


Abstract
Ceramide and sphingosine 1-phosphate (S1P) are sphingolipid metabolites with important signaling functions. Ceramides promote apoptosis, whereas S1P favors proliferation, angiogenesis and cell survival. The balance between these opposing signaling functions is referred to as the sphingolipid rheostat. A shift in this balance toward S1P is seen in glioblastoma (GBM) and other cancers, and results in tumor cell survival and resistance to chemotherapy. Sphingosine kinase (SK), the enzyme responsible for transforming sphingosine into S1P, plays the critical role in modulating the balance between S1P and ceramides. Chemotherapeutic agents or radiation therapy may induce short-term responses in GBM patients by increasing ceramide levels. However, we believe that the enzyme SK may cause the increased ceramide to be metabolized to S1P, restoring the abnormally high S1P to ceramide balance, and that this may be part of the reason for the near-100% recurrence rate of GBM. The use of maintenance therapy with an SK inhibitor, in patients with GBM who have tumor reduction or stable disease after therapy, should be investigated.
- Glioblastoma
- maintenance therapy
- tumor recurrence
- sphingosine kinase
- sphingosine 1-phosphate
- ceramide
- chemotherapy
- sphingolipid rheostat
- review
Background
Glioblastoma multiforme (GBM) is an aggressive primary brain neoplasm with a median patient survival of only 14.6 months (1, 2). Presenting symptoms include nausea, vomiting, blurred vision, headaches, and drowsiness. The tumor is particularly resistant to therapy. Standard initial treatment is maximal tumor resection followed by radiation therapy, with simultaneous administration of temozolomide (TMZ), an oral alkylating agent and imidazotetrazine derivative of dacarbazine (3-9). Recurrent GBMs are most commonly treated with bevacizumab (avastin), which suppresses angiogenesis, or lomustine, a lipid-soluble, alkylating nitrosourea which crosses the blood-brain barrier (10-12). However, these agents are only effective in a small minority of patients, and then only for a few months. Even with newer chemotherapy drugs and advances in surgical methods, overall patient survival rates continue to be extremely poor, and no cure for GBM exists (13-18). Alternative approaches, such as immunotherapy, oncogene therapy or molecular targeting agents, are being investigated, but so far none have been shown to have a significant impact on response rate or survival (19-22).
