


Abstract
Aim: Ran binding protein M (RanBPM) is a ubiquitous, nucleocytoplasmic protein that serves as a scaffolding molecule. This study aimed to investigate the role of RanBPM in gastric cancer. Materials and Methods: RanBPM expression in human gastric cancer tissue samples was analyzed using real-time polymerase chain reaction. The effect of RanBPM on cellular functions was examined in RanBPM-knockdown gastric cells and with in vitro cell functional assays. Results: Gastric tumors with distant metastases expressed lower levels of RanBPM transcripts compared to tumours without detectable metastases (p=0.036). RanBPM knockdown in gastric cancer cells reduced adhesion and promoted survival of gastric cancer cells after exposure to methotrexate and fluorouracil. Conclusion: RanBPM levels were reduced in gastric tumors with distant metastases. This suggests that loss of RanBPM expression may play an important role in gastric cancer tumor development and metastasis. Reduced RanBPM expression is also associated with chemoresistance of gastric cancer cells.
Gastric cancer remains the fourth most common cancer and the second leading cause of cancer-related deaths worldwide (1). Although its overall incidence and mortality rates have decreased, there has been a paradoxical increasing incidence of non-cardia, intestinal-type gastric cancer in younger patients over the past three decades in the USA, Spain and several European countries (2, 3). Associated genes and proteins such as human epidermal growth factor receptor 2, vascular endothelial growth factor A, fibroblast growth factor receptor and hepatocyte growth factor receptor have become targets for biological therapeutic agents (4, 5). Unfortunately, little progress has been made in the treatment of advanced or metastatic gastric cancer and median overall survival (OS) remains less than 1 year (6). Therefore, there has been much interest in the identification of potential new biomarkers or molecular targets to aid early diagnosis and facilitate the development of novel biological therapies.
Ran binding protein M (RanBPM), also as known as RanBP9, is a ubiquitously expressed nucleocytoplasmic protein (7, 8). The RanBPM gene is located on human chromosome 6p23 and encodes a protein of 729 amino acids in length with a molecular weight of 90 kDa (9). RanBPM contains three conserved domains, namely specific protein 1 and ryanodine receptor (SPRY) domain, platelet-activating factor acetylhydrolase IB subunit alpha (LIS1)-homology motif/C-terminal to LISH (LISH/CTLH) domain and CT11-RanBPM (CRA) domain (10). The SPRY domain mediates protein–protein interaction, for example, the interaction between RanBPM with N-methyl-N'-nitro-N-nitrosoguanidine (MNNG)-HOS transforming gene (MET) which is the receptor of hepatocyte growth factor (11). Proteins containing LISH/CTLH domain are involved in microtubule dynamics, cell migration and protein homodimerisation (12, 13). Both SPRY and LISH/CTLH domains are also involved in the cytoplasmic retention of the protein (14). The CRA domain mediates interaction between RanBPM and fragile X mental retardation protein, which may modulate RNA-binding of the latter (15). In addition to the conserved domains, there exist six prototypical SH3-binding domains at the proline-rich N-terminus of the RanBPM protein that are predicted to bind with high affinity to proto-oncogene SRC (short for sarcoma) and growth factor receptor-bound protein 2 (10). RanBPM protein contains two motifs that are important for its nuclear localization. They are a proline/glutamine-rich motif at the extreme N-terminus which is predominantly responsible for the nuclear localization of RanBPM, whilst the other motif at the carboxy-terminus also contributes to a lesser extent (14).
RanBPM is ubiquitously expressed in human and murine cell lines and has been observed in the nucleus, cytoplasm, plasma membrane and at cell junctions (8, 11, 16-18). RanBPM can be part of a large protein complex that is named the CTLH complex (19) and is also known as the muskelin/RanBPM/CTLH complex (MRCTLH) (20). RanBPM has been implicated in a variety of cellular processes, including cell adhesion, migration, microtubule organization and gene transcription (7, 15, 21-25). RanBPM may regulate immune cell functions by interacting with leukocyte- or lymphocyte-function-associated antigen 1 (17) and cluster of differentiation 39 (CD39) (26). RanBPM can also regulate splicing of target mRNA thus contributing to normal spermatogenesis and male fertility (27). RanBPM is also critical for the nerve system due to its interaction with neural cell adhesion molecule L1 and plexin A (28, 29). RanBPM is able to simultaneously inhibit cell adhesion and promote amyloid β-peptide production thus contributing to the pathogenesis of Alzheimer's disease (30, 31). Targeting RanBPM has shown certain therapeutic potential by rescuing the neurodegenerative changes (32).
