


Abstract
Background/Aim: Secreted phosphoprotein 24 kD (spp24) has been shown to inhibit bone morphogenetic protein 2 (BMP2)-induced cancer growth in several tumor models. In this study, we aimed to investigate the effects spp24 on the growth of prostate cancer caused by BMP2 in vitro and in vivo. Materials and Methods: The effects of BMP2 and spp24 on PC-3 cell viability were analyzed using MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) assay. A subcutaneous tumor model and intratibial tumor model was established using PC-3 cells. Tumor growth was assessed through gross examination and radiography during the experiment. Then, after sacrifice, tumor cell apoptosis and tumor cell proliferation were assessed by terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay and immunochemical analysis. Results: BMP2 stimulated the PC-3 cell proliferation in vitro and spp24 could abolish the effect of BMP2. In a xeneograft tumor model, BMP2 promoted the subcutaneous and intratibial tumor growth, while spp24 dramatically inhibited the tumor growth induced by BMP2. Histological examination showed that spp24 also abolished the BMP2-induced proliferating cell nuclear antigen (PCNA) expression and promoted tumor cell apoptosis. Conclusion: Spp24 can inhibit the growth of prostate cancer and its bone metastasis induced by BMP2; spp24 may have great potential to be a therapeutic agent in clinical situations.
Prostate cancer is one of the most common cancers in elderly men (1). Prostate cancer may account for 10% of all male cancer-related deaths (2). Prostate cancer-caused death may due to its invasion and metastasis (3). It has been shown that most patients can survive longer than 10 years if metastasis does not occur (4). Surgery, radiation therapy, endocrinological treatment and chemotherapy have been used in the treatment of prostate cancer alone or in combination. However, current therapies are not often curative and relapse often happened. Novel therapeutic strategies are needed for the treatment of prostate cancer. Some studies have shown that active components of natural products (5, 6), antagonists of hypothalamic growth hormone-releasing hormone (7), monoclonal antibodies (8, 9) and microRNA (10, 11) could suppress prostate cancer in vitro or in vivo.
Bone morphogenetic proteins (BMPs) can induce the formation of bone and cartilage. They are also involved in pivotal morphogenetic signals (12). Cancerous disease is often related with misregulation of the BMP signaling system. Recombinant human BMPs (rhBMPs) have been used in orthopedic applications. RhBMP2 causes more overgrown bone than any other BMPs and has been well accepted as an osteoinductive growth factor. BMPs and /or BMP receptor are also expressed in several malignancies (13). It has been shown that BMPs could promote tumor growth, invasion and metastasis (14-16). Dai et al. (17) have shown that BMP6 promotes osteoblastic prostate cancer bone metastases. Few studies have indicated the role of rhBMP2 on prostate cancer (18, 19).
Secreted phosphoprotein 24 kD (spp24) is a bone matrix protein that can bind to and affect the activity of BMPs (20, 21). Spp24 exists in the bone environment in several forms and can inhibit the BMP-induced bone formation. Several studies have shown that spp24 could suppress pancreatic cancer growth (22) and lung cancer (23) caused by BMP2. Previous studies have shown that BMP2 could promote proliferation and migration of PC3 cells (19, 24). However, it is not clear on the effects of spp24 on prostate cancer induced by BMP2. In this study, we investigated the influence of BMP2 on prostate cancer in vitro and in vivo. Furthermore, we also showed the ability of spp24 on the effects induced by BMP2.
Materials and Methods
Cell culture and experimental materials. The human prostate cancer cell line PC-3 was obtained from America Type Cell Collection (ATCC; Manassas, VA, USA). PC-3 cells were maintained in F-12K medium with 10% FBS and antibiotics (Fisher Scientific, Pittsburgh, PA, USA) in a humidified atmosphere of 95% air and 5% CO2 at 37°C with medium change every three days. Recombinant human BMP2 was purchased from MEDTRONIC, INC. (Medtronic Sofamor Danek, Minneapolis, MN, USA). Recombinant spp24 was produced in a bacterial expression vector as described previously (25).
