


Abstract
Background: There exist limited therapeutic opportunities regarding the treatment of endometrial cancer (EC), and novel therapies based on the molecular profiling of EC cells are required. Materials and Methods: We used microarray analysis of EC tumour samples in order to identify tumour-specific changes regarding gene expression. Results: It was found that gremlin 2, an inhibitor of bone morphogenetic protein (BMP) signaling, was repressed in EC samples, and that gremlin 2 inhibited tumour cell growth. Conclusion: Down-regulation of gremlin 2 may lead to carcinogenesis and progression of EC. We suggest that re-activation of gremlin 2-associated pathways could suppress EC progression and should thus be explored as a potential novel therapeutic approach.
Endometrial cancer (EC) is the most common type of gynaecological cancer, with an estimated worldwide incidence of approximately 320,000 in 2012; the incidence of the disease is increasing in recent years (1, 2). There exist few treatment options available regarding advanced and recurrent EC, and this has prompted the quest for innovative approaches, including targeted therapies.
Aberrant proliferation, migration, and metastasis are phenotypic hallmarks of cancer, in which the TGFβ superfamily of bone morphogenetic proteins (BMP) play critical roles (3). Gremlins are small, secreted single-domain proteins that antagonize BMP signaling. Gremlin 2, formerly known as protein related to Dan and Cerberus (PRDC), was isolated in 1998 from mouse embryonic stem cells, where it plays a role in neural development (4). Gremlin 2 inhibits BMP2 and BMP4 ligand-dependent signaling (5, 6). In animal models, gremlin 2 has been implicated in the development of both the embryo overall, and the ovarian follicle (5, 7-10). Furthermore, Yeung et al. reported that gremlin 2 inhibited BMP2-driven phosphorylation of Smad1/5 by BMP2 in an osteogenic differentiation model (11). In human ovarian tissue, we found that gremlin 2 reduced phosphorylation of Smad1/5/9 and maintained the arrest of primordial follicles, probably through inhibition of BMP4-mediated follicle development (unpublished observations). To the best of our knowledge, there has been no report of gremlin 2 in association with cancer. We analyzed EC microarray data and found that gremlin 2 was specifically repressed in tumour samples. Consistent with a potential anticancer role, we found that exposure to gremlin 2 suppressed the proliferation of EC cell lines.
Materials and Methods
The present study was approved by our Institutional Review Board (Approval No. 134) and written informed consent was obtained from all participants.
Microarray gene expression analysis. Surgical specimens were obtained from five patients with EC and five with leiomyoma and were subsequently frozen, as reported previously (12). Extracted RNA was assessed using the SurePrint G3 Human Gene Expression 8×60K v2 Microarray Kit (Agilent Technologies, Tokyo, Japan). Data were analyzed using the Ingenuity iReport (Tomy Digital Biology, Tokyo, Japan). Differentially expressed genes were classified as those with a p-value <0.05 and fold change greater than 2.
Immunohistochemistry. Studies to assess the expression profiles of molecules identified by microarrays were conducted on paraffin-embedded tissue blocks of four patients with atypical endometrial hyperplasia (AEH) and of 18 patients with International Federation of Gynecology and Obstetrics (FIGO) stage I, grade 1 or 2 (low-grade) EC undergoing surgery at our Hospital. Immunohistochemistry study of the most significantly repressed genes identified by microarray analysis in EC was then conducted. Sections of paraffin blocks were treated with antigen retrieval solution according to the manufacturer's instructions (Target Retrieval Solution S1699; Dako Japan Inc., Tokyo, Japan), incubated with an antibody against gremlin 2 (dilution 1:125) (Sigma-Aldrich, Tokyo, Japan) and stained using ImmPRES reagent (Vector Laboratories, Burlingame, CA, USA), secondary antibodies, and 3,3’-diaminobenzidine peroxidase substrate kit (Vector Laboratories, Burlingame, CA, USA).

Statistical significance vs. fold change of 1,346 genes via microarray analysis. Analysis of microarray data was performed using Ingenuity iReport. Probe set intensities were summarised and normalised using RNA. Genes that were significantly differentially expressed were identified following application of a moderated t-test with a false discovery rate-adjusted p-value cut-off of 0.05 and fold change cut-off of 2, yielding 1,346 differentially expressed genes.
