


Abstract
Background/Aim: Dendritic cell (DC) immunotherapy induces tumor-reactive T-cells. We optimized the maturation of murine DC against ovarian cancer. Materials and Methods: Immature DC were generated from bone-marrow progenitor cells and loaded with hypericin-photodynamic-treated (Hyp-PDT) tumor cells (primary maturation). Lipopolysacharide (LPS 1 μg/ml) was used as a secondary maturation stimulus. After 24 h, maturation was assessed using flow cytometry. For in vivo experiments, C57BL/6 mice were vaccinated subcutaneously with matured, loaded mature DC. Immune response was evaluated through immunohistochemistry and cytometric bead array. Results: Based on the existing protocols for DC generation, therapeutic vaccination of tumor-bearing mice with mature ID8-fLuc Hyp-PDT DC induces an immunogenic response, but provides no survival benefit. As loading with Hyp-PDT lysate induces partial primary maturation, we reduced the dose of LPS to 0.125 μg/ml and the duration of maturation to 6 hours, improving viability of mature DC. Conclusion: We optimized Hyp-PDT-based ID8-fLuc DC vaccine by reducing the amount of LPS and the duration of maturation.
Ovarian cancer is a silent killer. Today, four out of five women diagnosed with advanced-stage high-grade serous ovarian cancer die within 5 years of initial diagnosis, despite effective chemotherapies and radical surgery (1-3). New treatment strategies are desperately needed.
Interest in immunotherapy for the treatment of cancer has been increasing (4). Dendritic cells (DC) are a safe and effective method for inducing anti-tumoral cytotoxic T-cells. Clinical response rates from human DC vaccination trials in ovarian cancer remain, however, limited (5, 6). There are several possible strategies for improving the efficacy. On one hand, the DC vaccine itself can be optimized to improve the induction of cluster of differentiation 8+ (CD) T-cells and increase the interferon-γ (INF-γ) response (7, 8). On the other hand, the immunosuppressive tumor-microenvironment appears to be an important hurdle to an effective immune response (9, 10).
At present, there is no consensus on the optimal method for culturing DC. One way to improve the power of DC is to modify their antigen loading. Hypericin-based photodynamic therapy (Hyp-PDT) is a potent method for inducing type II immunogenic cell death (11). Treatment of tumor cells with Hyp-PDT induces several important damage-associated molecular patterns such as expression of surface calreticulin, surface heat-shock protein 70 (HSP70), surface HSP90, secretion of adenosine triphosphate (ATP) and release of high-mobility group 1, HSP70, HSP90 and calreticulin (11, 12). Loading DC with Hyp-PDT-treated tumor cell-lysate leads to an increase in interleukin-6 (IL-6), IL-1β production by DC and an expansion of INF-γ-producing T-cells (13-15). Another method for improving the function of DC vaccine in vivo is to alter the DC maturation process.
In this article, we describe the search for an improved strategy for producing immunogenic mature DC (DCm), loaded with Hyp-PDT-treated tumor cell-lysate, based on the existing protocols in a model of high-grade serous ovarian cancer with peritoneal spread (16-19). We focus on loading and maturation of immature DC (DCi), yielding more potent DCm.
Materials and Methods
Mice. Six- to eight-week-old C57BL/6, C57BL/6/BrDCHsd-Tyrc and C57BL/6J-Tyrc-2J/J mice were used. C57BL/6J-Tyrc-2J/J were bred in our own colony derived from a breeding pair purchased via Charles River from The Jackson Laboratory (ME, USA). C57BL/6, C57BL/6/BrDCHsd-Tyrc mice were purchased from Harlan, the Netherlands. For the in vivo experiment, only female mice were used. All animals were housed and treated according to the Federation for Laboratory Animal Science Associations guidelines (20). Ethical approval was obtained from the local Ethical Committee (p075/2014).
Preparation of whole tumor cell lysate. In all experiments, ID8 cells stably transduced with firefly luciferase (ID8-fLuc) cells were used to generate lysate. ID8-fLuc cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with Penicillin-Streptomycin (100 IU/ml, Lonza), Gentamicine (50 μg/ml), L-glutamine (2 mM) (all Lonza, Basel, Switzerland) and 10% fetal bovine serum. Cells were cultured in T75 adherent tissue cultures flasks (Nunc, Thermo Scientific, Waltham, MA, USA). For preparation of immunogenic lysates, the cells were first detached by addition of trypsin-EDTA (200 mg/l; Lonza) and snap-frozen in liquid nitrogen. Thereafter the cells were thawed, suspended in Dulbecco's phosphate-buffered saline (DPBS, Lonza) and exposed to Hyp-PDT treatment. For Hyp-PDT based elicitation of immunogenic cell death, the cells were incubated with 1 μM hypericin [prepared, purified and stored as detailed elsewhere (12)] for 2 h in DPBS followed by light irradiation (4.05 J/cm2) performed as described previously (8, 12, 20). Cells were recovered 4 h post-treatment. Thereafter the cells were exposed to three cycles of freeze-thawing to ensure 100% devitalization of cells i.e. no live cells within the lysate (8).