Ceramide and Sphingosine-1-Phosphate
Sphingolipids are components of the eukaryotic membrane. The major sphingolipid, sphingomyelin, is particularly found in the membranes of nerve cells. Sphingomyelins can be hydrolized by sphingomyelinases to ceramides and phosphorylcholine (23). Ceramides are an extremely important group of molecules consisting of sphingosine bases linked to fatty acids of different chain lengths.
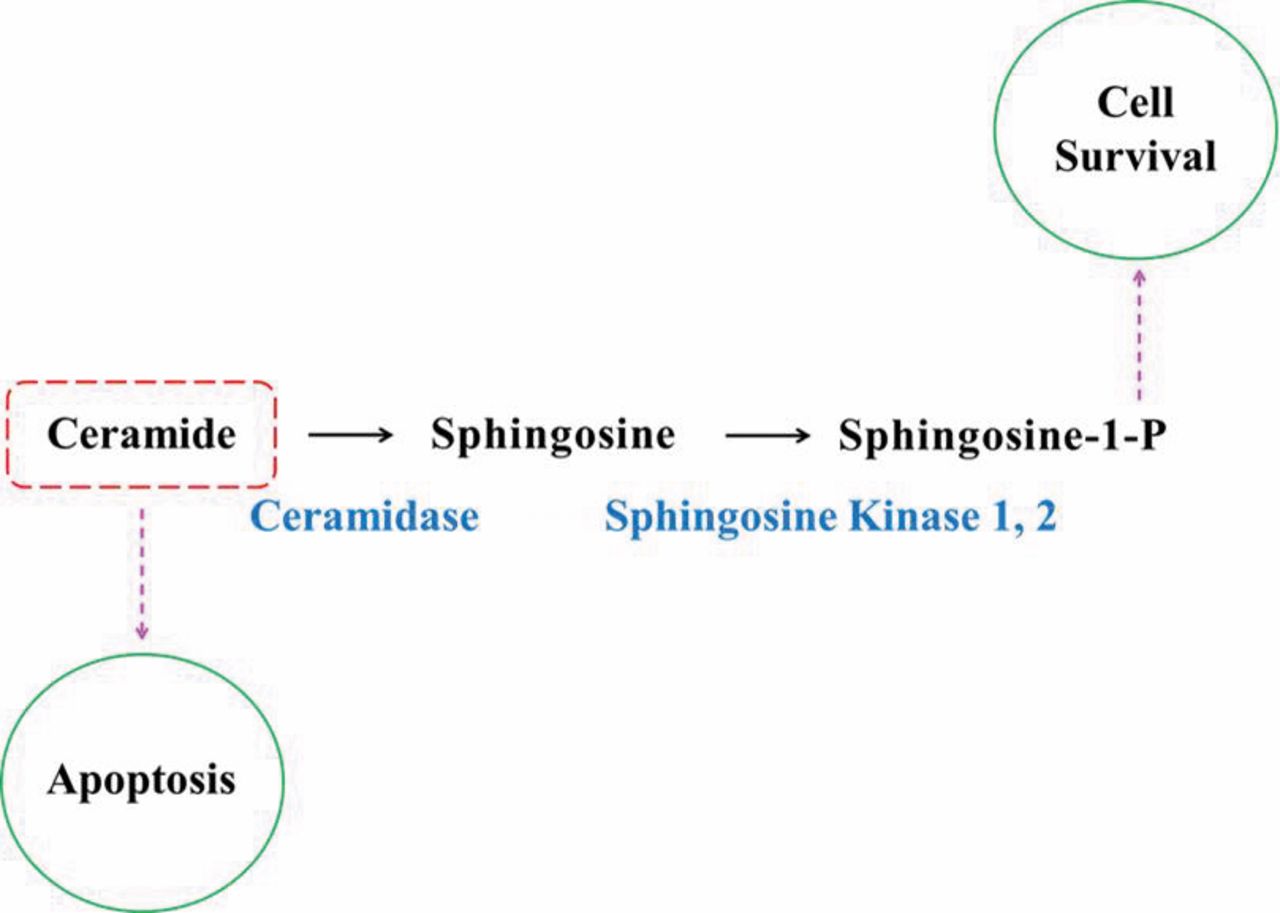
There are three major pathways for the generation of ceramide, the de novo, the sphingomyelinase and the salvage pathways (24-25) (Figure 1). In the de novo pathway, ceramides are generated from palmitate and serine in a series of steps initiated by the key enzyme serine palmitoyltransferase (24-26). In the sphingomyelinase pathway, sphingomyelin is hydrolyzed by sphingomyelinase (SMase) (27, 28). In the salvage pathway, ceramides are formed from the sphingolipid metabolite sphingosine by ceramide synthase (29). Sphingosine-1-phosphate (S1P) is formed when ceramide is broken down by ceramidase, and the resulting sphingosine molecule is phosphorylated by the enzyme sphingosine kinase (30) (Figure 2).
Historically, ceramide and S1P were thought merely as components of the cell membrane. In the 1990s, however, Obeid et al. showed that cell death could be caused by increases in ceramide, and Zhang et al. reported on the role of S1P in modulating cellular proliferation (31, 32). It is now known that ceramide possesses pro-apoptotic signaling functions, whereas S1P plays an important role in proliferation, angiogenesis and cell survival (33-40). S1P is also responsible for T-cell maturation (41, 42). The pro-apoptotic functions of ceramides can occur through numerous mechanisms, including increasing protein phosphatase 2A (PP2A), a tumor suppressor, through interaction with microtubule-associated protein 1 light chain 3 beta lipidation (LC3B-II) autophagolysosomes, through activation of protein kinase C, and by down-modulation of the gene c-myc (43-45). Ceramide-increasing agents have been shown to cause destruction of GBM stem cells, the persistence of which are a major cause of GBM recurrence after therapy (46-49).