Cell viability assay. PC-3 cells (4×103 cells/well) were plated in 96-well plates in F-12K medium containing 10% FBS and incubated for 24 h. Then, the cells were cultured with serum-free medium containing different concentrations of RhBMP2 and Spp24 (50 ng/ml BMP2 and 50 ng/ml BMP2+100 μg/ml Spp24) for 24, 48 and 72 h, respectively. Then, the medium was removed and another medium (without FBS) containing 10% 3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide (MTT) solution (0.5 mg/ml in PBS, Sigma, MO, USA) was added into each well. After a 4-h incubation at 37°C, the fomazan crystals formed were dissolved in 100 μl of dimethyl sulfoxide (Sigma, St. Louis, MO, USA). The absorbance at 570 nm was read using a Biotek microplate reader (BioTek Instruments Inc, Winooski, VT, USA). The data were displayed as the mean±SD of six wells for each group.
In vivo experimental groups. All mice underwent implantation of cells in both a subcutaneous model and an intratibial injection. Then, they were randomly divided into three groups. A control group received PC-3 cells alone; BMP-2 group received PC-3 cells+RhBMP2 (10 μg); spp24 group received PC-3 cells+RhBMP2 (10 μg)+spp24 (1.0 mg). There were six animals in each group. Repeat injections of 2 μg RhBMP and spp24 (1.0 mg) were given weekly until the end of the study period.
Subcutaneous tumor model and intratibial tumor model. The subcutaneous tumor xenograft model was established as previously reported (23). Eighteen 8-week-old male nude mice (BALB/c nu/nu) weighted 20-25 g were housed under specific pathogen-free facilities. All mice were acclimated for one week and, then, received subcutaneous injections of 5×105 PC-3 cells on their back. Briefly, PC-3 cells were suspended in 30 μl of PBS. After the mice were anesthetized with ketamine (4.0 mg/100 g), the skin on back was prepped with 70% ethanol and betadine. The PC-3 cells were then injected into the subcutaneous space on the back of mice. All the experiments were carried out according to the guidelines of animal welfare committee. The protocol adopted here was approved by the Animal Use Committee of Renji Hospital.
In addition, the intratibial tumor model was also established as previously reported (23). Briefly, after anesthetization, the skin was prepped with 70% ethanol and betadine. Then, a total of 1×106 cancer cells in 30 μl of PBS were injected into the proximal tibial cavity through the proximal tibial plateau with a 27.5-gauge needle. The wound was closed with VICRYL® suture.
Measurement of tumor size and weight. Tumor size and weight were measured as previously described (23). The tumor sizes were measured weekly using digital calipers and calculated by the average of the greatest length and width. All mice were sacrificed at 10 weeks. The tumor specimens were dissected and tumor weights determined using a precision electronic balance.
Radiographic analysis. Prior to their sacrifice, animals were anesthetized as described above. Radiographs were obtained using a Radiography System (Faxitron, IL, USA) exposed to X-ray at 35 kVp for 6 s. The bone lesions were evaluated blindly by two observers. The bone lesions were scored as following: 0, normal; 1, lystic lesions present within the medullary canal only; 2, involving one cortex; 3, involving two cortexes. The X-ray images were evaluated by three independent observers who were blinded with the study groups.
In vivo terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) apoptosis assay and immunohistochemistry. The tumor specimens were cut into pieces and fixed in 4% paraformaldehyde at 4°C for 72 h, dehydrated through a series of ascending ethanol solution (40%-100%), embedded in paraffin and sliced for TUNEL apoptosis assay and immunohistochemistry.
TUNEL apoptosis assay. Sections were firstly dewaxed with dimethylbenzene and rehydrated with ethanol solution (100%-70%) and distilled water. Then, the sections were incubated with 20 μg/ml proteinase K (without DNase) for 20 min at 37°C. After washing for three times, the sections were incubated with 3% H2O2 solution for 20 min at room temperature. After washing with PBS for three times, they were incubated with the TUNEL apoptosis assay solution for 60 min at 37°C. The reaction was stopped using the solution supplied by the TUNEL apoptosis assay kit. Then, the sections were incubated with horseradish peroxidase (HRP)-conjugated streptavidin solution for 30 min at room temperature. Finally, sections were developed with DAB kit and hematoxylin stanining. Four randomly selected fields from each section were analyzed. The percent of TUNEL-positive apoptotic cells were calculated in each mouse.
