


Abstract
Aim: To investigate the effects of vector-based small interfering RNA (siRNA) targeting MDR1 on the reversal of multidrug resistance in a mouse model of doxorubicin (DOX)-resistant human hepatocellular carcinoma. Materials and Methods: Three siRNAs plasmid vectors (MDR1 siRNA1, MDR1 siRNA2 and MDR1 siRNA3) targeting MDR1 were constructed and transfected into DOX-resistant human hepatocellular carcinoma Bel-7402/ADM cells. The expression of MDR1 mRNA and P-glycoprotein (P-gp) was detected with RT-PCR and western blotting, respectively. A nude mouse model of DOX-resistance was established with untransfected Bel-7402/ADM or Bel-7402/ADM transfected with MDR1 siRNA (Bel-7402/ADMsi). The nude mice with tumors from untransfected Bel-7402/ADM cells were treated with either saline (Group 1); intravenous DOX (Group 2); or the combination of intra-tumoral MDR1 siRNA and intravenous DOX (Group 3). The nude mice with tumors from Bel-7402/ADMsi cells were treated with intravenous DOX (Group 4). DOX and MDR1 siRNA were administered twice a week at 20 mg/kg/dose and 9.8 mg/kg/dose, respectively. Tumor growth was measured to assess reversal of multidrug resistance by MDR1 siRNA. Results: MDRl mRNA and P-gp expression of Bel-7402/ADM cells was reduced by transfection of three siRNAs with different silencing efficiency (p<0.05). DOX treatment (Group 4) resulted in significant reduction in tumor size in the Bel-7402/ADMsi tumor model (p<0.05), indicating reversal of multidrug resistance in tumor by MDR1 siRNA. However, the combination treatment of intratumoral MDR1 siRNA and DOX (Group 3) showed no significant anti-tumor efficacy in the untransfected Bel-7402/ADM (p>0.05) tumor model, suggesting poor in vivo transfection efficiency of MDR1 siRNA. Analysis of the tumor samples showed the reduced expression level of MDR1 mRNA and P-gp was due to efficacy of MDR1 siRNA. Conclusion: In vitro transfection of siRNAs' vectors targeting the MDR1 gene can effectively silence MDR1 mRNA and P-gp expression in DOX-resistant human hepatocellular carcinoma Bel-7402/ADM cells that resulted in reversal of multidrug resistance to DOX in the xenograft tumor model.
Hepatocellular carcinoma (HCC) is one of the most common cancers worldwide with high mortality and poor prognosis (1). Multidrug resistance (MDR) in hepatocellular carcinoma reduces the efficacy of chemotherapy (2). One form of MDR is due to over-expression of P-glycoprotein (P-gp), an MDR1 gene product (3). P-gp is a transmembrane phosphor-glycoprotein capable of transporting out of the cell diverse chemotherapeutic drugs, leading to reduced intracellular drug concentration and cytotoxicity (4). Cisplatinum (CDDP) and doxorubicin (DOX) are first-line therapies for HCC and their efficacy is affected by MDR1 expression level in a patient's tumor (5-7). Cyclosporin A (8), verapamil (9) and COX-2, as well as P-gp inhibitors (10-13), have been used to reverse MDR with limited success. Small interfering RNA (siRNA), transfected into cancer cells, could specifically inhibit gene expression (14, 15). Our previous work has shown that MDR1-targeted siRNA reversed resistance to DOX in DOX-resistant HCC cells in vitro (16). In the present study, we used plasmid-based siRNA for the knockdown of the MDR1 gene and P-gp overexpression in a DOX resistant HCC in model in vivo.
Materials and Methods
Cell culture. DOX-resistant human hepatocellular carcinoma cell line Bel-7402/ADM was obtained from KeyGEN Biotch (Nanjing, China). Bel-7402/ADM cells were cultured in RPMI 1640 medium (Gibco, Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Hyclone, Logan, UT, USA) at 37°C in 5% CO2 and saturated humidity. All media was supplemented with penicillin/streptomycin (Gibco) and DOX (0.5 μg/ml).