Sphingosine Kinase
Sphingosine kinase (SK), the signaling enzyme responsible for transforming sphingosine into S1P, plays a critical role in maintaining the balance between ceramides and S1P (50, 51). In humans, there are two forms of SK. SK1 is prominent in the cytoplasm and has been studied extensively. Cancer cell growth and survival are stimulated by up-regulation of SK1 (52-58). For example, increased amounts of SK1 in fibroblasts can cause their malignant transformation to fibrosarcoma cells (54). Over-expression of SK1 has been detected in carcinomas of the breast, prostate, colon, esophagus and lung, as well as in other cancers (59-71). Bektas et al. showed that melanoma cells resistant to therapy have a much higher S1P to ceramide ratio and express more SK1 than sensitive melanoma cells, and that increased SK1 levels correlated with overexpression of anti-apoptotic protein, B-cell lymphoma 2 (Bcl-2) (68). Benign adenomas of the colon express more SK1 than normal colon cells, and more aggressive metastatic cancers express more SK1 than cancers that have not metastasized (54). SK2 is localized in the nucleus and plasma membrane, and much less in the cytoplasm, and occurs predominantly in the liver and the kidneys. SK2 appears to have both cell-protective and pro-apoptotic functions (72-74). For example, Maceyka et al. reported that, unlike SK1, SK2 may also cause an increase in the activity of ceramide synthase and the salvage pathway, thus at least partially counter-acting its ceramide-reducing effect. They speculated that this may be due to the different locations where SK1 and SK2 predominate (74). Despite this, some studies have shown that depletion of SK2 has greater apoptotic effects against cancer cells than depletion of SK1 (50).
Both SK1 and SK2 play important roles in GBM development, progression and resistance to treatment. It has been reported that S1P stimulates invasiveness in human GBM cell lines and tissues through the receptors S1P1-5 (75-79). On the other hand, Yoshida et al. demonstrated that while S1P2 and S1P3 were increased in patients with GBM, S1P1 levels were decreased, and lower levels in S1P1 correlated with poor patient survival (80). Other studies have shown that SK levels are markedly increased in patients with GBM (81-83). Van Brocklyn et al. reported that patients with GBM and lower SK1 expression in their cancers survived three times longer than patients with high SK1 (78). Anelli et al. showed that SK1 is over-expressed during hypoxia in U87MG glioma cells (83). Quint et al. investigated the role of SK1, SK2, and of S1P receptors in primary, secondary, and recurrent GBM tissue samples, and showed that SK1 and S1P receptors were overexpressed as much as 44 fold compared to normal brain tissue (84). With a 25-fold increase, SK2 was highest in primary tumors. Abuhussain et al. showed that S1P levels are favored over ceramide levels in patients with glioma, and that increased S1P correlates with increased histological tumor grade (85). In that study, S1P levels were also nine times higher in areas of tumor compared to areas of normal gray matter, whereas ceramides, in particular C18-ceramide, were five-fold lower.
SK Inhibitors
Two sphingosine kinases inhibitors, D,L-threodihydrosphingosine (safingol) and N,N,N-trimethylsphingosine, have been investigated for a number of years as possible anti-cancer agents (86-92). While these agents did not appear to have significant anti-tumor activity alone, there was evidence that they might potentiate the anticancer effects of known chemotherapy drugs. These inhibitors are not specific to SK, and can affect many protein and lipid kinases (93). An important SK inhibitor, fingolimod (FTY720, gilenya), has been studied in numerous diseases for more than 20 years, and is now an approved treatment for patients with multiple sclerosis. Fingolimod is phosphorylated by SK2 to fingolimod-phosphate, which binds to S1P receptors and, in turn, inhibits both SK1 and SK2. Among its actions, fingolimod-phosphate is a superagonist of the S1P receptor on lymphocytes, and can prevent these cells from leaving the lymph node (87, 94-96). It is lipophilic and crosses the blood-brain barrier, and also binds to S1P receptors on neural and other cells. There is evidence for anticancer activity of this agent in experimental models, which may be secondary to its suppressant effects on SK (87, 97-98). However, not only does fingolimod act on SK, it also affects multiple other enzymes that can alter ceramide/S1P balance, including SMase, ceramide synthase, acid ceramidase, S1P lyase and S1P phosphatases (97, 99-100). For example, fingolimod inhibits both SMase and ceramide synthase, thus partially counteracting the ceramide-increasing effect of SK inhibition (99). In addition, because fingolimod has many “off-target” actions, it has been shown to cause occasional side-effects. Severe infections, skin cancers, cardiac rhythm disturbances, macula edema, and, in rarer cases, neurological abnormalities such as opthalmoplegia, ataxia and, dysarthria, have been reported (94, 96, 100). A number of other commonly-used FDA approved agents, including antihistamines, antimalarials, antineoplastics and cardiac medications have “off-target” effects and can affect SK; all of these agents have also been shown to act on multiple other enzymes involved in sphingolipid metabolism (Table I).