Cell culture. The human endometrial cancer cell line HEC-1A was obtained from ATCC (Manassas, VA, USA). The Ishikawa human endometrial cancer cell line was kindly provided by Dr. Masato Nishida (Tsukuba University, Ibaraki, Japan) (13, 14). Ishikawa cells were cultured in RPMI-1640 medium and HEC-1A cells were cultured in McCoy's 5A Modified Medium (GIBCO, Rockville, MD, USA) supplemented with 10% heat-inactivated foetal bovine serum (FBS; Omega, Tarzana, CA, USA), 1% penicillin-streptomycin-amphotericin B suspension ×100 (Wako Pure Chemical Industries, Ltd, Osaka, Japan).
MTT viability assay. HEC-1A and Ishikawa cells (5×103 cells/well) were plated into 96-well culture plates and left overnight to adhere. The attached cells were treated with up to 5 μg/ml of gremlin 2 for 24 h, 36 h, or 74 h. Cell viability was measured using a modified methylthiazol tetrazolium (MTT) assay according to the manufacturer's protocol (MTT Cell Proliferation Assay Kit, Cayman Chemical Company, Ann Arbor, MI, USA). Briefly, cells were incubated with the MTT Reagent for an additional 4 h, and dissolved formazan products were measured by reading the absorbance of samples at 570 nm on a microplate reader. Each experiment was repeated at least three times.
Cell migration assay. A Radius 96-well cell migration assay (Cell Biolabs, San Diego, CA, USA) was used to measure cell migration according to the manufacturer's instructions. Briefly, HEC-1A and Ishikawa cells were seeded and incubated until the wells were 80~90% confluent. The RadiusGel was removed and cells were allowed to migrate to a cell-free area for 24 h in the presence of up to 5 μg/ml of gremlin 2. The circular space was quantitatively evaluated using the Adobe Photoshop software (Adobe Systems Incorporated, San Jose, CA, USA). The assays were performed with three replicates and repeated twice.
Statistical analysis. Statistical significance was determined using a Mann–Whitney U-test comparison between two groups. Asterisks shown in the figures indicate p-values <0.05, which were considered significant. The analyses were performed using XLSTAT 2014 (Addinsoft, Paris, France).
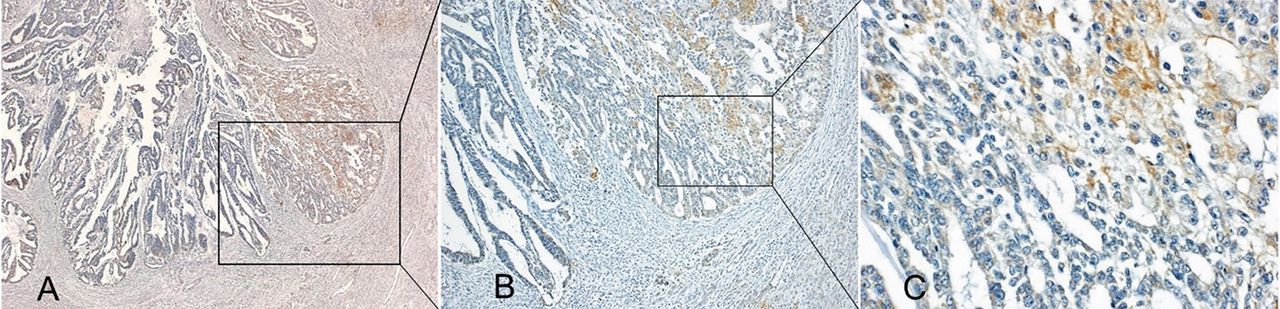
Immunohistochemical analysis of gremlin 2 at the invasion front of endometrial cancer cells of a representative case. Brown staining indicates gremlin 2 immunoreactivity. FIGO stage IA and Grade 1 endometrioid tissue. Gremlin 2 was not expressed in part of the invasion front. (A, scale 40×; B, scale 100×). Gremlin 2 was stained in the cytoplasm of endometrial cancer cells (C, scale 400×).

Gremlin 2 inhibits the growth of endometrial cancer cells in a dose-dependent manner. (A) Ishikawa and HEC-1A cells were incubated with the indicated concentrations of Gremlin 2 before being evaluated for viability using the MTT assay. (B) Inhibition of growth was time-dependent. Solid lines indicate the median absorbance of each sample following culture with Gremlin 2. Asterisks indicate a p-value <0.05, considered significant. Abs, Absorbance of samples at 570 nm.

Gremlin 2 did not inhibit or enhance migration of Ishikawa and HEC-1A cells. (A) Migration assay of HEC-1A cells after a 24-h incubation with gremlin 2. Scale bar=500 μm. (B) The graph indicates the % closure of the circular space in Ishikawa and HEC-1A cells after a 24-h incubation with up to 5 μg/ml of gremlin 2.