Immunohistochemistry. Tumor sections were firstly dewaxed and rehydrated. Then, the sections were heated with pressure in retrieval solution for 30 min. After incubated with 5% BSA to block non-specific staining, sections were incubated with proliferating cell nuclear antigen (PCNA) monoclonal antibodies diluted as instructed overnight at 4°C. Then, the sections were incubated in HRP-conjugated secondary antibody at 37°C for 1 h. Finally, sections were developed with DAB kit. The images were acquired using an Olympus Microscope (CX40, Olympus, Japan) with a digital camera (ProgRes C14; Jenoptic, Jena, Germany). PCNA-positive cells were numbered using software (Digital Micro System, Jena, Germany). Four randomly selected fields from each section were analyzed. The percent of PCNA-positive cells were calculated in each mouse.
Statistical analysis. Data management and statistical analysis were performed by SPSS16.0 (SPSS Inc, Chicago, IL, USA). Data were expressed as mean±standard deviation and analyzed by Mann–Whitney U-test. Statistical significance was defined as p≤0.05.
Results
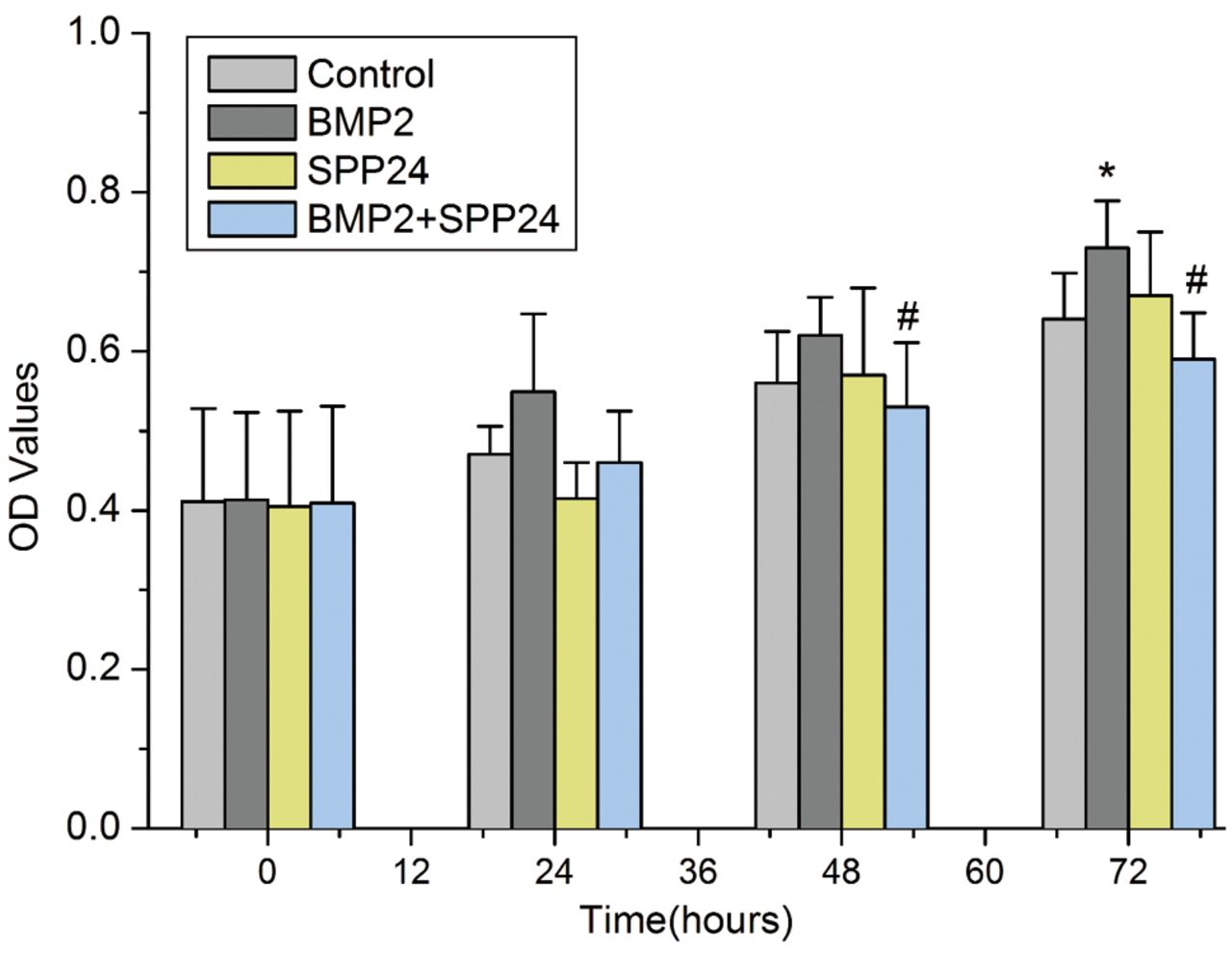
Spp24 inhibited PC-3 cell proliferation stimulated by BMP2. Figure 1 shows the PC-3 cell proliferation. The PC-3 cell proliferation was increased with time in all groups. PC-3 cells treated with BMP2 showed an increase in proliferation files compared with the control, especially at 72 h (p<0.05). However, spp24 changed PC-3 cell ability stimulated by BMP2, in particular at 48 and 72 h (p<0.05).
