


Abstract
Background/Aim: Lipopolysaccharide (LPS), a major component of the cell wall of Gram-negative bacteria, is known to possess strong immune-regulatory activity. We have found and reported the existence of biologically-active LPS in acetic acid bacteria. The LPS shows Limulus-positive activity and activation of macrophages to produce nitric oxide and tumor necrosis factor. In this study, we investigated the anti-allergic effect of an orally-administrated acetic acid bacteria extract containing LPS; the cedar pollinosis model was used. Materials and Methods: Acetic acid bacteria were isolated from various fruits by Nodai kaihen medium. Then, the anti-allergic effect of acetic acid bacteria extracts was investigated. BALB/c mice were immunized with a mixture of cedar pollen and alum into their peritoneal cavity; they also received additional immunizations of pollen to nasal cavity. After immunizing the mice with pollen into their nasal cavity to trigger an allergic reaction, the frequency of nose scratching was counted for 5 min. Results: The bacteria were cultured and prepared and the water-extract contained about 1-10 mg/ml of Limulus positive substances. The extract of acetic acid bacteria induced higher levels of interleukin (IL)-10 and FOXP3 mRNA expression in macrophages (RAW246.7 cell), as assessed by DNA microarray analysis. Oral administration of the acetic acid bacteria extract demonstrated significantly less scratching numbers than control water group with pollen immunization. Conclusion: These results showed that LPS in acetic acid bacteria has the potential to protect from an allergic reaction, especially from cedar pollinosis.
In recent years, immunopotentiating effects in many fermented foods have been reported, thus highlighting health-maintaining functions of foods (1). Fermented foods make use of certain microorganisms, such as the lactic acid bacteria Bacillus subtilis and yeasts; however, the use of Gram-negative bacteria is comparatively lesser known in this field. Gram-negative bacteria utilized for food production include Acetobacter aceti for vinegar, Xanthomonas campestris for xanthan gum and Acetobacter xylinum for nata de coco. These foods are believed to possess immunopotentiating effects as they contain ingredients derived from acetic acid bacteria. However, their active ingredients remain mostly unknown. We have previously discovered that acetic acid bacteria, the most extensively used class of Gram-negative bacteria in the world, (i) contain Limulus-positive substances; (ii) stimulating cultured macrophages leads to induction of tumor necrosis factor and nitrogen oxide; and (iii) these reactions are inhibited by polymyxin B (1). Moreover, we have succeeded in purifying lipopolysaccharides (LPS) from acetic acid bacteria, thereby demonstrating LPS presence in these bacteria. It has been previously reported that LPS from Gram-negative plant symbiotic bacteria in edible plants has a long history of being consumed in foods and that oral administration of LPS is industrially useful (2-7). Thus, acetic acid bacterial cells containing LPS are likely to be involved in the immune activation by fermented foods containing these bacteria.
The term “acetic acid bacteria” is the collective name for Gram-negative bacteria that produce acetic acid by oxidizing ethanol; these bacteria are found in genera such as Acetobacter, Gluconobacter and Gluconoacetobacter. Foods that contain acetic acid bacteria cells include nata de coco (8), tea fungus (Kombucha) (9) and Caucasian yogurt (10). These foods have been shown to possess physiological activities, such as anti-inflammatory and anti-allergic effects (11, 12). However, these effects have been demonstrated using the finished food products. The amount of acetic acid bacteria contained in these foods is minute and it is unclear whether the bacteria themselves are responsible for these beneficial effects.
In the present study, our objective was to clarify whether acetic acid bacteria consumed as a part of food exhibit anti-allergic activity. Mice were allowed to freely consume LPS-containing water extracts of a standard strain of acetic acid bacteria (A. aceti) and a strain isolated from the fruit Gluconobacter suboxydans; we examined their effects in a pollinosis model and measured their cytokine expression in the spleen. We found that the LPS-containing extracts of acetic acid bacteria exhibited anti-allergic activity via oral administration and demonstrated the enhanced expression of anti-inflammatory cytokines.