Results
Microarray analysis identified 1,346 genes significantly differentially expressed in EC compared to control samples. The most highly expressed gene was chemokine ligand 18 (CCL18) that was 35.6-fold higher in EC samples. The most significantly suppressed gene in EC compared to the control was gremlin 2 (8.5-fold reduction) (Figure 1).
Immunohistochemistry revealed that gremlin 2 was expressed in normal endometrium obtained from 4 patients with leiomyoma as well as in adjacent normal endometrium to AEH or EC, in 4 of 4 tissues of AEH and 5 of 5 tissues of low grade EC with FIGO stage IA without myometrial invasion. Among 13 patients with low-grade FIGO stage I with myometrial invasion, gremlin 2 was partially expressed in the myometrial invasion front in 11 (85%) patients (Figure 2).
The MTT assay revealed that gremlin 2 significantly inhibited the growth of Ishikawa and HEC-1A cells in a dose-dependent manner (Figure 3). When Ishikawa cells were incubated with 1.0 μg/ml and 5.0 μg/ml of gremlin 2 and HEC-1A cells were incubated with 5.0 μg/ml of gremlin 2, proliferation was significantly reduced compared with control without gremlin 2 (p=0.029, p=0.025 and p=0.048, respectively). This inhibition was more pronounced upon extended treatment, as proliferation of Ishikawa cells was significantly attenuated after incubation with gremlin 2 for 72 h (p=0.024). Although a similar tendency was observed in HEC-1A-treated cells, it did not reach statistical significance. On the other hand, the migration assay did not show any influence of gremlin 2 on Ishikawa or HEC-1A cells (Figure 4).
Discussion
Our microarray analysis revealed that one gene (CCL18) was up-regulated, and one gene (gremlin 2) was extensively down-regulated in EC samples compared to normal controls. We had previously reported frequent and high expression of CCL18 in EC compared to normal endometrium (12). CCL18 is a key molecule involved in the epithelial-to-mesenchymal transition, that is an essential process required for metastatic invasion. To date, there have been no reports on the association between gremlin 2 and tumorigenic state. Consistent with the microarray analysis, immunohistochemistry revealed that gremlin 2 was expressed in normal endometrium, AEH, and non-invasive low-grade EC, but was reduced in the invasive-front sections of the myometrium of low-grade EC tissue.
Gremlin 2 inhibits signal transduction initiated by BMP2 and BMP4. In primary breast cancer tissues and breast cancer cell lines, BMP4 is the most frequently and highly expressed member of the BMP family (15). In experiments using breast cancer cell lines, Ketolainen et al. reported that BMP4 stimulated cell migration and invasion, but inhibited proliferation (16). Consistent with this, Guo et al. reported that cancer cell migration and invasion were inhibited by both RNAi-mediated knockdown of BMP4, and treatment with the BMP4 antagonist, Noggin (17). Kallioniemi and colleagues reported that BMP4 expression was elevated in 25% of primary pancreatic cancer tissues, and BMP4 increased cell migration but suppressed the growth of pancreatic cancer cells (18). In contrast, BMP-2 has 92% homology with BMP-4 (19); however, there are conflicting reports regarding its role in tumour cell proliferation, invasion, and migration (20-22).
In the present study, gremlin 2 inhibited the proliferation of the Ishikawa and HEC-1A EC cell lines, but did not influence cell migration. We conducted an MTT assay to determine whether gremlin 2 affected breast cancer cell viability, and observed a significant reduction of proliferation (unpublished data). We assume that the role of gremlin 2 in cancer is to antagonize BMP-4, in much the same manner as it acts during ovarian follicle development (6). However, our findings would then not be consistent with previous studies of BMP-4 in cancer cell lines and tissues. Gremlin 2 probably has effects on other signal transduction pathways, including those in which BMP-2 is involved.
We suggest that this activation of gremlin 2, or the pathways associated with it, should be investigated for potential strategies to treat EC.
Acknowledgements
A part of the microarray analysis of CCL18 was previously reported in Anticancer Research (13). The Ishikawa human EC cell line was kindly provided by Dr. Masato Nishida (Tsukuba University, Ibaraki, Japan). This study was supported by Grant-in-Aid for Researchers, Hyogo College of Medicine (Hiroshi Tsubamoto) and by Grant-in-Aid for Graduate Students, Hyogo College of Medicine (Riya Sakane).
- Received September 17, 2015.
- Revision received October 22, 2015.
- Accepted October 30, 2015.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.