Isolation of bone marrow-derived progenitor cells. Murine DC were produced based on existing protocols (16-18). The mice were euthanized by cervical dislocation. Subsequently the lower part of the mouse was disinfected with ethanol 70%. Tibia and femur of both hind legs of the mouse were isolated by sharp dissection. The remaining soft tissue was removed by sterile gauze and the bone was placed in clean DPBS. For disinfection, the bones were placed in ethanol 70% for 30 s and then again in DPBS. Next the ends of the bones were cut away at the epiphysis and bone marrow was flushed from the bone using DPBS and collected into a Falcon tube (Cell Star, Greiner bio-one, Stonehouse, UK). The cells were then cultured in the presence of recombinant murine granulocyte-macrophage colony-stimulating factor (rmGM-CSF 20 ng/ml; PeproTech, Rocky Hill, USA) at a maximum density of 750×103 progenitor cells/ml in Roswell Park Memorial Institute medium (RPMI; Lonza) augmented with Penicillin-Streptomycin (100 IU/ml; Lonza), β-mercaptoethanol (50 mM; Sigma-Aldrich, Diegem, Belgium), L-glutamine (2 mM; Lonza) and 10% fetal bovine serum (Gibco, Thermo Scientific). After 3 days, fresh medium was added up to a volume of 150% together with rmGM-CSF (20 ng/ml). After 5 days, 50% of the total medium was removed, centrifuged at 400 × g for 5 minutes and cells were resuspended in fresh medium. At this stage, the culture was again supplemented with rmGM-CSF (20 ng/ml). At day 7, DCi were harvested by gentle dislocation with a cell scraper and co-incubated for 90 min with thawed lysate (150 μg protein/106 DCi, 1×106 DCi/100 μl DC medium) previously sonicated for 15 min and heavily vortexed in order to obtain a homogeneous suspension.
Fluorescence-activated cell sorting (FACS). DC maturation markers were assessed using flow cytometry. First cells were incubated for 5-10 min with 0.5 μl of fragment, crystallizable receptor blocker (Fc block) (Anti-Mouse CD16/CD32 clone 93; eBioscience, Vienna, Austria) to avoid non-specific binding. Next DC were incubated at 4°C for 20-30 min with phycoerythrin–cyanine 7 (PE-Cy7)-conjugated monoclonal antibodies to CD40 (clone 3/23; BioLegend, San Diego, CA, USA) CD86 (GL1; BD Bioscience, Erembodegem, Belgium) major histocompatilibity complex-II molecule (MHCII/IA-IE) (clone M5/114.15.2; eBioscience) CD80 (clone 16-10A1; eBioscience) major histocompatilibity complex-I molecule (MHCI/H2-Kb) (clone AF6-88.5.5.3; eBioscience) and allophycocyanin(APC)-conjugated monoclonal antibody to CD11c (clone N418 eBioscience). DC were incubated with PE-Cy7-conjugated antibody to CD197 [C-C chemokine receptor type 7 (CCR7)] (clone 4B12 eBioscience) at 37°C. Zombie Yellow (BioLegend, San Diego, CA, USA) was added as a viability dye. Acquisition was performed using Cellquest software on a LSR Fortessa FACSort cytometer (BD Bioscience). Analysis was performed using FlowJo Single Cell Analysis software (TreeStar, Ashland, OR, USA).
In vivo vaccination. Mice were intraperitoneally inoculated with 5×106 ID8-fLuc cells and vaccinated subcutaneously 21, 28 and 35 days later with 1×106 DCm loaded with ID8-fLuc Hyp-PDT lysate. For the survival experiment, loaded DC were matured with 1 μg/ml LPS for 24 h. In the immunomonitoring experiments, loaded DC were matured with 0.5 μg/ml LPS for 18 h. In order to allow correct evaluation of survival, mice were repeatedly drained of ascites when they weighed 32 g, as described before (19).
Cytokine analysis. Cytometric bead array (CBA; BD Bioscience) was used for the determination of the cytokine content of supernatants from ascites. We tested for IL-1β, IL-10, IL-12p70, monocyte chemo-attractant protein1 (MCP1/CCL2), tumor necrosis factor (TNF), IL-17a, IL-6 and INF-γ. The procedure was performed in accordance with the manufacturer's protocol. Acquisition was performed using Cellquest software on a LSR Fortessa FACSort cytometer (BD Bioscience). Analysis was performed using FCAP Array Software 3.0 (BD Bioscience).
Immunohistochemistry. Tumor tissue on the peritoneum and omentum was sampled and paraffin-embedded. Slides were permeabilized and stained with a primary antibody for CD8a (BioXcell, West Lebanon, NH, USA). Stained sections were then incubated with a horseradish peroxidase-labeled secondary antibody. Sections were counterstained with Mayer's hematoxylin solution and scored subjectively for intensity on three representative tumor samples (22).
Statistical analysis. Statistical analysis was performed using GraphPad Prism 6 (GraphPad Software, Inc., San Diego, CA, USA). Student's t-tests were used; significance level was set at p<0.05.