Small interfering RNA and in vitro transfection. For the construction of MDR1 small interfering RNA (MDR1 siRNA) vector, three MDR1 siRNAs (MDR1 siRNA1, 5’-CACCG AAGACATGACCAGGTATGCCTTTCAAGAGAAGGCATACCTGGTCATGTCTTCTTTTTTG-3’; MDR1 siRNA2, 5’-CACC GCTGGTTGCTGCTTACATTCATTCAAGAGATGAATGTAAGCAGCAACCAGCTTTTTTG-3’; MDR1 siRNA3, 5’-CACCGGAGCTTAACACCCGACTTACTTCAAGAGAGTAAGTCGGGTGTTAAGCTCC TTTTTTG-3’) were synthesized and cloned into the pGPU6/GFP/Neo plasmid (Zoonbio Biotechnology, Shanghai, China). For in vitro transfection of Bel-7402/ADM cells, the cells were plated to 30-50% confluence and transfected with three MDR1 siRNA plasmids using Lipofectamine 2000 in Opti-MEM (Invitrogen, Carlsbad, CA, USA). The cells were examined by fluorescence microscopy 48 h after transfection. For selection, the cells were cultured in the presence of 500-2000 μg/ml G418 (Life Technologies, Grand Island, NY, USA) for 7 days. The expression of MDR1 mRNA and P-glycoprotein (P-gp) in Bel-7402/ADM cells was assessed by the reverse transcription-polymerase chain reaction (RT-PCR) and Western blotting, respectively, after 10 passages of selected green fluorescent protein (GFP)-expressing cells.
Animal care. Twenty-four BALB/C male nude mice, aged 4-6 weeks and weighing 20-25 g, were purchased from the Animal Department of the College of Medicine, Yangzhou University, Yangzhou, China. All mice were maintained in a high-efficiency particulate air (HEPA)-filtered environment at 24-25°C and humidity was maintained at 50-60%. All animals were fed with autoclaved laboratory rodent diet. Animal experiments were approved by the Animal Committee of Nanjing Origin Biosciences, China.
Xenograft mouse tumor model. A mouse model of human hepatocellular carcinoma was established with untransfected Bel-7402/ADM or Bel-7402/ADM transfected with MDR1 siRNA (Bel-7402/ADMsi). Stocks of Bel-7402/ADM and Bel-7402/ADMsi tumors were established by subcutaneously injecting 5×106 Bel-7402/ADM or Bel-7402/ADMsi cells in the flank of nude mice. The subcutaneous tumors were harvested at the exponential growth phase and resected under aseptic conditions. Necrotic tissues were removed and viable tissues were cut with a scissors and minced into 1-mm3 pieces. Animals were anesthetized by injection of 0.02 ml of a solution of 50% ketamine, 38% xylazine and 12% acepromazine maleate. Two tumor fragments were transplanted to the flank of nude mice with 8-0 surgical sutures. All surgical procedures and animal manipulations were conducted under HEPA-filtered laminar-flow hoods.
Treatment. Treatments were initiated when the average tumor size had reached 100 mm3. Mice with tumors derived from untransfected Bel-7402/ADM cells were randomly divided into three groups of six. Group 1 (Vehicle control-Bel-7402/ADM) received saline only; Group 2 (DOX-Bel-7402/ADM) received intravenous DOX treatment twice a week at 20 mg/kg/dose; Group 3 (MDR1 siRNA+DOX-Bel-7402/ADM) received intra-tumoral MDR1 siRNA treatment twice a week at 9.8 mg/kg/dose and intravenous DOX treatment twice a week at 20 mg/kg/dose. Six mice with tumors derived from Bel-7402/ADMsi cells were assigned to Group 4 (DOX-Bel-7402/ADMsi) and received intravenous DOX treatment twice a week at 20 mg/kg/dose. All treatments were continued for four weeks. Tumor growth was measured twice a week with calipers. Tumor volume was calculated using the formula (L x W2) x ½, where W and L represent the perpendicular minor dimension and major dimension, respectively. Animal body weights and clinical signs were recorded over the course of the experiments. All animals were sacrificed four weeks after treatment initiation. At autopsy, the tumors were removed and weighed.
RT-PCR. At the end of the study, tumor tissues were collected and snap-frozen in liquid nitrogen. Total RNA of tumor cells and tissues was isolated with Trizol reagent (Invitrogen, Carlsbad, CA, USA) and reversely transcribed using a PrimeScript RT-PCR kit (Takara, Shiga, Japan) according to the manufacturer's instructions, followed by PCR amplification with specific primers. The following primers were used to amplify most of the coding region of MDR1: sense, 5’-CCAAAGTCAACAAGGAGTGC-3’, and antisense, 5’-TCTTCAACAGTGGTTTATCGCA-3’). Samples were subjected to 30 cycles of PCR amplification using a thermo-cycler (Eppendorf, Hamburg, Germany). Each cycle included denaturation at 94°C for 30 sec, annealing at 52°C for 30 sec and primer extension at 72°C for 1 minute. An aliquot of each amplification mixture was subjected to electrophoresis on a 2% agarose gel and DNA was visualized by GelRed staining (Biosharp, Hefei, China). Gels were scanned and images were analyzed using UNSCANIT software (SilK Scientific, Orem, UT, USA). Results are expressed by relative mRNA expression (MDR1/β-actin).