RanBPM promotes pro-apoptotic pathways in response to DNA damage (33). P73l promotes cell-cycle arrest and apoptosis, while NH2-terminally truncated forms of p73 are known to have oncogenic potential (34). Binding of RanBPM with p73 significantly enhances the pro-apoptotic activity of p73 by inhibiting its ubiquitination and extending its half-life (34, 35). RanBPM is an active binding partner of macrophage galactose (Gal)-type C-type lectin gene 1 (MGL-1) and promotes MGL-1 tumor suppressor activity by stabilizing MGL-1 and prolonging its half-life through the ubiquitin-proteasomal pathway (36). Previous studies have also shown that RanBPM prevents deregulated expression of some genes such as RON (Recepteur d' Origine Nantais) tyrosine kinase, the adhesion molecule L1 cell adhesion molecule (L1CAM), and transcription factor E74-Like Factor 3 (ELF3) (37). In short, these findings suggest that RanBPM functions as a putative tumor suppressor to modulate cellular events, implicating that it might be involved in cancer development.
However, little is known about the role of RanBPM in malignant diseases. The aim of the current study was to examine the expression of RanBPM in human gastric cancer and determine any association with disease progression and patient survival. Moreover, we also examined the influence of RanBPM on gastric cancer cell functions using in vitro functional assays.
Materials and Methods
Cell lines. Human gastric cancer cell lines AGS and HGC27 were obtained from the European Collection of Cell Cultures (Salisbury, Wiltshire, UK). PC-3, DU-145, LNCaP, CAPHV10, PZHPV7, MDA-MB-231, MCF-7, BT549, BT20, ZR751, A549, SKMES1 and MRC5 were purchased from the American Type Culture Collection (Manassas, VA, USA). PNT-1A and PNT-2C2 were kindly provided by Professor N Maitland, University of York. HECV was obtained from Interlab (Naples, Italy). The cells were maintained in Dulbecco's modified Eagle's medium/F12 (DMEM/F12) supplemented with amphotericin B, penicillin, streptomycin, and 10% foetal calf serum (FCS) (Sigma-Aldrich Inc., Poole, Dorset, UK). The cells were incubated at 37°C with 5% CO2 and 95% humidity.
Gastric tissues. Gastric adenocarcinoma or Siewert type III gastro-oesophageal junction adenocarcinoma tissues (n=245) with matched adjacent normal tissues (n=158) were collected immediately after surgical resection at the Beijing Cancer Hospital with informed consent from the patients. Tissue samples were stored at the Tissue Bank of Peking University Oncology School. Clinical and histopathological information was recorded. All protocols were reviewed and approved by the Beijing Cancer Hospital Research Ethics Committee (MTA10062009).
RNA preparation and reversetranscription–polymerase chain reaction (RT-PCR). Total RNA was isolated from gastric tissues and cells using TRI® Reagent (Sigma-Aldrich Inc.). RNA was converted to cDNA using iScript™ cDNA Synthesis Kit (Bio-Rad Laboratories Ltd., Hercules, CA, USA). Following reverse transcription, PCR was performed using REDTaq® ReadyMix™ PCR Reaction Mix (Sigma-Aldrich Inc.). Primer sequences are listed in Table I. Cycling conditions were preheating at 94°C for 5 min, followed by 30 cycles of 30 sec at 94°C for denaturation, 30 sec at 55°C for annealing and 40 sec at 72°C for elongation, followed by a final 10 min extension at 72°C. The PCR products were separated on a 1% agarose gel stained with Sybr Safe DNA gel stain (Invitrogen Ltd., Paisley, Scotland, UK) and then visualized under ultraviolet light. The quality of DNA was verified and normalized using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as a house-keeping gene.