Spp24 inhibited the subcutaneous tumor growth promoted by BMP2. We further observed the effects of BMP2 and spp24 on tumor growth in vivo using a subcutaneous tumor model (Figure 2). The tumor sizes were increased over time (Figure 2A and 2B). BMP-2 significantly promoted tumor growth during the observation period. BMP-2 had increased the tumor size by 2.0-fold (Figure 2B) and the tumor weight by 3.0-fold (Figure 2C) compared with the control. The spp24 significantly blocked the effects of BMP2 on tumor growth. In some mice treated with spp24, the size of the tumor mass became almost undetectable (Figure 2A).
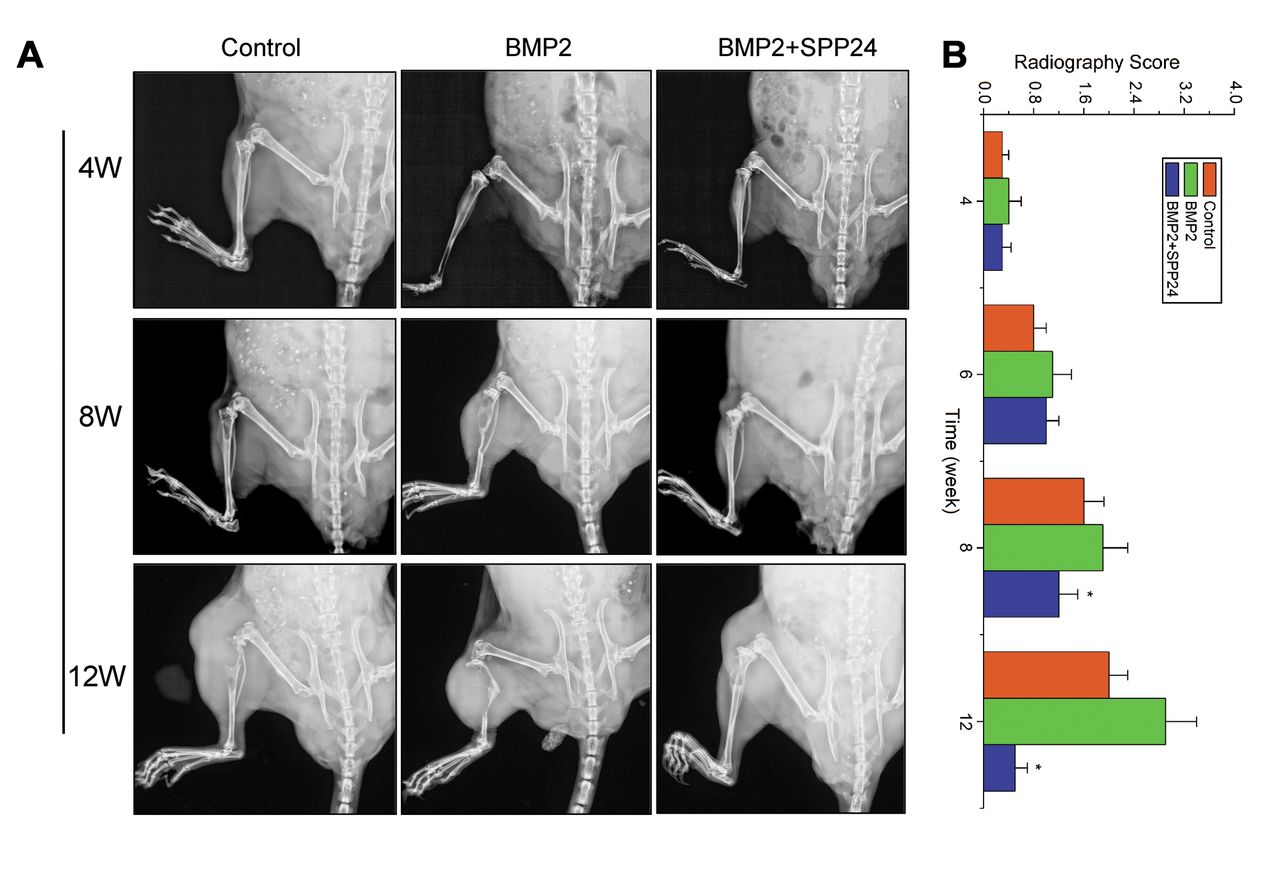
Spp24 inhibited the intratibial tumor growth promoted by BMP2. We also observed the effects of BMP2 and spp24 on tumor growth in vivo using an intratibial tumor model (Figure 3). Excellent agreement was found between observers (Kappa value=0.86). The bone destructions were heavier in mice treated with BMP2 than the control (Figure 3A). The radiological score was significantly higher in BMP2-treated mice compared with the control (Figure 3B). However, spp24 significantly inhibited the bone destructions stimulated by BMP2. The radiological score was obviously decreased in treated mice with both BMP2 and spp24 compared with the mice in control and the mice treated with BMP2 alone (Figure 3B).
TUNEL- and PCNA-positive cell in tumor specimens. Cell apoptosis in tumor specimens were assayed using TUNEL staining (Figure 4). Few TUNEL-positive cells were observed in control and BMP2 groups (<5.0%). However, obvious TUNEL-positive cells were found in tumors of mice treated with both BMP2 and spp24. Spp24 probably inhibited tumor growth by inducing tumor cell apoptosis.

The effects of spp24 on the proliferation of PC-3 cells stimulated by bone morphogenetic protein-2 (BMP2). BMP2 significantly stimulates PC-3 cell proliferation at 72 h. However, spp24 could abolish the effects of BMP2 at 48 and 72 h. *p<0.05 vs. control; #p<0.05 vs. BMP2 group.