Western blotting. P-gp expression in cancer cells and tissues was analyzed by Western blotting. Cells were lysed in 100 μl RIPA lysis buffer (50 mmol/l Tris-HCl, pH 7.5, 1% NP-40, 150 mmol/l NaCl, 1 mg/ml aprotinin, 1 mg/ml leupeptin, 1 mmol/l Na3VO4, 1 mmol/l NaF) at 4°C for 30 min. Cell debris was removed by centrifugation at 1,2000 × g for 20 min at 4°C. Frozen tissues were homogenized in tissue extraction buffer (50 mM Tris pH 7.4, 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, sodium orthovanadat, sodium fluoride, EDTA, leupeptin), supplemented with phenyl-methanesulfonyl fluoride solution (PMSF). The homogenate was incubated for 30 min on ice with frequent vortexing and, then, centrifuged at 12,000 × g for 30 min at 4°C. Protein concentrations were determined by the Bradford assay (Bio-Rad, Hercules, CA, USA). An equal amount of lysate (40 μg) was resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA). The membranes were blocked with 5% nonfat milk at room temperature for 1 h and then incubated for 2 h with primary antibodies to anti-P-170 glycoprotein (BD Biosciences, San Diego, CA, USA) and β-actin (Cell Signaling Technology, Boston, MA, USA). The membranes were then incubated for 1 h with an appropriate horseradish peroxidase-linked secondary antibody (Santa Cruz Biotechnology, Dallas, TX, USA). Electro-chemi-luminescence was performed according to the manufacturer's instructions using a ChemiDoc™ Touch Imaging System (Bio-Rad). Quantity One software (Bio-Rad) was used to quantify the density of bands.


MDR1 mRNA expression was down-regulated by MDR1 siRNA transfection in DOX-resistant human hepatocellular carcinoma cell line Bel-7402/ADM. A: MDR1 mRNA expression level was detected by RT-PCR. (M) DNA marker; (1) MDR1 siRNA1; (2) MDR1 siRNA2; (3) MDR1 siRNA3; (4) Vector control; (5) Untransfected control. B: Relative MDR1 mRNA expression in each group. *p<0.05, when compared to vector and untransfected control.
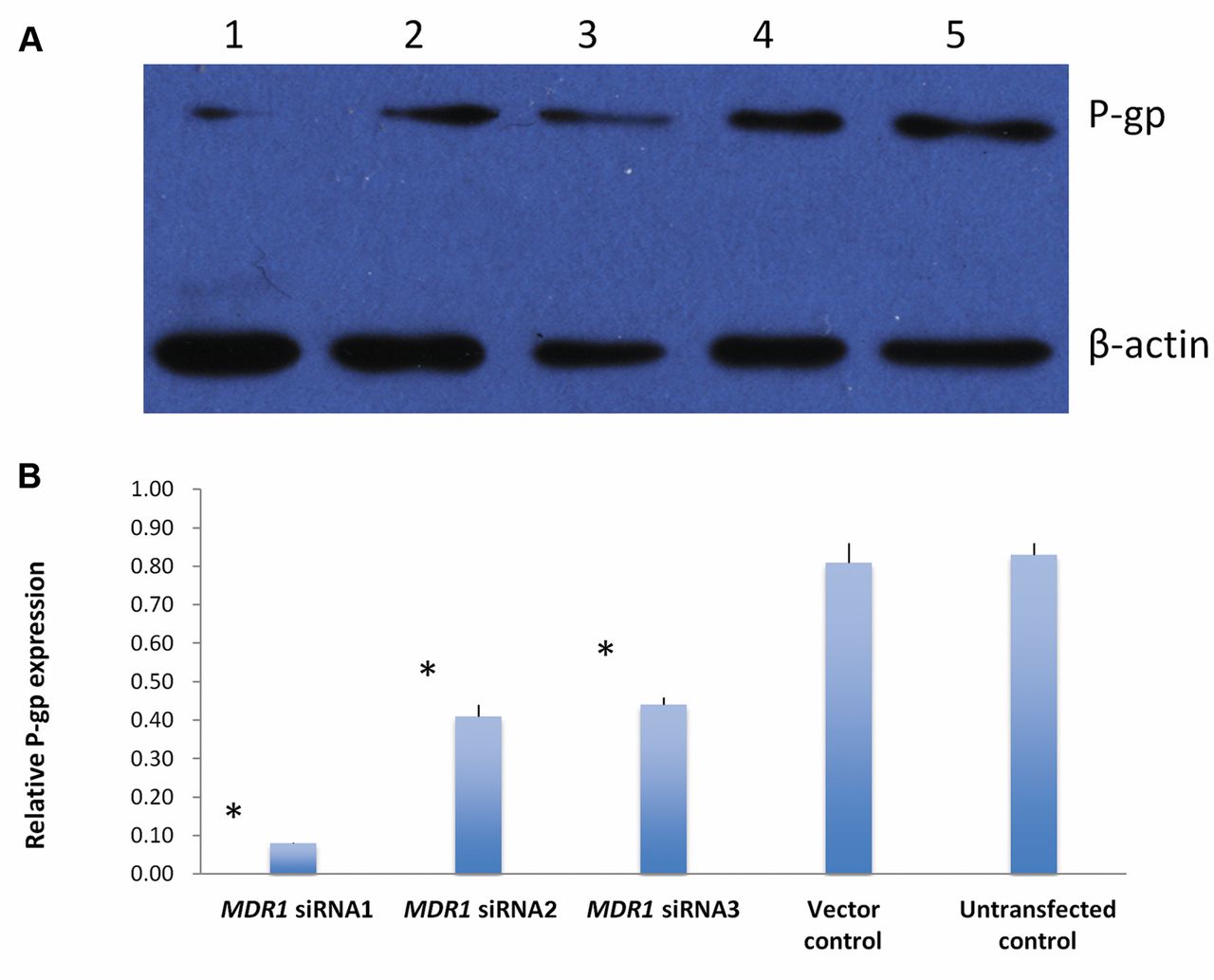
P-gp expression was reduced by MDR1 siRNA transfection in DOX-resistant human hepatocellular carcinoma cell line Bel-7402/ADM. A: P-gp expression level was detected by Western blotting. (1) MDR1 siRNA1; (2) MDR1 siRNA2; (3) MDR1 siRNA3; (4) Vector control; (5) Untransfected control. B: Relative P-gp expression. *p<0.05, when compared to vector and untransfected control.
