.8% in this patient [at passage 0 (P0)] and was increased by tumor passaging: 3.2% in the orthotopically P2 PDX and 32% in the P3 PDX (Figure 2). The increase of CD44+ CSCs was also observed in the subcutaneously passaged P2 and P3 PDXs. These results suggest that although the histological appearance was similar in the patient cancer and PDXs, heterogeneity of cancer cells within the cancer tissues changed on tumor passaging.
Change in miRNA expression after tumor engraftment. Because miRNAs are important regulators of CSC and NTC functions, we analyzed the miRNA expression of CSCs and NTCs collected from the patient tumor and PDXs established and passaged subcutaneously or orthotopically. We analyzed the expression of five miRNAs that function as oncogenes in colon cancer (miR-25, miR-210 and miR-221) and as regulators of stem cell functions (miR-203 and miR-423) (16-20). We compared the expression levels of miRNAs between the patient tumor (P0) and the engrafted tumor (P1) (Figures 3 and 4). The expression levels of miR-203 and miR-210 were higher in CSCs of P1 PDX than P0 CSCs, and those of miR-25, miR-221 and miR-423 were similar between P1 and P0 CSCs. However, the expression levels of all five miRNAs were much higher in NTCs of P1 PDX than P0 NTCs. These results suggest that the expression of some miRNAs is modulated by tumor engraftment and the effects are much larger in NTCs than CSCs.

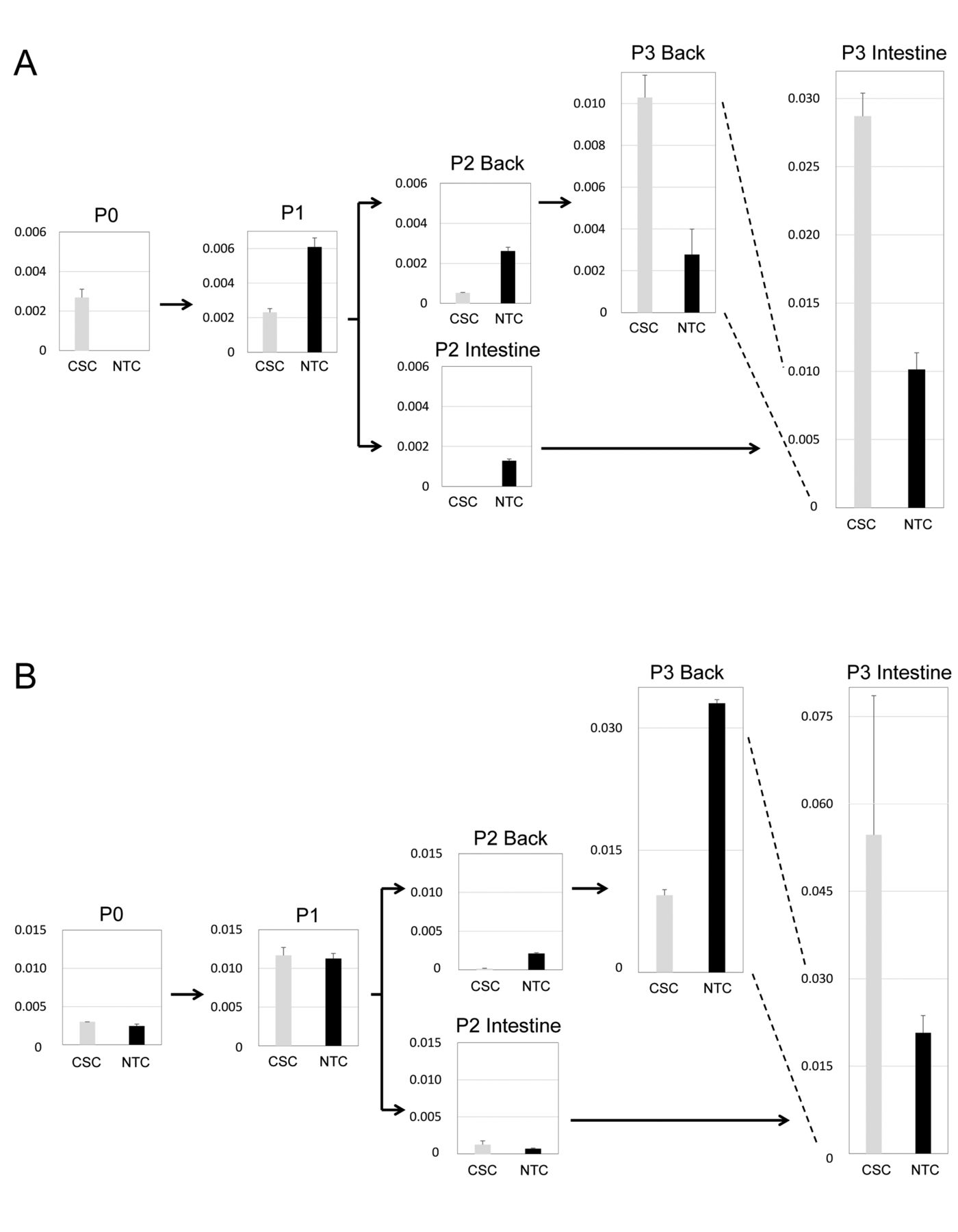
Change of miRNA expression due to xenotransplantation. The miRNA expression profile of the cancer stem cells (CSCs) and non-tumorigenic cancer cells (NTCs) from the patient colon cancer and PDX tumors were measured using semi-quantitative real-time polymerase chain reaction. The intensity of miRNA expression in CSCs and NTCs were significantly affected by the establishment of PDX [passage (P) 0 vs. P1], and further affected by serial transplantation. The results for two representative miRNAs, miR-203 (A) and miR-423 (B), are presented. The data are the mean+S.D. (n=2).