Quantitative PCR (QPCR). Real-time quantitative PCR was used to determine the level of RanBPM mRNA expression using the prepared cDNA samples. The reaction was carried out in StepOnePlus Real-Time PCR system (Applied Biosystems, Waltham, MA, USA), with the following cycling conditions: 95°C for 10 min, followed by 100 cycles of 95°C for 10 seco, 55°C for 35 sec, and 72°C for 10 sec. GAPDH was used as an internal control to normalize RanBPM gene expression. An internal standard, simultaneously amplified with the samples, was used to generate the levels of the transcripts.
Construction of ribozyme transgenes and knockdown of RanBPM in gastric cancer cells. Hammerhead ribozymes targeted against human RanBPM were designed based on the secondary structure of the mRNA generated using Zuker's RNA Mfold program (38). The ribozymes were synthesized and cloned into pEF6/V5-His-TOPO plasmid vector (Invitrogen Ltd.). The ribozymes were then transfected into AGS and HGC27 cells using a Gene Pulser Xcell™ electroporation system (Bio-Rad). This was followed by selection of successfully transfected cells using selection medium containing 5 μg/ml blasticidin (Sigma-Aldrich Inc.) for up to 2 weeks. Following verification, stable transfectants were cultured in maintenance medium containing 0.5 μg/ml blasticidin. The primer sequences used for the RanBPM ribozymes are shown in Table I.
Primer sequences used in the current study.
Wounding/migration assay. The migration assay was carried out as previously described (39). A total of 3×105 cells were seeded into each well of a 24-well plate. Once the cells reached confluence, the monolayer of cells was scraped to produce a linear wound using a fine-gauge needle. The medium was changed twice to discard any floating cells, and the plate was then returned to the incubator to maintain a constant temperature of 37°C. Cells were photographed every hour for 8 h using a CCD camera attached to a microscope at ×20 magnification. The distance between the edges of the wounding gap was measured using ImageJ software (http://imagej.net/index.html, National Institutes of Health, Bethesda, MD, USA).
Cell-matrix adhesion assay. The cell-matrix adhesion assay was performed as previously described (40). A total of 2×104 cells were added to each well of a 96-well plate which had been pre-coated with 5 μg of Matrigel™ (BD Biosciences, Oxford, UK), air-dried and rehydrated with serum-free medium. After 40 minutes of incubation, non-attached cells were removed by washing with balanced salt solution three times, and adhered cells were fixed with 4% formaldehyde (v/v) for 30 min, prior to staining with 0.5% crystal violet (w/v). The number of adhered cells was counted under a microscope from five random fields per well, and the absorbance of colorimetric products was analysed after solubilisation with 10% acetic acid using a spectrophotometer (ELx800™; Bio-Tek, Winooski, VT, USA) at a wavelength of 540 nm.
In vitro cell-growth assay under normal culture conditions and after exposure to methotrexate (MTX) and fluorouracil (5-FU). Cells (2,500/200 μl) were seeded into each well of a 96-well plate, and treated with MTX or 5-FU at 2.5 μg/ml and 100 μg/ml respectively. Plates were fixed with 4% formaldehyde (v/v) after 1, 3 and 5 days. After staining the cells with 0.5% crystal violet (w/v) and solubilizing with 10% acetic acid, the absorbance was measured using a spectrophotometer (ELx800™; Bio-Tek) at a wavelength of 540 nm.
In vitro invasion assay. Trans-well inserts with 8 μm size pores were coated with 50 μg of Matrigel™ (BD Biosciences) and air-dried. Following rehydration, 2×104 cells were seeded into each insert and incubated for 3 days. The cells that had migrated to the other side of the insert through the matrix were fixed with 4% formaldehyde (v/v), and stained with 0.5% crystal violet (w/v). The absorbance of the colorimetric products was determined at a wavelength of 540 nm using a spectrophotometer (ELx800™; Bio-Tek) after solubilization with 10% acetic acid.
Ran binding protein M (RanBPM) expression in a gastric cancer cohort.
Statistical analysis. Data were analysed using SigmaPlot software (version 12.5; SPSS, Inc., Chicago, IL, USA). The relationship between RanBPM expression and tumor grade, TNM staging and nodal status was assessed using the Mann–Whitney test for non-parametric data. The statistical comparisons between the RanBPM knockdown cell lines (HGC27ΔRanBPM and AGSΔRanBPM) and the control cell line, using cells containing a closed pEF plasmid vector (HGC27pEF and AGSpEF), were made using Student's two sample t-test and two-way ANOVA.