Immunohistochemistry. The tumor tissue was collected and fixed in 10% buffered formalin and paraffin-embedded at the end of the study for immunohistochemistry analysis. The sections were incubated with primary antibodies against P-170 glycoprotein (BD Biosciences) overnight at 4°C after permeabilization with a solution of 0.1% sodium citrate and 0.1% Triton X-100 and blocking with 10% rabbit serum. After washing in PBS, the slices were incubated with horseradish peroxidase-labeled secondary antibody (1:200; Maixin Bio-Tech Co., Ltd, Fuzhou, China) for 30 minutes at room temperature. After color development using 3,3-diaminobenzidine (DAB) for10 min, the slices were counterstained with hematoxylin. The slides were viewed at 400× magnification and positive cells were recognized by the appearance of brown-stained cells. Expression levels were quantified by the average optical density (AOD) of the positive cells in 5 fields/sample with Image-Pro Plus 6.0 software (Media Cybernetics, Silver Springs, MD, USA).
Statistical analysis. Data, expressed as means ± standard deviation (SD), were analyzed by one-way analysis of variance (ANOVA) using the SPSS software version 16.0 (SPSS Inc., Chicago, IL, USA), where p<0.05 was considered to be statistically significant.
Results
Down-regulation of MDR1 mRNA expression by MDR1 siRNAs in Bel-7402/ADM cells. Fluorescence microscopy and RT-PCR were used to assess MDR1 siRNA transfection efficacy and MDR1 mRNA expression, respectively. As shown in Figure 1, efficient MDR1 siRNA transfection of Bel-7402/ADM cells was confirmed by strong GFP expression (Figure 1B). Significant down-regulation of MDR1 mRNA expression was obtained by transfection with three MDR1 siRNAs in Bel-7402/ADM cells as compared to untransfected and vector controls (Figure 2) (p<0.05). The relative MDR1 mRNA expression level was 0.09 ± 0.002 for MDR1 siRNA1; 0.33 ± 0.021 for MDR1 siRNA2; and 0.20 ± 0.015 for MDR1 siRNA3. MDR1 siRNA1 showed the most silencing efficiency on Bel-7402/ADM cells among three MDR1 siRNAs (p<0.05).