Three major pathways for the generation of ceramide.
More recently, numerous, newer and purer, inhibitors have emerged (90, 118-121). Many of these agents have been shown not to affect the broad range of enzymes that the earlier-studied SK inhibitors do. French et al. reported anti-tumor activity with three SK1 inhibitors (SKI-I: 5-naphthalen-2-yl-2H-pyrazole-3-carboxylic acid (2-hydroxy-naphthalen-1-ylmethylene)-hydrazide; SKI-II: 4-[4-(4-chloro-phenyl)-thiazol-2-ylamino]-phenol; SKI-V: 2-(3,4-dihydroxy-benzylidene)-benzofuran-3-one)) in a JC mouse mammary adenocarcinoma cell line and in a syngeneic BALB/c mouse solid tumor model of JC mammary adenocarcinoma cells (86). SKI-II was found to be especially effective in this model. Paugh et al. studied a SK1 specific inhibitor ((2R,3S,4E)-N-methyl-5-(4’-pentylphenyl)-2-aminopent-4-ene-1,3-diol (SK1-I) in human leukemic cell lines and acute myelogenous leukemia xenografts, and showed that SK1-I blocked tumor growth and induced apoptosis. Unlike early SK inhibitors, SK1-I does not inhibit SK2, protein kinase B, protein kinase C or other serine/threonine kinases (122). The selective SK2 inhibitor, ABC294640 (yeliva), has been studied in prostate cancer cell lines and TRAMP-C2 xenografts, and found to cause reduced cell viability and decreased expression of c-myc. This agent is currently part of a phase I clinical trial in advanced solid tumors (123). Neubauer et al. showed that selective targeting of SK2, instead of SK1, could provide additional therapeutic benefits (124). Schrecengost et al. used ABC294640 in xenograft prostate cancer models, and reported that it significantly blocked cancer growth (125).

Sphingosine-1-phosphate is generated from sphingosine by way of ceramide.
Commonly used agents with SK activity.
A few studies of SK inhibitors have been performed in GBM. Van Brocklyn et al. have shown that SK isoforms play a critical role in the growth and aggressiveness of GBM cells in vitro (78). They reported that the SK1 inhibitor (2-(p-Hydroxyanilino)-4-(p-chlorophenyl)thiazole) significantly decreased the rate of proliferation in the GBM cell lines U-87 MG, U-1242 MG and M059K (78, 89). Similarly, Bektas et al. used the SK1 inhibitor 2-(p-hydroxyanilino)-4-(p-chlorophenyl)thiazole to promote cancer cell death in the TMZ-resistant GBM cell lines U251 and D54MG (89, 126). Kapitonov et al. used the inhibitor SK1-I against LN229 and U373 GBM cell lines, non-established human GBM6 cells, and GBM xenografts, and showed that targeting SK1 inhibits protein kinase B (Akt) signaling, prompts apoptosis, and suppresses tumor development in human GBM cell lines and GBM xenografts (127). Inhibition of S1P resulted in blocked angiogenesis. Other investigators have reported similar results (128-134) (Table II).