Materials and Methods
Preparation of acetic acid bacteria extract. G. suboxydans was obtained as follows: First, the fruit was immersed in an isolation culture medium for acetic acid bacteria (Nodai kaihen medium, 1% glucose, 1.5% polypeptone, 0.8% yeast extract, 0.5% ethanol, 0.3% acetic acid, 0.02% cycloheximide; supplier, address) and left for three to seven days at room temperature. Then, the surface layer of the medium was spread on a plate comprising the isolation medium and agar and the plate was incubated at 30°C. After colonies emerged, they were dissolved in distilled water and heated at 60°C for 15 min. The sample was centrifuged and the supernatant was collected. Using this supernatant as a template source, polymerase chain reaction (PCR) was performed with 16s RNA primers to determine DNA sequence. The sequence was subjected to a homology search on the DNA Data Bank of Japan (DDBJ) and a phylogenic tree was formed using the TreeView software (link or supplier with address). The sample was then cultured in a 3 litter jar fermenter and bacterial cells were collected by centrifugation. The collected bacteria cells were then subjected to phenol–water extraction to extract acetic acid bacteria-derived LPS in accordance with the method established by Westphal et al. (13). The resultant extract was dialyzed to remove phenol. Subsequently, deoxyribonuclease (DNase) was added to obtain the final concentration of 20 μg/ml. After reacting at 37°C for 2 h, ribonuclease (RNase) was added to obtain the concentration of 40 μg/ml and allowed to react at 60°C for 30 min. Finally, Proteinase K was added and the mixture was incubated at 37°C overnight, after which high-purity LPS was obtained from the acetic acid bacteria through phenol extraction. For measuring Limulus, a chromogenic method was employed using a commercially available instant kit, Endospecy ES-50M (Seikagaku Corporation, Tokyo, Japan). LPS purified from the acetic acid bacteria contained neither nucleic acids nor proteins. The molecular weight distribution was determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE); we confirmed that the LPS obtained was equivalent to that described in previous reports (data not shown).
Allergy test. The glycolipids from the acetic acid bacteria were dissolved in pure water at 1 μg/kg/day. We allowed the mice to drink the solution ad libitum from a water jar throughout the experimental period. For mice in the control cage, distilled water was given instead. To BALB/c female mice, a suspension containing a mixture of a cedar pollen extract (Biostir, Kobe, Japan) at 30 μg/300 μl and alum (Imject Alum; Thermo Scientific, Waltham, MA, USA) was intraperitoneally administered. This administration was repeated 3 and 8 days later. The control group received a PBS solution instead (primary sensitization). Afterwards, an antigen solution of the cedar pollen extract was injected into the nostrils of the mice at 0.5 μg/5 μl. This intranasal sensitization was performed within two weeks. One week after the last intranasal sensitization, the cedar pollen extract antigen solution was intranasally injected again and the frequency of nose scratching by the mice was measured for 10 min thereafter.
cDNA microarray analysis. Subsequently, we collected blood and spleen samples from the mice. After preparing serum from the blood samples, total immunoglobulin E (IgE) levels were measured by ELISA (Morinaga Inst. Biol. Sci., Yokohama, Japan). RNA was extracted from the spleen samples with TRIzol (Life Technologies Corporation, Carlsbad, CA, USA); the RNA extract from each animal was mixed at an equal volume in each of the respective test groups. Using these pooled RNA samples, we performed semiquantitative gene expression analysis with the fibrous DNA microarray Genopal® (Mitsubishi Rayon, Tokyo, Japan).
Statistical analysis. For statistical comparisons, two-way ANOVA with Bonferroni post hoc tests were used. All statistical analyses were performed with the Prism 5 software (Graphpad, Software, La Jolla, CA, USA).