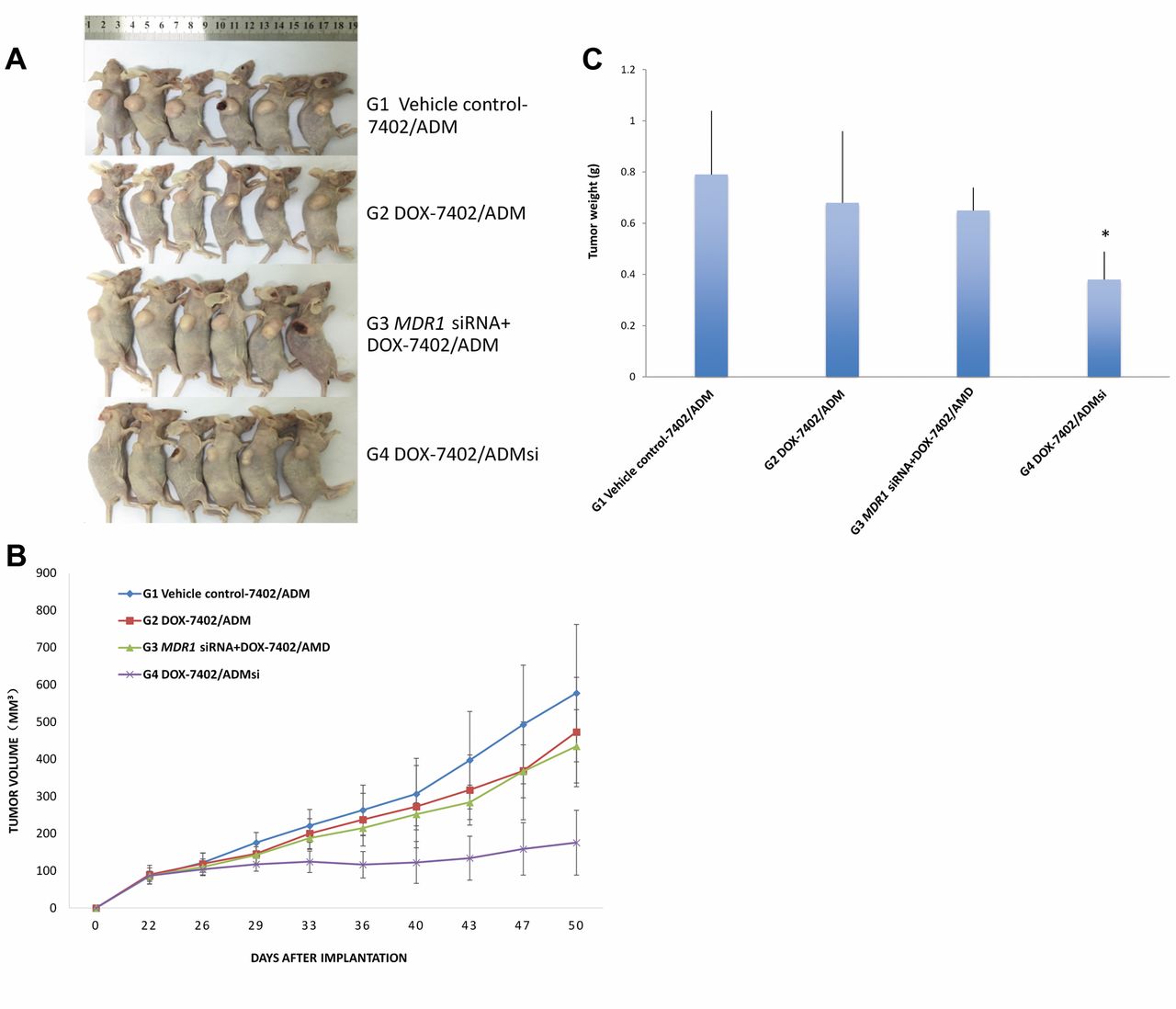
Reversal of multidrug resistance by MDR1-siRNA in the mouse model of DOX-resistant human hepatocellular carcinoma. A: Images of tumor-bearing mice in each group. B: Tumor growth curves after treatment with MDR1-siRNA and DOX. C: Final tumor weight in the treated groups of mice. Data are represented as mean ± SD of 6 animals of each group. *p<0.05, when compared with vector and untransfected-control groups.