We also investigated the tumor cell proliferation activity in vivo by immunohistochemical staining for PCNA (Figure 5). BMP2 groups showed stronger PCNA-positive cells compared with the control and BMP+spp24 group. Consistent with the in vitro data, spp24 also inhibited the in vivo cell proliferation stimulated by BMP2.
Discussion
Several therapeutic strategies, such as surgery, radiation therapy, endocrinological treatment, have been used for prostate cancer management. However, current therapies are not often curative and relapses often happen. Some studies have indicated that tumor originating from prostate can express BMPs (19, 26, 27) and BMPs may stimulate tumor cell proliferation, invasion and metastasis in vitro and in vivo (27). In this study, we showed that BMP2 could stimulate prostate cancer cell proliferation in vitro and tumor growth and bone metastasis in vivo. We further demonstrated that a bone matrix protein, spp24, could abolish the effects of BMP2 on prostate cancer. Our data showed that spp24 have great potential to be a therapeutic agent.
BMPs are a group of growth factors. BMP2 through BMP7 belong to the transforming growth factor-beta (TGF-β) superfamily of proteins. Misregulation of BMP signal system may be related with the cancerous disease. Several studies have shown that BMPs could promote tumor invasion and metastasis (15-17). However, few studies have observed the effects of BMPs, especially BMP2, on prostate cancer. It has been shown that PC-3 cells expresse the BMP receptor (19). We hypothesized that BMP may influence the activity of PC-3 cells. Few previous studies have demonstrated that BMP2 stimulates the proliferation, migration and invasion of PC3 cells (19, 24). In the present study, we observed that BMP2 could stimulate PC-3 cell proliferation in vitro. In subcutaneous and intratibial tumor model, our data also showed that BMP2 markedly promoted the subcutaneous and intratibial tumor growth. PCNA-1, a proliferation biomarker, positive cells were commonly found in tumor specimens of BMP2-treated mice, which also indicated that BMP2 dramatically stimulated cell proliferation. Our data further confirmed previous findings that BMP could stimulate cancer cell growth in vitro and in vivo.