Results
Anti-pollen effect by LPS administration. Hot water extracts of a standard acetic acid bacterial strain and an acetic acid bacterial isolate obtained from the fruit (apple, persimmon, pear, plum, grape) by screening (G. suboxydans) were given to pollen-immunized mice every day via their drinking water at the same time as the initial immunization. With pollen administered to their nostrils, the frequency of nose scratching by the mice was measured for 10 min thereafter (n=3-10). The results, shown in Figure 1, revealed that the immunized mice scratched their noses noticeably more frequently, whereas those that received the standard strain of acetic bacteria (Figure 1A) or G. suboxydans from persimmon (Figure 1B) at an LPS equivalent of 1 μg/kg/day exhibited significantly less nose-scratching activity, which was similar in frequency to that in the non-immunized control mice.
Serum IgE concentration. Using ELISA, we measured the blood IgE levels of the pollen-immunized mice and the acetic acid bacteria-derived LPS-fed mice. As shown in Figure 2, IgE level was 2.6 ng/ml in the non-immunized mice and 61.5 ng/ml in the pollen-immunized mice, thus showing an apparent induction of blood IgE production in the immunized mice. Conversely, the mice that were orally administered LPS had an IgE level of 47.2 ng/ml, thus exhibiting a 23% decrease. However, this is not indicative of an apparent suppressive effect on IgE production.

Pollinosis suppression effect of acetic acid bacteria-derived LPS extracts. BALB/c mice, with pollinosis induced by immunization and cedar pollen, were fed acetic acid bacteria extracts (at a bacterial cell equivalent of 100 μg/kg, an LPS equivalent of 1 μg/kg/day) in their drinking water, and the frequency of nose scratching induced by intranasally-administered pollen was measured. A: Mice given an extract of Acetobacter aceti (n=3). B: Mice fed with an extract of Gluconobacter suboxydans (n=5-10).
Cytokine expression in LPS-ingested mouse. Cytokine gene expression in the spleen was analyzed using a highly quantitative DNA array. For each sample, the spleens from four animals in each group were collectively subjected to RNA extraction and analysis. From a total of 209 genes analyzed, we selected genes that were expressed at a sufficient level and, thus, of high reliability with a two-fold or greater difference in the expression level between the LPS-administered and non-administered groups. The results are shown in Figure 3. In the group that received an aqueous extract of acetic acid bacteria, mRNA expression levels of Ccl2 (chemokine (C-C motif) ligand 2), Ccl7 (chemokine (C-C motif) ligand 7) and Mmp12 (Matrix metalloproteinase-12) genes were increased, whereas expression levels of Ccr3 (C-C chemokine receptor type 3), FcerI (high-affinity IgE receptor), Il9 (interleukin-9) and Isg15 (interferon-induced 17 kDa protein, CD49b, integrin alpha subunit) genes were decreased. Products of these genes have been reported to be involved in anti-allergic effects and tissue repair mechanisms.
Discussion
Because acetic acid bacteria can survive in high-sugar or alcohol environments, they are often found in very sweet fruit or flower nectar. In the present study, we were able to isolate acetic acid bacteria from readily-available fruit. The isolated bacteria were confirmed to produce acetic acid from alcohol (data not shown). Acetic acid bacterial products have been shown to exhibit, as physiological activities, anti-inflammatory and anti-allergic effects (Figure 1). Although these bacteria contain polysaccharides and nucleic acids, the most influential component for the anti-allergic effect is thought to be LPS. For this reason, we focused our attention on the LPS content of the acetic acid bacteria to examine their anti-allergic efficacy.
The anti-allergic effect has been reported for foods containing acetic acid bacteria cells and is also suggested on the basis of the gene expression analysis of acetic acid bacteria-derived LPS (11, 12). In the present study, we examined whether oral ingestion of the cell components of acetic acid bacteria could lead to suppression of pollinosis. As shown in Figure 1, an apparent suppression of pollinosis was observed in the mice that received the bacterial components simultaneously with immunization. In the mice that were ingested with acetic acid bacteria-derived LPS and presented less frequent nose scratching, the blood IgE level was reduced by 23% (Figure 2). We believe that this level of decrease in the induction of IgE production is not sufficient to completely explain the strong pollinosis suppression. Therefore, we hypothesize that there must be another mechanism responsible in addition to IgE production.