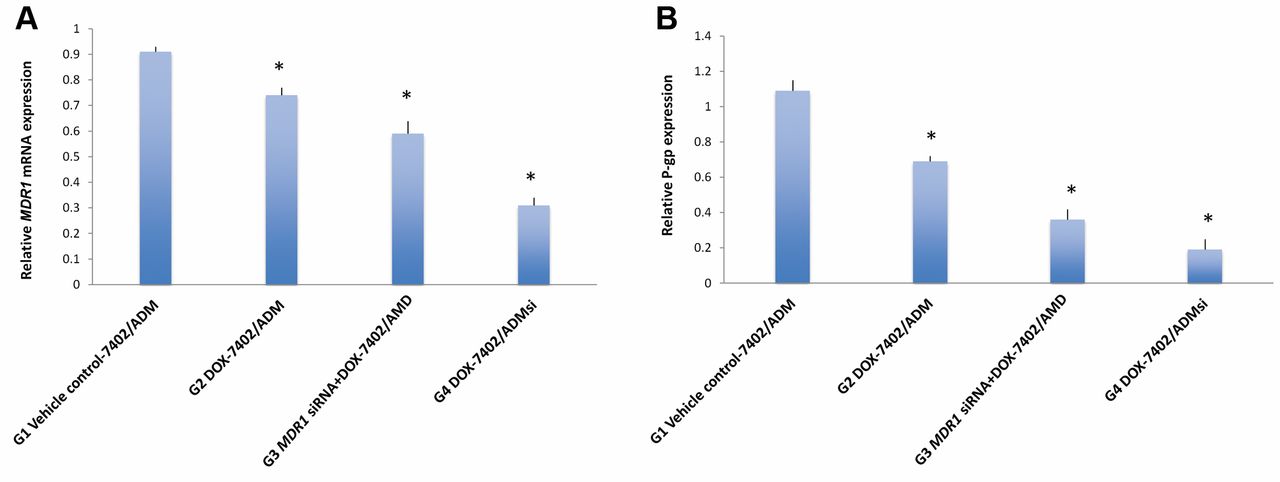
Down-regulation of MDR1 mRNA and P-gp expression by MDR1 siRNA in the mouse model of DOX-resistant human hepatocellular carcinoma. A: MDR1 mRNA expression in the tumor detected by RT-PCR. Data are represented as mean ± SD of 6 animals of each group. *p<0.05, when compared with vector and untransfected control groups. B: P-gp expression in the tumor detected by Western blotting. Data are represented as mean ± SD of 6 animals of each group. *p<0.05, when compared with vector and untransfected control groups.

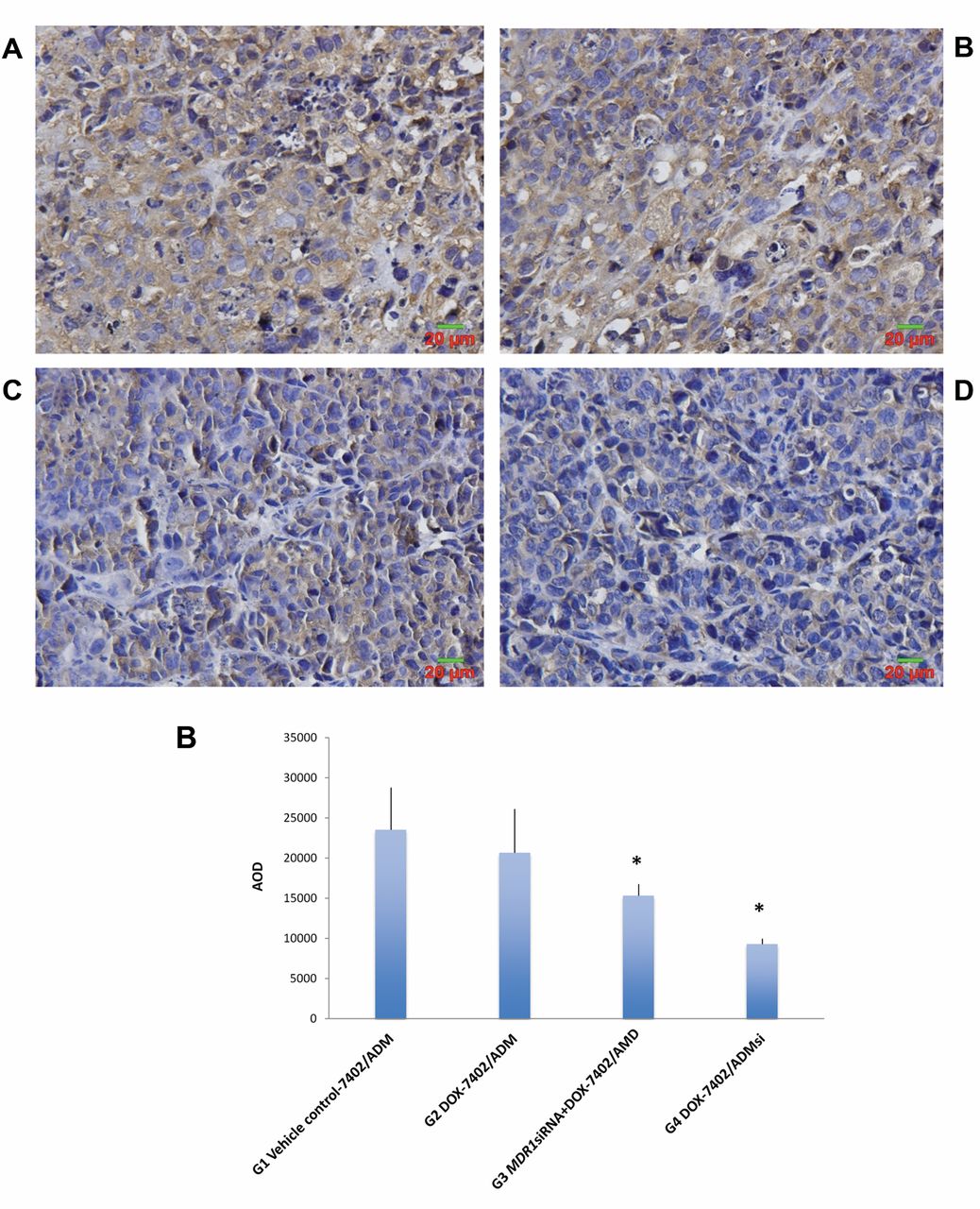
Immunohistochemistry analysis of P-gp expression in the mice of treated groups. A: Representative immunohistochemistry staining for P-gp in the tumors (a): G1, (b): G2, (c): G3, and (d): G4. Magnification ×400. B: Average optical density (AOD) of immunohistochemistry staining of tumor samples. Data are represented as mean ± SD of 6 animals of each group. *p<0.05, when compared with vector and untransfected control groups.
Reduction of P-gp expression by MDR1 siRNA in Bel-7402/ADM cells. The effect of MDR1 siRNAs on P-gp expression was determined by Western blotting. As shown in Figure 3, significant reduction of P-gp expression was obtained by transfection with three MDR1 siRNAs in Bel-7402/ADM cells as compared to untransfected and vector control (p<0.05). More reduction of P-gp expression was achieved by MDR1 siRNA1 (0.08 ± 0.001) than MDR1 siRNA2 (0.41±0.025) and MDR1 siRNA3 (0.44 ± 0.034) (p<0.05).