Results
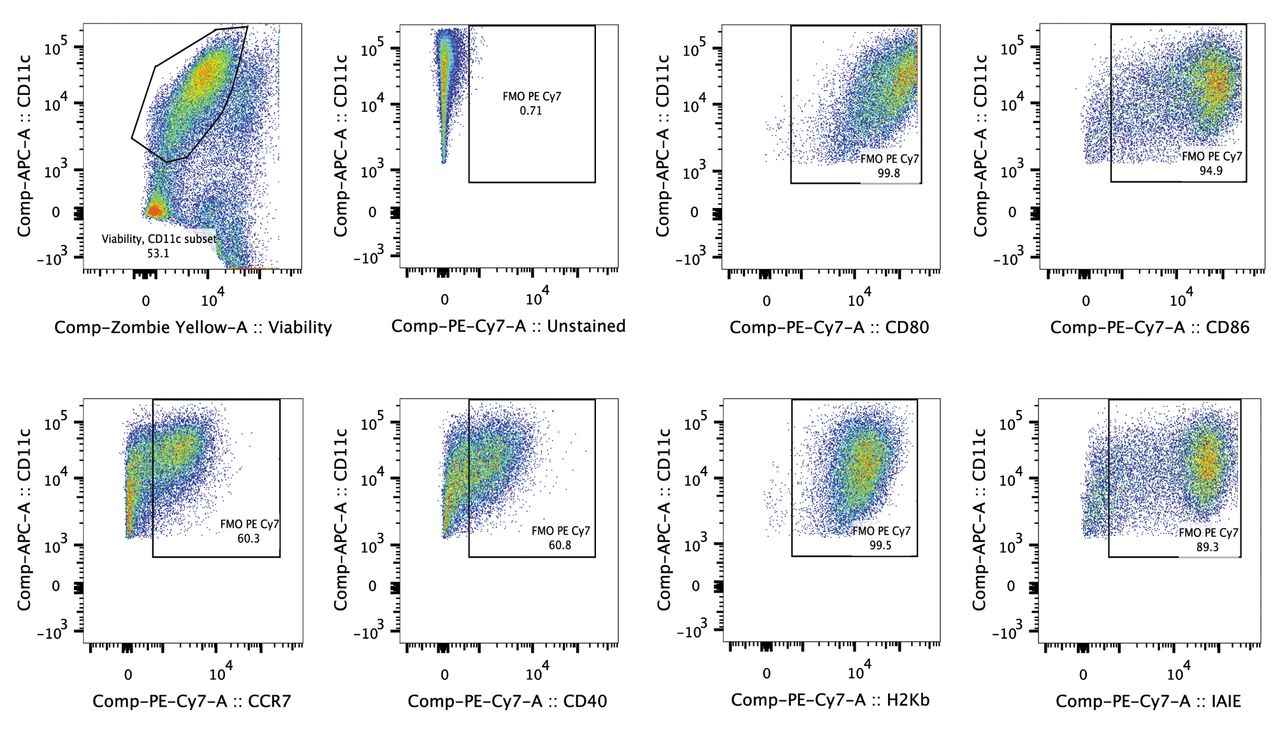
DC loaded with ID8-fLuc Hyp-PDT lysate are highly auto-fluorescent. Mature DC are large cells (10-15 μm) with cytoplasmic extensions (23). Cultured DC are highly auto-fluorescent, which makes correct quantification of markers difficult (24). Abundantly expressed maturation markers are especially difficult to quantify in channels of strong fluorochromes (such as PE), as large quantities of background fluorescence are observed in these channels. By evaluating the auto-fluorescence of ID8-fLuc Hyp-PDT DCm, we observed that Alexa-Fluor 700 (AF700), allophycocyanin-cyanine 7 (APC-Cy7) and PE-Cy7 were suitable fluorochromes for the determination of the level of expression of maturation markers on ID8-fLuc Hyp-PDT DCm (Figure 1). Based on these results, we chose PE-Cy7 as a fluorochrome for the maturation markers in our panel. Zombie Yellow was used as a cell death/permeabilization marker (Table I). Figure 2 shows the gating strategy used to quantify phenotypic maturation.

Evaluation of auto-fluorescence on ID8-fLuc hypericin photodynamic therapy-treated mature dendritic cells (Hyp-PDT DCm). Cells were acquired with all lasers and detectors of the LSR Fortessa FACSort cytometer activated. Auto-fluorescence is limited using Alexa Fluor 700 (AF700), allophycocyanin-cyanine 7 (APC-Cy7) and phycoerythrin–cyanine 7 (PE-Cy7). FITC: Fluorescein isothiocyanate; PerCP-Cy5.5: peridinin chlorophyll-cyanine 5.5; APC: allophycocyanin.

Exemplary gating strategy for murine dendritic cells (DC). Firstly the doublets are excluded. Next, live cluster of differentiation 11c (CD11c) positive cells are selected. Expression of maturation markers on live CD11c-positive cells is determined using a fluorochrome minus one control (FMO).