Subcutaneous tumor formation and growth in control, bone morphogenetic protein 2 (BMP2) and BMP2+ secreted phosphoprotein 24 kD (SPP24) groups at 12th week. A: The gross observation at necropsy; B: The tumor growth curves; C: The tumor weight at the 12 weeks, p<0.01 vs. other groups.

Intratibial tumor growth and radiographic analysis. A: The bone lesions at 4, 8 and 12 weeks in control and experimental groups. B: The radiographic score at 4, 6, 8 and 12 weeks, *p<0.01 vs. bone morphogenetic protein 2 (BMP2) group. SPP24, Secreted phosphoprotein 24; W, week.
It has been shown that inhibition of BMP/TGF-β activity may limit tumor growth (23). Spp24 can bind with BMP2 and inhibit its osteogenic activity (28, 29). Several studies have indicated that spp24 could inhibit cancer cell proliferation and tumor growth (22, 23). Lee et al. showed that spp24 could ameliorate the effects of BMP2 on lung cancer in a subcutaneous and intratibial tumor model (23). Recently, Li et al. indicated that spp24 could block the stimulatory effects of BMP2 on pancreatic cancer in vitro and in a xenograft tumor model (22). However, it is not clear whether spp24 could influence the effects of BMP2 on prostate cancer. Our data showed that spp24 could ameliorate the effects of BMP2 on prostate cancer in vitro and in vivo. Our results also support the fact that spp24 may have great potential for the treatment of cancerous diseases.
Bone metastasis is the common complication of prostate cancer. Usually, bisphosphonate agents and radionuclide treatment are adopted in metastasized bone lesions. Lai et al. reported that osteoblast-derived BMP2 may enhance the migration of prostate cancer cell through Akt/ERK pathway (19, 30). Feeley et al. (19) also showed that BMPs influence the formation of the osteolytic prostate cancer lesions and noggin, a BMP antagonist, could inhibit the BMP-induced growth of osteolytic