Results
Expression of RanBPM in gastric cancer. We first determined the expression of RanBPM transcript levels in gastric cancer tissues using real-time PCR. RanBPM expression was detected in both gastric cancer tissues and adjacent normal tissues. Slightly increased levels of RanBPM were found in the tumour samples, in comparison with matched background tissues, although this did not reach statistical significance (p=0.33) (Table II). In an examination of cDNA libraries of different cancer cell lines using conventional PCR, the expression of RanBPM was detected in several cancer cell lines, including two gastric cancer cell lines, HGC27 and AGS, seven prostatic cell lines and five breast cancer cell lines, although there were slight differences in their expressions (Figure 1). RanBPM was also expressed by HECV (vascular endothelia) and MRC5 (fibroblast) cell lines, which also exist in the tumours.

Expression of Ran binding protein M (RanBPM) in gastric (A), prostate (B), breast (C), and lung (D) cancer cell lines were detected using reverse transcription polymerase chain reaction.
The level of RanBPM transcript expressed in tumors with distant metastases (M1) was significantly lower than that seen in tumors without distant metastasis (p=0.025) (Table II). In contrast to the reduced expression in tumors with distant metastases, differential expression was seen in tumours with different levels of local invasion. The lowest expression of RanBPM was seen in tumors with invasion confined to the lamina propria, muscularis mucosa or submucosa (T1) compared to its expression in T4 tumors where disease invaded the serosa (p=0.036). Moderate expression levels were seen in tumours invading the muscularis mucosa (Table II).
Creation of gastric cancer cell sub-lines with RanBPM knockdown. The two RanBPM-knockdown gastric cancer cell lines (HGC27ΔRanBPM and AGSΔRanBPM) were created by transfection with anti-RanBPM ribozyme transgenes. After selection with blasticidin (5 μg/ml) for up to two weeks, RanBPM expression was determined using real-time PCR (Figure 2). Reduced RanBPM transcripts were seen in both HGC27ΔRanBPM and AGSΔRanBPM cells that were transfected with anti-RanBPM ribozymes in comparison with the corresponding control cells. The RanBPM-knockdown cells were then used in the subsequent experiments.