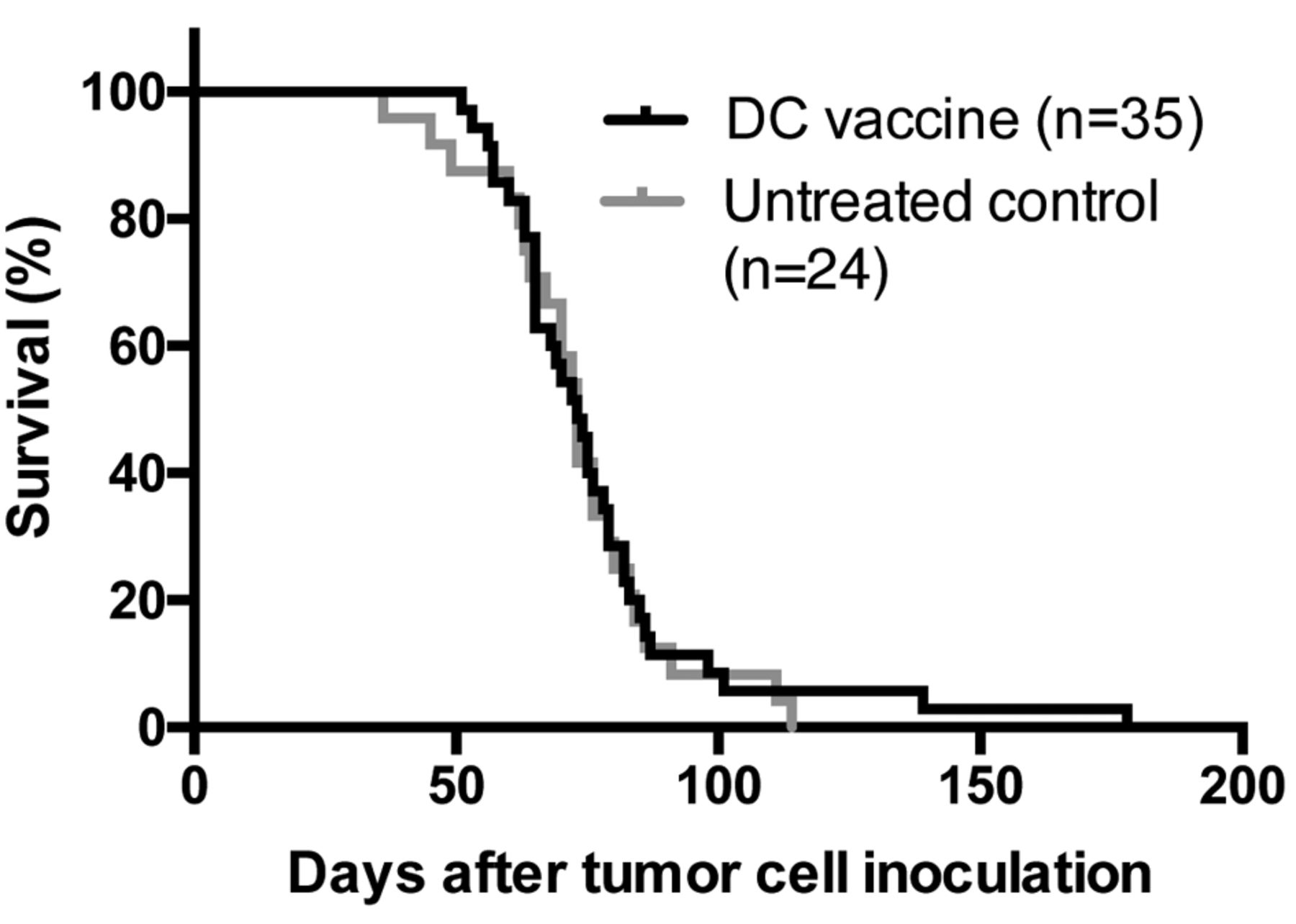
Functionality of cultured DC. We determined whether the ID8-fLuc Hyp-PDT DCm vaccine could improve overall survival in a mouse model of advanced ovarian cancer. Survival of treated animals remained very similar to that of the untreated controls, with a median survival of 73 days in both groups as depicted in Figure 3. In total, 6% (n=2/35) of DC-treated mice were long-term survivors, determined as being outliers by ROUT testing (Q=1%).
Evaluation of in vivo immunogenic response after vaccination. Next we evaluated whether the ID8-fLuc Hyp-PDT DC vaccine led to an immune response even when there was no survival benefit for the mice. For this experiment, mice were sacrificed after the second or third vaccine. Peritoneal fluid was collected for the determination of cytokines and immunohistochemistry was performed on tumor tissue. The treated animals showed significantly higher concentrations of IL-12p70, a potent immunostimulatory cytokine, after the third vaccination. INF-γ and IL-6 were not significantly increased in ascites after vaccination in this experiment (Figure 4A and B). Furthermore, we observed an influx of CD8+ cells into the tumor after the second vaccination, which was less pronounced after the third vaccine (Figure 4C and D).

Survival of tumor-bearing mice in a Kaplan–Meier curve. We compared mice that received ID8-fLuc hypericin photodynamic therapy-treated dendritic cell (Hyp-PDT DC) immunotherapy compared to untreated controls. There was no significant difference between treated or untreated animals. (Results from five pooled experiments).