Up-regulation of oncogenic miRAs in the CSCs of orthotopically passaged PDXs. Because the transplantation site affects the tumorigenicity and formation of distant metastases in colon cancer PDXs (9,10), we analyzed the miRNA expression levels of CSCs of orthotopically and of subcutaneously passaged PDXs. The expression level of miR-203 was higher in the CSCs, but was unchanged in the NTCs of the orthotopically passaged PDXs (Figure 3A, intestine). The expression level of miR-423 was low in the CSCs, but was variable in the NTCs of the orthotopically passaged PDXs (Figure 3B, intestine). The expression level of miR-25 was significantly higher in the passaged P3 tumors, especially in the CSCs of orthotopically passaged P3 PDX (Figure 4A). The expression levels of miR-210 and miR-221 were significantly higher in the CSCs of orthotopically passaged P3 PDX (Figure 4B and C, intestine). It is of note that miR-25, miR-210 and miR-221 function as oncogenic miRNAs in gastrointestinal cancer (17, 21). These results suggest higher tumorigenicity of the orthotopically transplanted PDX tumor will be caused by the significant up-regulation of oncogenic miRNAs in the CSCs.

Increase of the expression of oncogenic miRNAs by serial transplantation. The miRNA expression profile of the cancer stem cells (CSCs) and non-tumorigenic cancer cells (NTCs) from the patient colon cancer and patient-derived tumor xenograft (PDXs) tumor were measured using semi-quantitative real-time polymerase chain reaction. The intensity of expression of oncogenic miRNAs miR-25 (A), miR-210 (B) and miR-221 (C) were significantly increased by serial transplantation. The increase was extremely higher in the P3 PDX orthotopically passaged into the intestine. The data are the mean+S.D. (n=2).
Discussion
In this study, we focused on the miRNA expression of CSCs and NTCs isolated from a patient cancer specimen and PDXs. Because CSCs are a minority population of cancer cells in human colon cancer tissues (Figure 2) (15), specific characteristics of CSCs are difficult to reveal without actually isolating them. We compared the miRNA expression of CSCs and NTCs between the surgical specimen of colon cancer and the initially engrafted tumor (P1 PDX). The difference in the expression level of miRNAs, such as miR-203, miR-25 and miR-221 between the patient cancer and P1-PDX was much larger in NTCs than in CSCs. Thus, our results suggest the possibility that CSCs are more resistant than NTCs to the change of microenvironment after initial engraftment (22). Therefore, it is possible that the global change of the expression level of at least some miRNAs does not reflect that of CSCs.
Our findings allow us to propose a molecular mechanism for the enhancement of the aggressive behavior of the PDXs transplanted orthotopically into the intestine compared with those transplanted subcutaneously (9, 10). The expression levels of oncogenic miRNAs, such as miR-25, miR-210 and miR-221, were extremely elevated in the CSCs of the orthotopically passaged PDXs, but not in the CSCs of the subcutaneously passaged PDXs. Moreover, the elevation of these miRNAs was more evident in the CSCs than in the NTCs. It is reported that dynamic clonal change occurs at initial engraftment and during serial passaging of human breast cancer PDXs (23). Thus, it is highly possible that dominant CSC clones after serial passaging were different between the orthotopically passaged PDXs and the subcutaneous PDXs. In addition, human stromal components in PDXs are replaced by murine equivalents through serial passaging (24). Among the stromal components, cancer-associated fibroblasts play an important role in cancer progression and modulate the tumor microenvironment by communicating with tumor and other stromal cells (25, 26). In a recent study, it became clearer that the characters of fibroblasts differs depending on their anatomical localization (27). Therefore, the difference of the murine microenvironment, such as cancer-associated fibroblasts, endothelial cells and cytokines, between orthotopically passaged PDXs and subcutaneously passaged PDXs, may have affected the miRNA expression levels in the CSCs of the PDXs.
Considering that CSCs have extremely high tumorigenic capability and are responsible for metastatic progression, analyses of the miRNA expression in CSCs will further clarify the gene-expression changes that need to be considered when using the PDXs as pre-clinical models.
Acknowledgements
This study was supported by Grants-in-Aid for Scientific Research from the Japan Society of the Promotion of Science and by the grants from the Japan Foundation for Applied Enzymology and the Ithoh-Chubei Foundation.
- Received April 5, 2016.
- Revision received May 20, 2016.
- Accepted May 24, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}