In a report on cytokine production analysis in mice that were given an LPS-rich food, activation of regulatory T-cells (Tregs) and suppression of B-cells have been suggested (14). To investigate the anti-allergic effects of an acetic acid bacteria-derived LPS extract, we subjected the spleens of pollen-immunized mice to gene expression analysis (Figure 3). The results of the analysis showed increased expression levels of Ccl2, Ccl7 and Mmp12 and decreased expression levels of Ccr3, high-affinity IgE receptor, also known as FcerI, Il9, Isg15 and Cd49b. Both of Ccl2 and Ccl7 have almost identical temporal and spatial profile. CCL2 significantly stimulated healing in diabetic wounds by restoring the macrophage response (15). MMP-12 is thought to be important in the knee medial collateral ligament injury healing showing significant differences in both the early inflammatory and in the matrix tissue synthesis phases (16). CCR3 is a chemokine receptor, initially thought to be specific to eosinophils, but subsequently identified on TH2 cell subsets, basophils, mast cells, neural tissue and some epithelia. The murine CCR3 receptor regulates both the role of eosinophils and mast cells in allergen-induced airway inflammation and hyperresponsiveness (17). FcεRI is central to the induction and maintenance of an allergic response (18). IL-9 is thought to goblet cell hyperplasia and eosinophilic inflammation induced by chronic allergen exposure in a murine model. The redundancy or complementariness of IL-9, a Th2 cytokine, can occur in specific aspects of the allergic response (19). ISG15 is a crucial molecule in the regulation of the bacteria and virus protection. Human neutrophils, monocytes and lymphocytes can release ISG15; however, its function on allergic response(s) is not known. CD49b has been reported to mediate neutrophil accumulation by means of significant inhibition of KC (a CXC chemokine) response by a monoclonal antibody against CD49b (20). These findings suggest that the anti-allergic effect of acetic acid bacteria involves induction of anti-inflammatory action, as well as suppression of the expression of IgE receptors. To further elucidate these details, we plan on continuing our analysis in the future.