Evaluation of immune response after vaccination. A and B: Cytokine analysis by cytometric bead array (CBA) of ascites after three ID8-fLuc hypericin photodynamic therapy-treated dendritic cell (Hyp-PDT DC) vaccines. In the DC-treated group we observed significantly higher levels of interleukin-12p70 (IL-12p70) compared to the untreated controls (A). Levels of IL1-b, IL-10, tumor necrosis factor (TNF) and interferon-γ (IFNγ) were slightly increased after ID8-fLuc Hyp-PDT DC vaccination, results were however not significant (B). C and D: Immunohistochemistry for cluster of differentiation 8 (CD8) showed CD8 infiltration in a peritoneal metastasis before start of vaccination C) and intense CD8 staining in tumor tissue after the second ID8-fLuc Hyp-PDT DC vaccine (D) (magnification ×64).
Loading DCi with ID8-fLuc Hyp-PDT lysate induces partial maturation. We examined the ability of ID8-fLuc Hyp-PDT lysate to induce maturation of murine DCi. For this experiment, DCi were co-cultured with ID8-fLuc Hyp-PDT lysate and stimulated with 1 μg/ml LPS, a commonly used maturation stimulus. Figure 5 clearly shows that Hyp-PDT lysate induces maturation, but that addition of LPS is necessary to increase mainly CD40, CD86 and IA/IE expression.