prostate cancer metastasis. Therefore, a BMP2 inhibitor may be used to block bone metastasis of prostate cancer. Lee et al. demonstrated that spp24 could inhibit the bone destruction caused by lung cancer cells, thus indicating that spp24 may be used for treatment of bone metastasis (23). In our study, we also observed that spp24 could inhibit the bone destruction induced by BMP2.
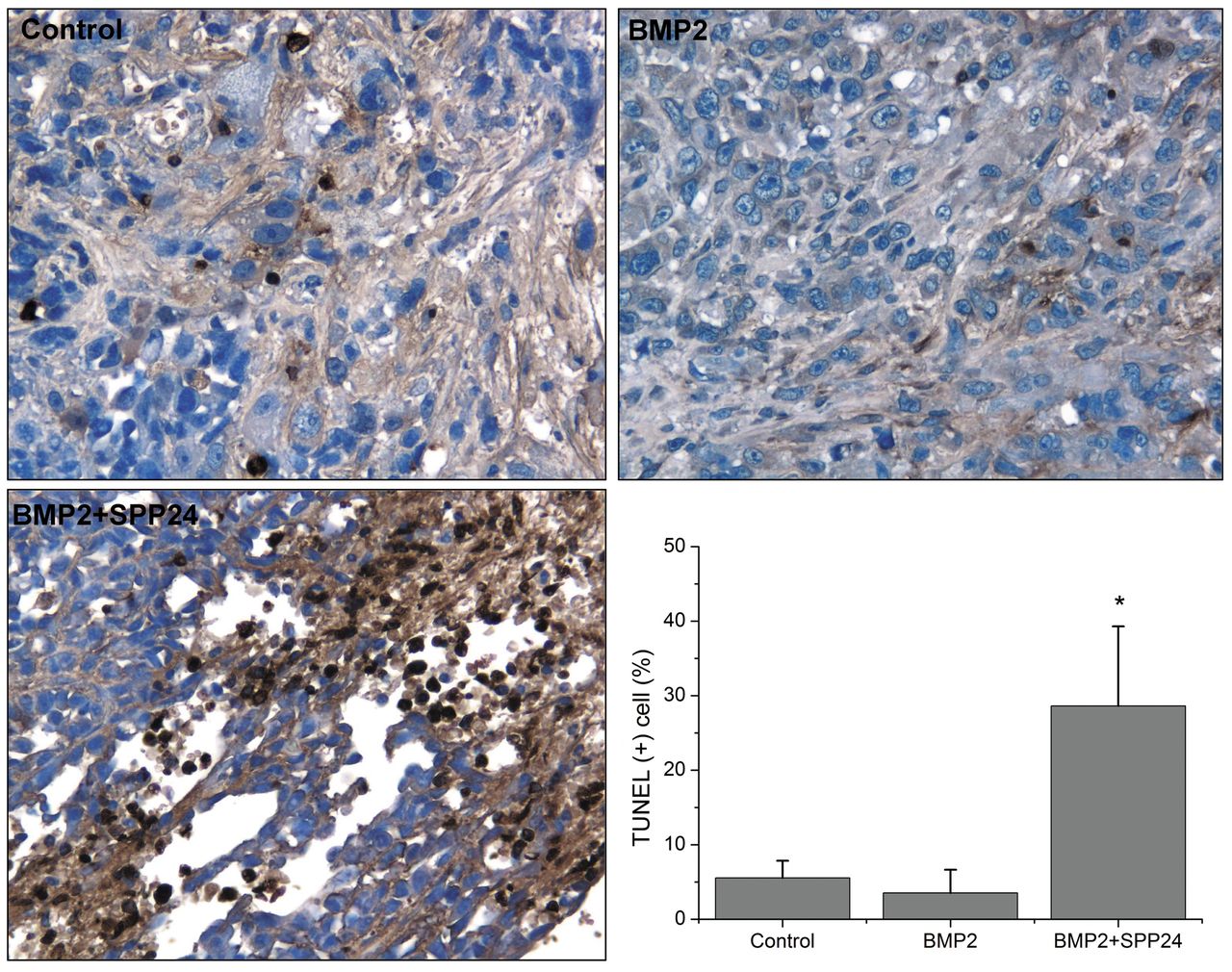
Cell apoptosis assay in vivo specimens using terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) method (×200). No differences for cell apoptosis were found between the control and BMP2 group. More TUNEL-positive cells were found bone morphogenetic protein-2 (BMP2) + secreted phosphoprotein 24 kD (SPP24) group compared to the control and BMP2 group. *p<0.01 vs. control and BMP2 group.
The mechanism of spp24 on BMP2-induced cell proliferation is not completely understood. Spp24 may bind with BMP2 and, consequently, inhibit the function of BMP2. In addition, some studies have shown that spp24 could induce cancer cell apoptosis in vitro and in vivo (22). In our study, we also found that spp24 dramatically induced apoptosis of tumor cells; however, BMP2 did not affect cell apoptosis.
There are several limitations of this study. The mechanism of spp24 on BMP-induced effects is not investigated. Further exploration is needed. In addition, the bone metastasis model is established by intratibial injection of tumor cells, which is different with the pathophysiological conditions. Moreover, we did not show the effects of spp24 alone on prostate cancer growth, in this study.
In conclusion, our data showed that BMP2 can stimulate prostate cancer cell proliferation, tumor growth and bone metastasis. Spp24 obviously ameliorates the effects of BMP2 on prostate cancer. Spp24 may have a great potential for the treatment of prostate cancer and its bone metastasis.