MDR1 siRNA reversed multidrug resistance to DOX in vivo. Two mouse tumor models of hepatocellular carcinoma were used to assess the effect of MDR1 siRNA on reversing multidrug resistance in vivo. One model was established with untransfected Bel-7402/ADM cells. The Bel-7402/ADM cells transfected with the most efficient MDR1-siRNA1 vector was selected to establish another tumor model (Bel-7402/ADMsi). As shown in Figure 4, no significant anti-tumor efficacy was found in the DOX treatment group (Group 2), as compared to vehicle the control (Group 1), in the tumor model established from untransfected Bel-7402/ADM (p>0.05), suggesting the Bel-7402/ADM tumor is multidrug resistant to the DOX. Significant tumor growth inhibition was demonstrated in the DOX treatment group (Group 4) in the tumor model from Bel-7402/ADMsi as compared to DOX treatment group (Group 2) in the tumor model established from untransfected established Bel-7402/ADM (p<0.05), indicating reversal of multidrug resistance in tumor by MDR1 siRNA. However, the combination treatment of MDR1 siRNA and DOX (Group 3) showed no significant anti-tumor efficacy as compared to the DOX treatment group (Group 2) in the tumor model established from untransfected Bel-7402/ADM (p>0.05), indicating poor in vivo transfection efficiency of MDR1 siRNA.