Preventing loss of ceramide-induced tumor response. As noted, though ceramides and S1P have opposing signaling functions, they are closely connected. S1P may be dephosphorylated to form sphingosine, and sphingosine then re-acylated to form ceramide. Similarly, sphingosine can be phosphorylated by SK to produce S1P. Cuvillier et al. were the first to use the term “sphingolipid rheostat” to describe the balance between ceramide and S1P, and concluded that a shift in this balance plays a role in the determination of the cell's fate (135). An increase in ceramides predisposes to cell death, whereas an excess of S1P is protective. A number of diseases or disease conditions are associated with abnormalities of the rheostat (136-140). SK determines whether S1P or ceramide will dominate, and the fate of the cell is determined by the greater relative content of these opposing signaling molecules. Cancer is associated with an increase in S1P within the cell, and with decreases in ceramide (141-145).
SK inhibitors alone will increase ceramide levels, but not as much as when given in combination with an agent which stimulates SMase or ceramide synthase. SK inhibitors have been used in combination with cytotoxic chemotherapy with the goal of increasing ceramides (146-149), and we have previously suggested that increased apoptosis of GBM cells may be achieved using combinations of agents which each increase ceramides (150). Noack et al. used SKI-II with TMZ against the human GBM cell line NCH82, and found that the combination enhanced caspase-3 dependent cell death and autophagy (151). Similarly, Riccitelli et al. showed that an SK1 inhibitor increased chemo-sensitivity to TMZ in a human GBM cell line (152). Estrada-Bernal et al. used FTY720 in combination with TMZ in xenografts of GBM stem cells, and found that tumor volume significantly decreased and mouse survival times increased (130). Treatment with FTY720 with TMZ resulted in longer survival times compared to FTY720 or TMZ alone. However, we believe that the optimal use of SK inhibitors is not as ceramide-inducing agents, but rather in preventing the increased ceramides that are produced after chemotherapy or radiation therapy from being later metabolized to S1P. For example, if this theory is correct, maintenance therapy with SK inhibitors could extend survival of patients with GBM who have first been treated with radiation therapy and temozolomide and have achieved a response or stable disease.
SK inhibitors as single agents in GBM.
In conclusion, there is, we believe, an important mechanism by which tumor progression can occur after response to therapy. Over time, the ceramides that are induced by chemotherapy or radiation therapy are converted to S1P through the actions of SK. Then the excess of S1P over ceramide in the tumor, which was in effect before treatment, is restored. This results in loss of response, as is typically seen after a short period of time in GBM and other solid tumors. Loss of response to ceramide-inducing agents has been seen in patients who tumors express high levels of SK. For example, patients with estrogen receptor-positive breast carcinoma treated with tamoxifen, an agent which decreases acid ceramidase (138), had shorter recurrence times if their cancers had higher SK levels (87). Likewise, patients with head and neck carcinoma with high levels of SK had a much shorter time to progression after radiation therapy (153). An SK inhibitor might prevent the ceramide from being later converted to S1P, reducing the chance of loss of response. Indeed, one of the reasons that patients with chronic myelogenous leukemia (CML) treated with tyrosine kinase inhibitors (TKIs) have a much lower recurrence rate than do patients with solid tumors or CML patients treated with chemotherapy or interferon, may be because TKIs not only increase ceramide by stimulating ceramide synthase, but also inhibit SK (65, 154-156). In view of the near-universal tendency of GBM to recur, long-term maintenance therapy with an SK inhibitor may be needed to prevent relapse and progression of disease.
Footnotes
This article is freely accessible online.
Conflicts of Interest
Dr. Peter P. Sordillo is a member of the Scientific Advisory Board of SignPath Pharma, a developmental stage biotechnology company that is studying liposomal curcumin, liposomes and other agents. Dr. Helson is CEO of SignPath Pharma. Laura Sordillo reports no conflicts.
- Received March 4, 2016.
- Revision received April 12, 2016.
- Accepted April 13, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Intense Uptake of Liposomal Curcumin by Multiple Myeloma Cell Lines: Comparison to Normal Lymphocytes, Red Blood Cells and Chronic Lymphocytic Leukemia Cells
- {alpha}-Mangostin protects against high-glucose induced apoptosis of human umbilical vein endothelial cells
- Aneuploid Cell Survival Relies upon Sphingolipid Homeostasis
- The Kynurenine Pathway: A Primary Resistance Mechanism in Patients with Glioblastoma