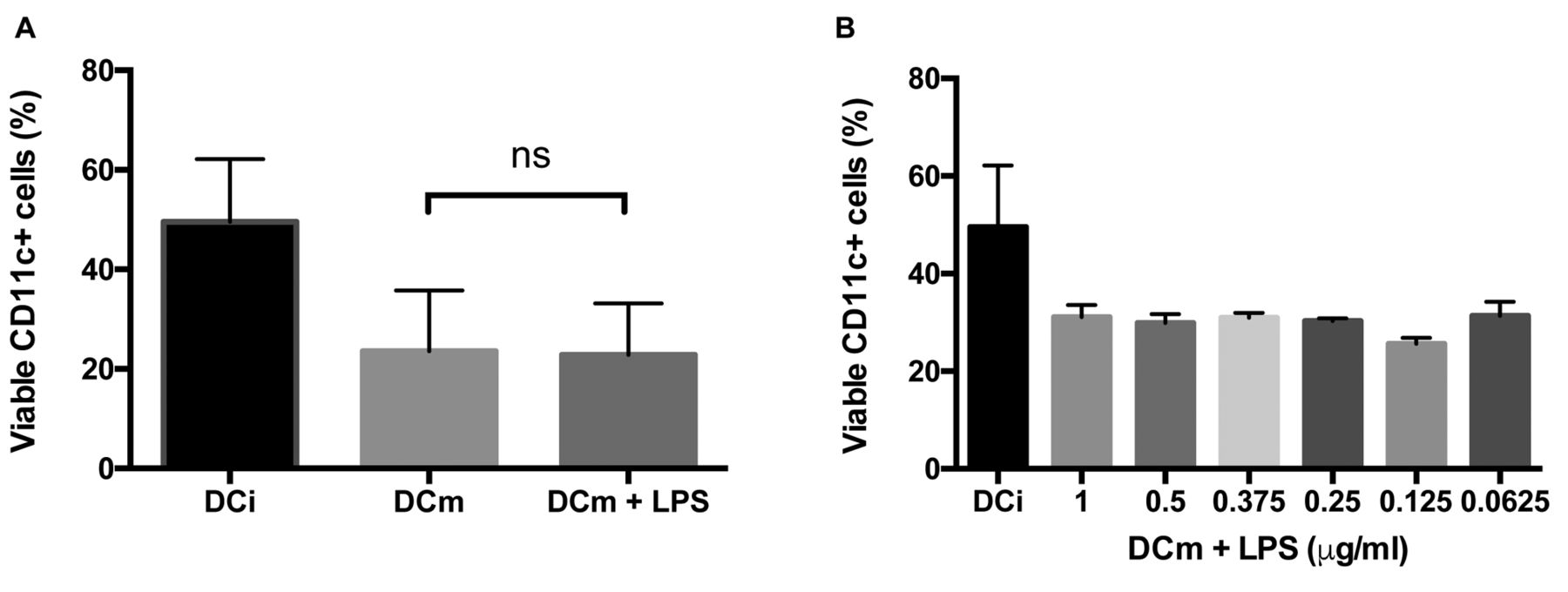
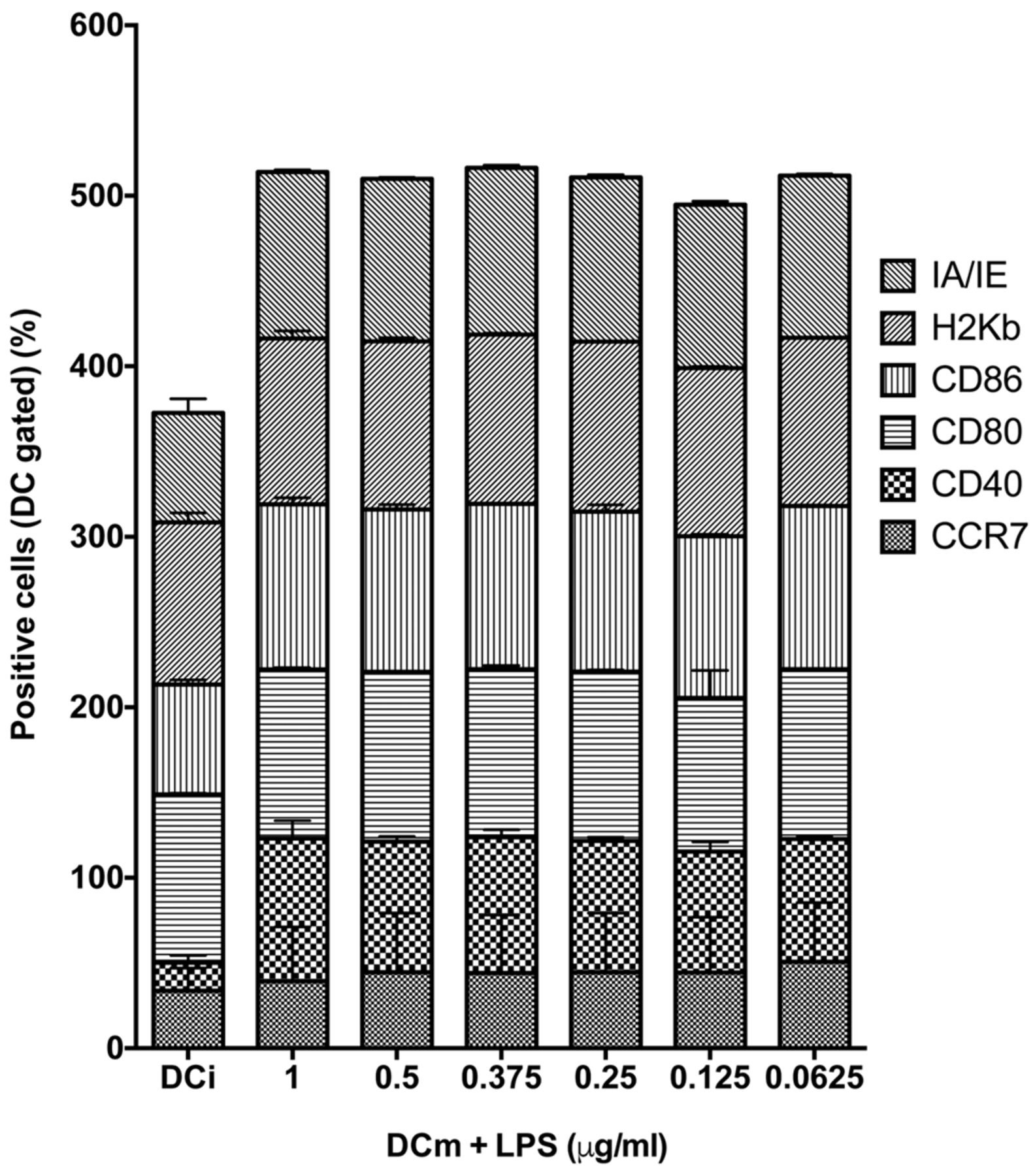
Maturation can lead to DC exhaustion. However, maturation through administration of LPS leads to poor survival of DC, due to exhaustion. We tested if a reduction in the administered dose of LPS would improve survival without impairing DC maturation (Figure 6). Therefore, we administered progressively lower doses of LPS to DCi loaded with ID8-fLuc Hyp-PDT lysate and quantified the maturation markers. We observed that even the lowest concentration of LPS tested (0.0625 μg/ml) was was able to induce optimal maturation (Figure 7). The viability of DC, however, did not improve with use of a lower dose of LPS.
Duration of DC maturation. In order to prevent exhaustion, the length of DC maturation using LPS was changed. Figure 8A shows an important decrease of DCm viability over time. After 6 h of maturation, nearly full phenotypic maturation of the DC was apparent (Figure 8B).