Reversal of multidrug resistance is associated with reduced expression of MDR1 mRNA and P-gp by MDR1 siRNA. At the end of the study, tumor tissues from all groups were collected. MDR1 mRNA expression in the tumors was analyzed by RT-PCR. As compared to vehicle control, significant down-regulation of MDR1 mRNA expression was found in the DOX treatment group (Group 2) and the combination treatment group (Group 3) in the tumor model developed from untransfected Bel-7402/ADM, as well as the DOX treatment group (Group 4) in the tumor model developed from Bel-7402/ADMsi (p<0.05) (Figure 5A). The tumors in Group 4 showed significantly lower MDR1 mRNA expression than the tumors from Group 2 and 3 (p<0.05). P-gp expression in tumors was analyzed by both western blotting (Figure 5B) and immunohistochemistry (Figure 6). As compared to vehicle control, significant reduction of P-gp expression was found in the DOX treatment group (Group 2) and combination treatment group (Group 3) in the tumor model established from untransfected Bel-7402/ADM, as well as the DOX treatment group (Group 4) in the tumor model developed from Bel-7402/ADMsi (p<0.05). Similar to MDR1 mRNA expression, the tumor in Group 4 showed significantly lower P-gp expression than the tumors from Group 2 and 3 (p<0.05). These results indicate association of reduced expression level of MDR1 mRNA and P-gp with reversal of multidrug resistance.
Discussion
In cancer, over-expression of MDR1 P-gp is a possible cause of chemotherapy-based treatment failure (17). The gene silencing induced by RNAi was shown to be specific and potent, offering an alternative strategy for overcoming drug resistance (18). In vitro, suppression of P-gp using various siRNA delivery and siRNA expression systems has been relatively successful (19-21).
In the present study, we synthesized three siRNAs targeting the MDRl gene and screened their silencing efficiency in the DOX-resistant human hepatocellular carcinoma cell line Bel-7402/ADM. It has been found that MDRl mRNA expression of Bel-7402/ADM was down-regulated after MDR1 siRNA transfection. MDR1 gene silencing by MDR/siRNA was accompanied by significant P-gp reduction. The three siRNAs showed different RNAi efficiencies, although all three siRNAs were efficient to reduce mRNA and P-gp expression of Bel-7402/ADM. Among them, MDR1 siRNA1 had the greatest silencing effect. This could be attributed to the secondary structure of MDR1 cDNA, stem-loop structures and the extent of base pairing and hydrogen bonding outside the targeted site (22, 23).
In experiments to evaluate the reversal effect of MDR1 siRNA on chemotherapy resistance in the DOX-resistant tumor implant, the MDR1 siRNA found most effective for in vitro down-regulation of MDR1 mRNA and P-gp was further investigated in a xenograft model. MDR1 siRNA transfected and untransfected Bel-7402/ADM cells were implanted into nude mice followed by DOX treatment. In the mice implanted with MDR1 siRNA-transfected Bel-7402/ADM cells, DOX treatment resulted in significant reduction in tumor size, while MDR1 mRNA and P-gp were significantly decreased. These results demonstrate in vivo reversal of multidrug resistance by MDR1 siRNA in vitro transfection of Bel-7402/ADM cells and the association of MDR1 mRNA and P-gp expression level with multidrug resistance.
Limitations exist for in vivo siRNA delivery (24). Systemic delivery of siRNA can lead to rapid siRNA degradation in the physiological milieu (25). Localized injection, on the other hand, could minimize the degradation of the injected siRNA and is, therefore, beneficial for superficial tumors (26). In thisstudy, we intratumorally injected MDR1 siRNA into untransfected Bel-7402/ADR tumors in vivo followed by DOX treatment. No significant tumor growth inhibition was found, although down-regulation of MDR1 mRNA and P-gp were observed. This result suggests that intratumoral injection of MDR1 mRNA can reduce MDR1 expression to some degree, but might not be efficient enough to effectively silence MDR1 expression in order to reverse multidrug resistance in a tumor. Therefore, a more stable MDR1 siRNA vector or delivery system needs to be developed for more effective in vivo transfection of multidrug resistant tumors.
In conclusion, in vitro transfection of siRNA vectors targeting the MDR1 gene can effectively silence MDR1 mRNA and knock down P-gp expression in DOX-resistant human hepatocellular carcinoma Bel-7402/ADR cells and thereby confer sensitivity to DOX in a xenograft tumor model.
Footnotes
This article is freely accessible online.
Conflicts of Interest
None of the Authors has a conflict of interest in regard to this study.
- Received March 23, 2016.
- Revision received April 27, 2016.
- Accepted April 28, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}