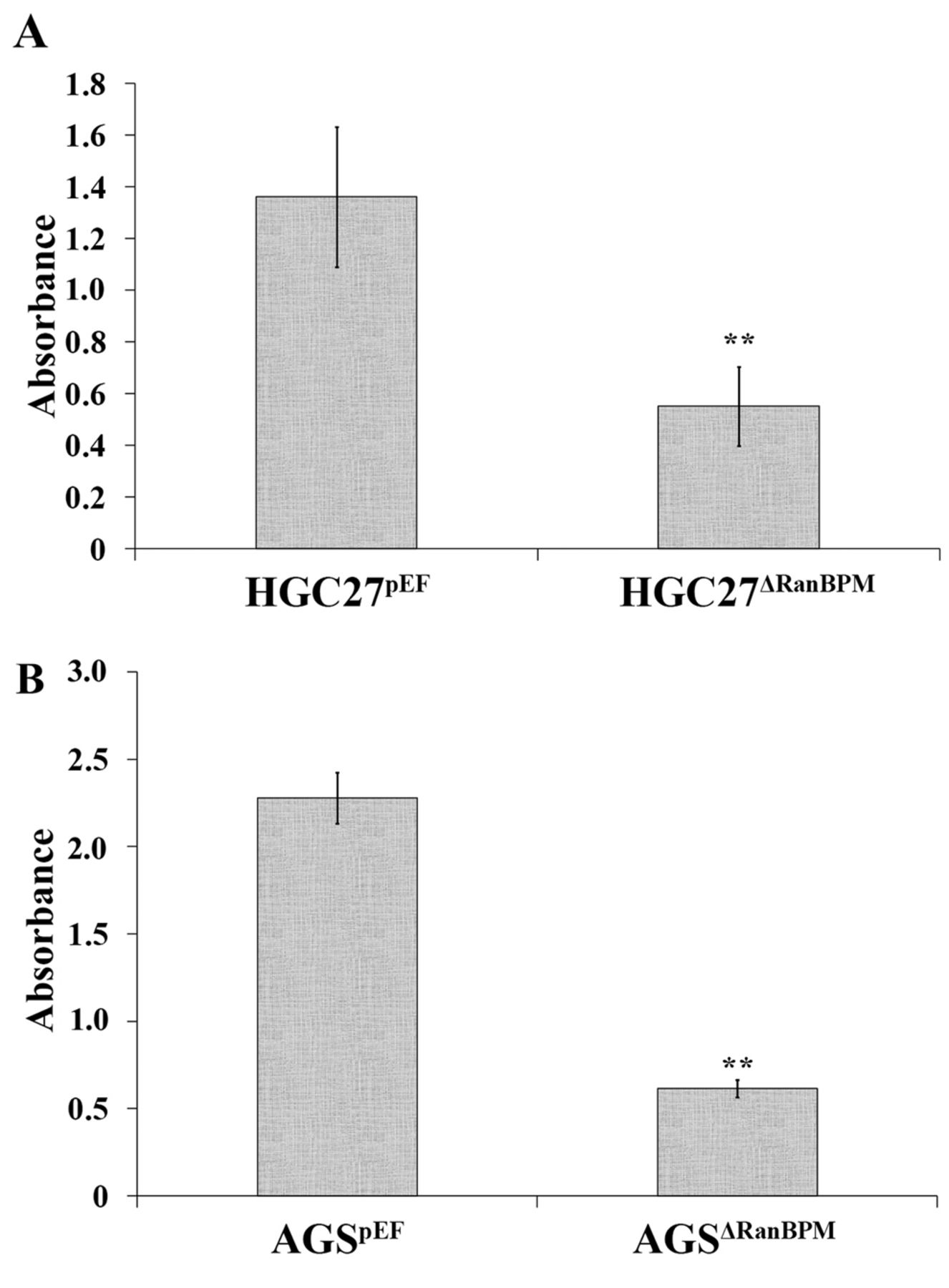
Knockdown of RanBPM reduced gastric cancer cell adhesion. The adhesive ability of HGC27ΔRanBPM and AGSΔRanBPM cells was examined using an in vitro cell-matrix adhesion assay, that assessed the capacity of cancer cells to adhere to an artificial basement membrane. A significant decrease in cell adhesion was seen in both HGC27ΔRanBPM and AGSΔRanBPM cells compared to HGC27pEF cells (p<0.001), and AGSpEF cells (p<0.05) (Figure 3).
RanBPM knockdown promoted cell migration. We investigated the influence of RanBPM on cell migration by measuring the movement of cells after wounding. Knockdown of RanBPM resulted in a significant increase in cell motility in HGC27ΔRanBPM cells compared to HGC27pEF control cells, particularly in the first 6 h after wounding (Figure 4A). RanBPM knockdown in AGS cells did not exhibit such a strong effect on cell migration, although AGSΔRanBPM cells tended to move slightly faster than the AGSpEF control cells (Figure 4B).
Effect of RanBPM knockdown on invasive capacity of gastric cancer cell. We examined the influence of RanBPM on the invasive capacity of both HGC27 and AGS cell lines. Interestingly, knockdown of RanBPM exerted a differential effect on cancer cell invasion between the two gastric cancer cell lines. Reduced cancer cell invasion was seen in HGC27ΔRanBPM cells compared to HGC27pEF (Figure 5A), whilst increased cancer cell invasion was observed in AGSΔRanBPM cells compared to AGSpEF control cells (Figure 5B).

Knockdown of Ran binding protein M (RanBPM) in gastric cancer cell lines. A: Knockdown of RanBPM in HGC27 cells was verified using real-time quantitative polymerase chain reaction. B: Reduced expression of RanBPM was seen in AGSΔRanBPM cells compared to AGS WT and AGSpEF. HGC27 WT and HGC27 pEF: HGC27 wild-type and HGC27 cells transfected with empty plasmids, respectively. HGC27ΔRanBPM: HGC27 cells with RanBPM knockdown. Data are the mean±SD/SEM; n=3; *p<0.05.