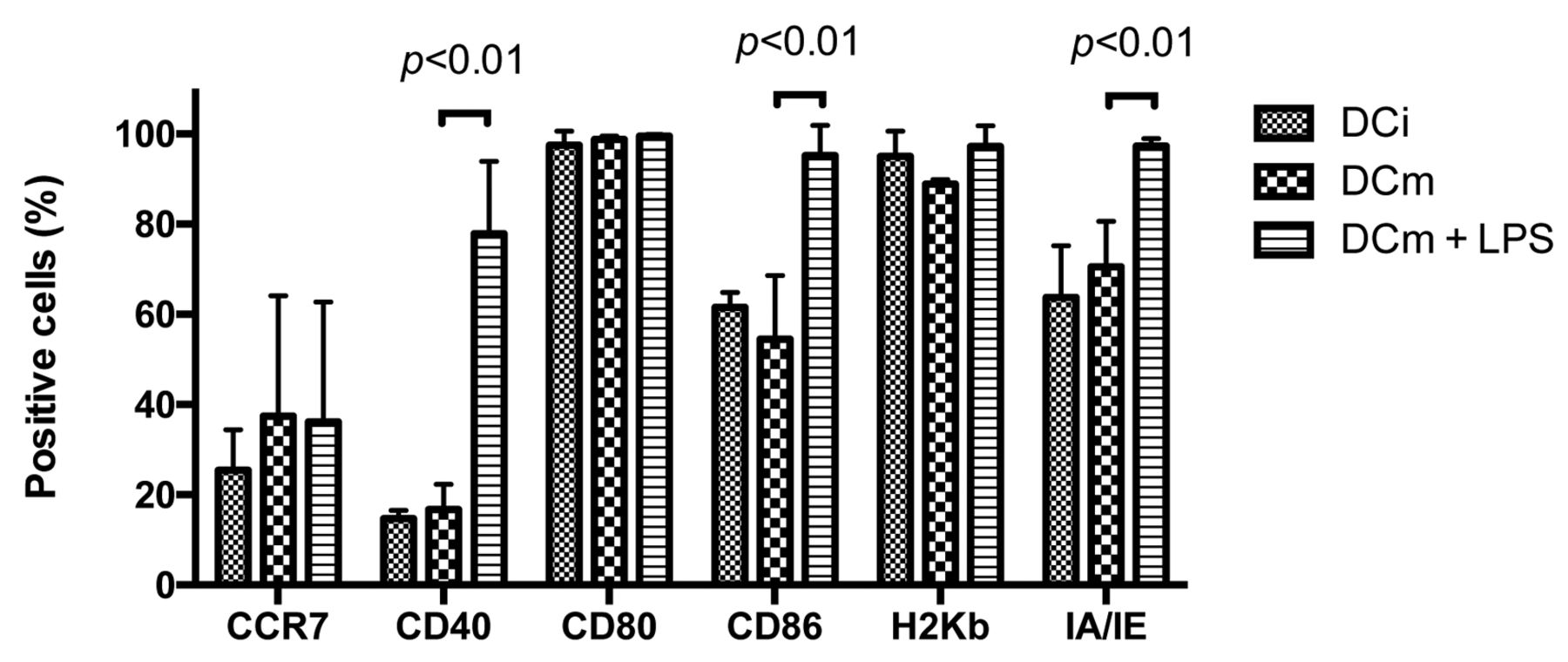
Dendritic cell (DC) maturation is dependent on stimulation by lipopolysaccharide (LPS). We determined the presence of the most important maturation markers through fluorescence-activated cell sorting (FACS) analysis. We observed a significant difference in expression of cluster of differentiation 40 (CD40), CD86 and major histocompatilibity complex-II molecule (MHCII/IA-IE) when we administered LPS at a concentration of 1 μg/ml.

Viability of dendritic cells (DC) measured by fluorescence-activated cell sorting (FACS) analysis. All DC were loaded with ID8-fLuc hypericin photodynamic therapy-treated dendritic cell (Hyp-PDT DC) lysate. A: We compared the percentage of viable immature Dc (DCi), ID8-fLuc Hyp-PDT-loaded mature DC (DCm) and ID8-fLuc Hyp-PDT-loaded DCm cultured in the presence of 1 μg/ml lipopolysaccharide (LPS) (DCm+LPS). There was no significant difference in viable cluster of differentiation 11c+positive (CD11c) cells between the DCm group and DCm+LPS group. B: We evaluated the viability of loaded DC matured with different concentrations of LPS. We observed no significant difference in viable CD11c+ cells between DCm groups.
Maturation markers used to stain murine dendritic cells.

Dendritic cell (DC) maturation was not significantly influenced by lowering the concentration of lipopolysaccharide (LPS). We determined the maturation markers of ID8-fLuc hypericin photodynamic therapy-treated dendritic cell (Hyp-PDT DC) lysate-loaded DCm. There were no significant differences between the different concentrations of LPS. MHCII/IA-IE: Major histocompatilibity complex-II molecule; MHCI/H2-Kb: major histocompatilibity complex-I molecule; CD: cluster of differentiation; CCR7/CD197: C-C chemokine receptor 7.
Discussion
Murine DC cultures are widely used in preclinical immunotherapy research and different methodologies have been proposed but no consensus has yet been reached. This study proposes a protocol for DC generation that is adapted to a strong immunogenic lysate (Hyp-PDT). We also show that ID8-fLuc Hyp-PDT DC immunotherapy, when administered subcutaneously, can yield an immunological response after therapeutic vaccination in an orthotopic model of advanced ovarian cancer, however, without survival benefit. This finding is an important stepping stone on which to build an effective combination strategy of DC immunotherapy with other immunomodulatory agents, such as checkpoint inhibitors, chemotherapeutics, Toll-like receptor-agonists (TLR), etc. Hyp-PDT DC immunotherapy has been shown to be efficient in a therapeutic setting in a model of high-grade glioma, when sequentially combining vaccination with temozolomide-based tumor regression (25).
Although loading of DCi with ID8-fLuc Hyp-PDT lysate induces a degree of maturation, an additional stimulus was needed to achieve full DC maturation. We were able to reduce both the dose of administered LPS and the duration of stimulation. Merely reducing the dose of LPS did not improve viability of DCm. Reducing the LPS exposure time improved viability of DC. Langenkamp et al. already showed that cytokine (IL-6, IL-12, IL-10 and tumor necrosis factor-α) expression of LPS-stimulated DC was abrogated after 24 h of incubation. Therefore we chose to use 0.125 μg/ml LPS as a secondary maturation stimulus and maturation duration of 6 h. By this optimization, we hope to avoid exhaustion of DCm in vivo and achieve a stronger antitumoral effect.