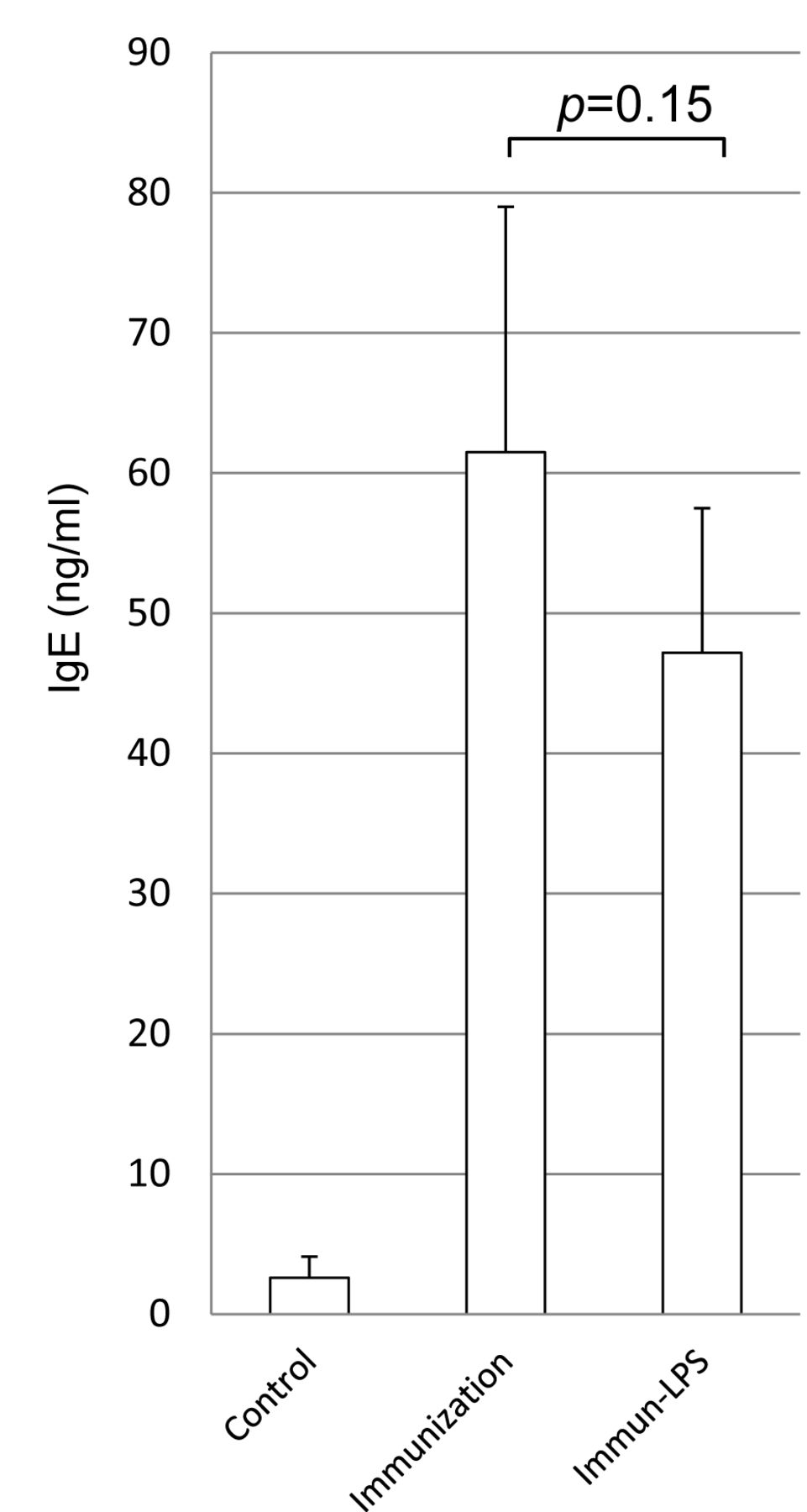
Blood IgE levels of pollen-immunized BALB/c mice with pollinosis induced by immunization with cedar pollen and fed with Gluconobacter suboxydans extract (at a bacterial cell equivalent of 100 μg/kg, an LPS equivalent of 1 μg/kg/day) in their drinking water. After intranasal administration of pollen, the frequency of nose scratching was recorded and their IgE levels were measured using ELISA.

Gene expression levels in the spleen of the murine pollinosis model. With the mice not immunized with pollen (control) as the standard, the relative levels of gene expression intensity are shown. The white column represents the pollen-immunized group that did not receive LPS, while the black column represents the pollen-immunized LPS-administered group. Spleens of identical weight were collected from five mice and collectively subjected to RNA extraction.
Footnotes
Conflicts of Interest
The Authors have no financial conflicts of interest.
- Received April 3, 2015.
- Revision received May 8, 2015.
- Accepted May 11, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Pollen Allergy Suppression Effect by the Oral Administration of Acetic Acid Bacteria (Gluconacetobacter hansenii)
- Preventive Effect of Oral Administration of Pantoea agglomerans-derived LPS in a Hypertensive Rat Model Upon Salt Loading
- Dewaxed Brown Rice Feed Improves Fatty Liver in Obese and Diabetic Model Mice
- Lipopolysaccharides Derived from Pantoea agglomerans Can Promote the Phagocytic Activity of Amyloid {beta} in Mouse Microglial Cells
- Immunopotentiator from Pantoea agglomerans 1 (IP-PA1) Promotes Murine Hair Growth and Human Dermal Papilla Cell Gene Expression
- Dewaxed Brown Rice Contains a Significant Amount of Lipopolysaccharide Pointing to Macrophage Activation via TLRs
- Effect of Lipopolysaccharide Derived from Pantoea agglomerans on the Phagocytic Activity of Amyloid {beta} by Primary Murine Microglial Cells