Involvement of RanBPM in resistance of gastric cancer cells to chemotherapeutic agents. We investigated the influence of RanBPM on cell proliferation in vitro over a period up to 5 days using the colorimetric method described in the Materials and Methods section. Both HGC27ΔRanBPM and AGSΔRanBPM cells exhibited an increase in in vitro cell proliferation compared to respective control cell lines, but not to a significant level. Selga et al. recently performed a microarray study, analysing differential gene expression in HT29 colon cancer cells sensitive, or resistant, to MTX (41) (data accessible at NCBI GEO database, accession GDS3330). In our analysis of the GEO profile of RanBPM in this database, reduced expression of RanBPM was seen in HT29 MTX-resistant cells compared with MTX-sensitive HT29 cells (p=0.0026) (Figure 6). In order to inquire into the role of RanBPM during chemotherapy, optimization experiments using different concentrations of the two drugs were performed. Appropriate dosages of 5-FU for HGC27 and AGS were identified as 200 μg/ml and 100 μg/ml respectively, and the appropriate concentrations of MTX were 10 μg/ml and 2.5 μg/ml respectively. After exposure (for up to 5 days) to MTX and 5-FU, the survival rates of HGC27ΔRanBPM and AGSΔRanBPM were higher than those of HGC27pEF and AGSpEF control cells. In comparison to MTX, 5-FU was more effective, as reflected in the survival rates of cells. AGSΔRanBPM exhibited significantly higher survival rates compared to the AGSpEF control cells on exposure to MTX (Figure 7D). In contrast, HGC27 cells tended to be less responsive to MTX treatment (Figure 7B). A significant increase in chemoresistance was evident in both HGC27 and AGS RanBPM-knockdown cells when they were treated with 5-FU for 5 days (Figure 7A and C).

Ran binding protein M (RanBPM) promotes cell-matrix adhesion. A: Knockdown of RanBPM (AGSΔRanBPM) reduced adhesion of HGC27 cells. B: A decrease was also seen in AGS with RanBPM knockdown in comparison with cells transfected with empty plasmids (AGSpEF). Blue-dyed cells were AGSpEF adhered to the matrix membrane (C), Blue-dyed cells were AGSΔRanBPM adhered to matrix membrane (D). Data are the mean±SD/SEM; n=6; **p<0.01.

Ran binding protein M (RanBPM) reduces cell migration. A: HGC27ΔRanBPM knockdown cells moved farther in comparison with HGC27 cells transfected with empty plasmid (HGC27pEF) as control after 8 h. B: Increased migration was also seen in AGS cells with RanBPM knockdown (AGSΔRanBPM) compared to AGSpEF. **p<0.01 vs. pEF control. Data are the mean±SD/SEM; n=3.

Ran binding protein M (RanBPM) affects invasion of gastric cancer cells. Knockdown of RanBPM (ΔRanBPM) results in reduced invasiveness of HGC27 cells, whereas an increase was seen in AGS cells. Data are the mean±SD/SEM; n=3; *p<0.05.

Expression of Ran binding protein M (RanBPM) in HT29 methotrexate (MTX)-resistant cells. Selga et al., recently performed a microarray analysing differential gene expression in HT29 colon cancer cells sensitive, or resistant, to MTX (data accessible at NCBI GEO database, accession GDS3330).
Discussion
In the present study, we examined the expression of RanBPM in both gastric cancer tissues and gastric cancer cell lines. In the tissue samples obtained from patients, we identified elevated expression of RanBPM in gastric tumor tissue though this was not statistically different to the levels of adjacent background gastric tissues. Expression of RanBPM was also detected in gastric cancer cell lines (HGC27 and AGS). Screening of RanBPM mRNA expression in other cell lines showed that RanBPM was also ubiquitously expressed by other cancer cell lines and non-cancer cell lines (HECV and MRC5). These results suggest that the expression level of RanBPM in tissues may be heterogeneous. Further investigations using fluorescence in situ hybridization or immunohistochemical staining may help clarify the expression of RanBPM and determine its distribution in tissues and cells.

Ran binding protein M (RanBPM) is involved in chemotherapy resistance in gastric cancer cells. A: HGC27 cells with RanBPM knockdown (ΔRanBPM) showed stronger tolerance to fluorouracil (5-FU) (200 μg/ml) when treated for 3 days and 5 days. B: Treatment with 10 μg/ml methotrexate (MTX) for 3 days and 5 days revealed higher survival of HGC27ΔRanBPM cells in comparison to HGC27pEF as control. C, D: Higher survival rate was also seen in AGSΔRanBPM cells when treated with 100 μg/ml 5-FU and 2.5 μg/ml MTX for 3 and 5 days respectively. *p<0.05 vs. Day 1 control; **p<0.01 vs. Day 1 control; #p<0.05 vs. pEF control; ##p<0.01 vs. pEF control. Data are the mean±SEM; n=6.