Currently, we continue to use LPS, which is a TLR4 agonist, as a maturation stimulus. Other studies suggest that the combination of R-848 (TLR7/8 agonist) in combination with LPS or poly(I:C) (TLR3 agonist) (26–28) can lead to a more suitable microenvironment to create antitumoral cytotoxic T-cells. We will investigate this further in the near future.
Up till now, in vivo testing showed no improvement in survival of the vaccinated mice. This can probably be explained by two factors. Firstly, this is a model of the peritoneal spread of ovarian cancer. Thus, it skips the ‘natural’ transition from primary ovarian tumor (at the ovary) to metastatic ovarian tumor and starts directly with peritoneal implants. This selective modeling may create a more immunotherapy-resistant version of tumors that did not undergo normal immunosurveillance and thus have a more deranged immuno-microenvironment (29). Secondly, it was recently demonstrated (through AY27 rat bladder carcinoma model) that some cancer models may exhibit resistance to immunogenic danger signaling or immunogenic phagocytosis (12) thereby making vaccines based on these cells less effective in vivo. The former shortcoming could be corrected by ‘resetting’ the tumor microenvironment either by other immunotherapy or through suitable chemotherapy (11). The latter could be corrected by exogenously adding crucial danger signals to increase immunogenic potential (15). Hence, currently we are looking into possible combination strategies to reset the immune system before using our ID8-fLuc PDT DC vaccine.
To conclude, for the maturation of DCi loaded with the strongly immunogenic Hyp-PDT lysate we propose use of 0.125 μg/ml LPS as a secondary maturation stimulus and maturation duration of just 6 h. Further investigations are needed to clarify the in vivo effect of such optimization.
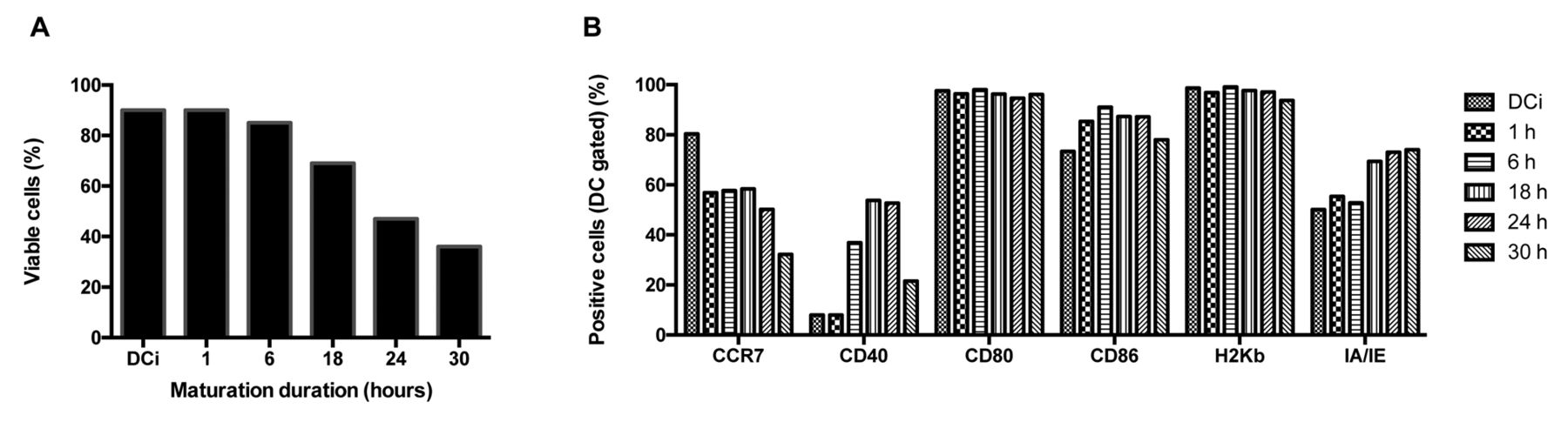
Influence of duration of dendritic cell (DC) maturation on viability and phenotypic maturation. In this experiment, we harvested ID8-fLuc hypericin photodynamic therapy-treated (Hyp-PDT)-loaded DC maturated with 0,125 μg/ml lipopolysaccharide (LPS) at different time-points. A: Over time, viability of mature dendritic cells (DCm) gradually decreased. B: Phenotypic maturation of DC during LPS stimulation. Cluster of differentiation 40 (CD40) up-regulation is present from 6 hours onwards. MHCII/IA-IE: Major histocompatilibity complex-II molecule; MHCI/H2-Kb: major histocompatilibity complex-I molecule; CD: cluster of differentiation; CCR7/CD197: C-C chemokine receptor 7.
Acknowledgements
The Authors would like to thank the Olivia Hendrickx Research Fund for their support. A. Coosemans and A.D. Garg are supported by the Fund for Scientific Research Flanders (FWO).
Footnotes
This article is freely accessible online.
- Received August 22, 2016.
- Revision received September 20, 2016.
- Accepted September 22, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}