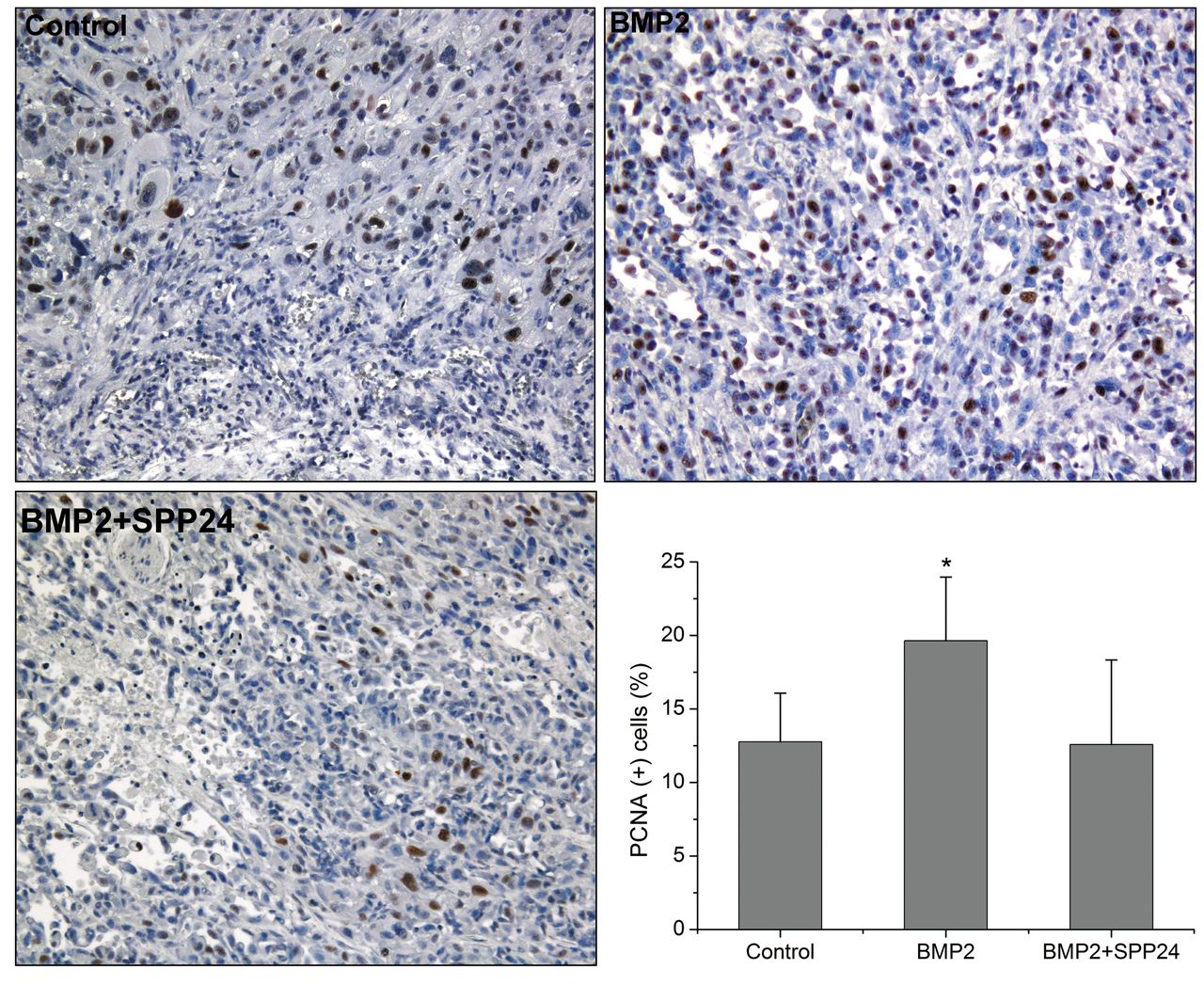
Immunohistochemical staining for proliferating cell nuclear antigen-1 (PCNA-1) in vivo specimens (×100). More PCNA-1 positive cells were found in bone morphogenetic protein2 (BMP2) group compared with control and BMP+ secreted phosphoprotein 24 kD (SPP24) group. *p<0.01 vs. control and BMP2+SPP24 group.
Acknowledgements
This research was funded by Shanghai Pujiang Program (NO. 15PJD026), Medical-Engineering Joint Fund of Shanghai Jiao Tong University (NO. YG2014MS51), Shanghai Fund for Young Physician Development (NO. 20141051), Incubating Program for Clinical Research and Innovation of Renji Hospital (NO. PYXJS16-006).
Footnotes
This article is freely accessible online.
Conflicts of Interest
The Authors declare that they have no conflict of interest.
- Received June 7, 2016.
- Revision received July 4, 2016.
- Accepted July 5, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}