Aberrant expression of RanBPM was seen in tumours with different levels of local invasion. For example, the most locally invasive tumours (T4) expressed higher levels of RanBPM compared to T1 tumours. This suggests that RanBPM may play a promotory role during local invasive growth of gastric cancer. In contrast to its expression in tumors with different degrees of local invasion, the expression of RanBPM in tumors with distant metastases, namely M1 tumors, was significantly lower compared to levels seen in tumours with no distant metastases. This suggests that a different role may be played by RanBPM in distant metastases of gastric cancer, that is in line with findings by Jun-Dong Wei et al. showing that RanBPM acts as a negative regulator of cell motility (42). However, its exact impact on cellular functions and cell–cell interactions during the metastatic process is yet to be investigated.
RanBPM has been shown to act as a scaffolding protein, assisting signal transduction and protein–protein interactions. For example, the role played by RanBPM in the semaphorin3A/plexinA1 pathway and the interaction between amyloid precursor protein and lipoprotein receptor-related protein (29, 43). In the current study, we examined the impact of RanBPM knockdown on the cellular functions of gastric cancer cells. RanBPM knockdown exerted differential effects on adhesion, motility, invasion and growth of HGC27 and AGS cells. Knockdown of RanBPM impaired cell-matrix adhesion of gastric cancer cells. It has been reported that intercellular adhesion molecule-1, integrins, and other adhesion-related proteins are essential in cell adhesion, which mediates the critical response of cells to the surrounding environment (44). Therefore, further investigation is required to elucidate how RanBPM is involved in the coordination of cell adhesion through interactions with these adhesion molecules. RanBPM can regulate cell migration by coordinating relevant signalling via transmembrane receptors (36). On the other hand, the results of our wounding assay show that knockdown of RanBPM in gastric cancer cells enhanced their motility. This indicates that RanBPM may be a mediator involved in inhibiting gastric cancer cell migration. A number of molecules, such as 3-phosphoinositide-dependent protein kinase-1, have been demonstrated to participate in signal transductions that regulate cell migration (45, 46). In addition, RanBPM may play an important role in the signal transduction of different pathways, including extracellular signal-regulated kinase1/2, wingless-related integration site, NOTCH, and phosphatidylinositol-4,5-bisphosphate 3-kinase/ Protein kinase B (Akt) pathways (37). The RanBPM knockdown cells not only became less adhesive, but also displayed enhanced motility to a certain degree. However, the involvement of the aforementioned pathways is yet to be clarified.
We further focused on cancer cell invasion of RanBPM-knockdown cells. Interestingly, with knockdown of RanBPM, HGC27 cells appeared to be less invasive whilst AGS knockdown cells exhibited enhanced invasiveness. This suggests that different interacting partners of RanBPM may exist in the two gastric cancer cell lines, leading to differential responses in their invasiveness. Further investigation into the underlying mechanism is yet to be carried out.
Although knockdown of RanBPM in gastric cancer cells did not remarkably affect their in vitro growth, enhanced survival was seen in both HGC27 and AGS RanBPM knockdown cells when they were exposed to chemotherapeutic drugs. RanBPM has been demonstrated to be an activator of apoptotic pathways induced by DNA damage (33). Together with the findings from the current study, it is suggested that RanBPM plays a role in differential responses to certain chemotherapeutic agents and consequently influences tumour chemoresistance. Further investigation will help highlight the potential application of RanBPM in personalized disease management, for example in the selection of appropriate chemotherapeutic agents.
Acknowledgements
The Authors wish to thank Cancer Research Wales and Albert Hung Foundation for their kind support. Miss Shuai Shao is a recipient of the Chinese Medical Research Studentship of Cardiff University.
Footnotes
↵* Current address: Capital Medical University, 10 Xitoutiao, Feng Tai District, Beijing 100069, China.
- Received January 18, 2016.
- Revision received February 19, 2016.
- Accepted February 22, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}