


Abstract
Background: Oral administration of lipopolysaccharide (LPS), a major outer-membrane component of Gram-negative bacteria, has been found to prevent infection in mammals and fish. Oral administration of LPS is believed to increase phagocytic activity and promote cytokine production, thus associating it with priming. The present study aimed to elucidate the effect of oral administration of LPS in birds, which phylogenetically lie between fish and mammals, using chickens as a model. Materials and Methods: LPS derived from Pantoea agglomerans (LPSp) was added to the feed or water for oral administration to broiler chickens. For the survival assay and gene expression analysis (Genopal), LPSp was administered at a dose of 10 μg/kg of body weight (BW)/day. LPSp was administered at a dose of 0.2 μg/kg or 200 μg/kg to stimulate peripheral blood mononuclear phagocytosis (latex beads) and nitric oxide (NO) production. Results: LPSp (10 μg/kg BW/day) administration significantly inhibited mortality in broiler chickens on commercial farms. Furthermore, oral administration resulted in a transient increase in phagocytic activity and improved the ability to produce NO. On examining splenic cytokine induction following intraperitoneal administration of LPS derived from Escherichia coli (LPSe), we found significantly increased interleukin (IL)-1β mRNA expression. Conclusion: Innate immunity activation in chickens, as seen in infection prevention, was induced by oral administration of LPSp. This infection prevention involved increased phagocytic activity and enhanced gene expression and appears to be a phylogenetically-preserved innate immunity mechanism.
Innate immunity is not only a phylogenetically-preserved biological defense mechanism at the forefront of foreign body removal but is also responsible for the crucial role of antigen presentation involved in the activation of specific immunity (1). Phagocytes (primarily macrophages), which are cells that identify and remove foreign bodies, are central to innate immunity (2). When functioning properly, innate immunity is useful for the control of infection and prevention of cancer. However, to maintain innate immunity, macrophages must remain active. Macrophages are activated in 2 stages: (i) priming and (ii) triggering of the secretion of cytokines and reactive oxygen species (3). Macrophages in the priming state exhibit no morphological change or secretion of cytokines and reactive oxygen species; however, increased phagocytic activity and secondary stimulation enhances their secretion of cytokines, including tumor necrosis factor (TNF) and interleukin (IL)-1β (4).
It has been suggested that great social benefits would be gained if the priming state could be induced in macrophages, as this would aid in preventing the spread of infection and cancer metastasis. In a study by Nakamoto et al. in mice, the stress of open surgery lasting 30 min led to severely inhibited serum TNF activity (normal period, <1%), increased mortality due to Staphylococcus aureus infection (0%→67%) and increased spread of pulmonary cancer from Meth A fibrosarcoma (37.4→54.3 lesions). However, intravenous injection of a macrophage activation substance (OK-432) 3 h prior to surgery alleviated the inhibition of TNF activity and reduced mortality, as well as the number of cancer metastases (11%, 0) (5). This suggests that activation of macrophages (increasing inflammatory cytokines) may alleviate the inhibition of innate immunity caused by severe stress and prevent infection, as well as cancer metastasis. In particular, if priming could be induced by substances that can be ingested as food, this would greatly aid in practical application. However, there are few known orally administered substances that induce a priming effect.
We previously reported that lipopolysaccharide derived from Pantoea agglomerans (LPSp), a Gram-negative plant symbiotic bacteria found in edible plants, has a long history of being consumed in foods and that the oral administration of LPS is industrially useful (6-9). LPS is a major component of the outer membrane of most Gram-negative bacteria and acts as an extremely strong stimulator of innate immunity in various eukaryotic species ranging from insects to humans (10). It has been reported that oral and percutaneous administration of LPSp effectively prevented the onset of hyperlipidemia (rabbits), diabetes (mice), infection (mice and fish) and atopic dermatitis (dogs) (7). Furthermore, it has been reported that oral administration of LPSp in chickens improved eggshell strength (11), inhibited atrophy of the spleen and bursa of Fabricius upon dexamethasone-induced stress and improved antibody titers after vaccination (12).
It has been reported that oral administration of LPSp increases infection prevention in invertebrates and crustaceans, including kuruma prawn (Penaeus japonicus) and phagocytic activity in blood cells (13). In fish, LPSp has been shown to improve infection prevention and phagocytic activity, inhibit IL-10 mRNA production and enhance IL-1β mRNA production (14). On the other hand, in mice, LPS increases TNF induction action (in mice, after oral administration of LPSp, the inducibility of serum cytokines (TNF) was significantly higher than that in the control group upon secondary bacterial stimulation) (14). Oral administration is expected to induce this priming effect in birds, which phylogenetically lie between fish and mammals; however, there is no evidence supporting such a claim. In the present study, we orally administered LPS to broiler chickens (Gallus gallus) as a bird model and examined how LPS affects priming and infection prevention. Upon oral administration of LPSp to chickens (Ross 308 broiler), the results of breeding and induction of priming were evaluated on the basis of peripheral blood mononuclear cell phagocytosis activity, nitric oxide (NO)-producing ability and splenic cytokine mRNA expression. Our results strongly suggest that oral administration of LPS induces the priming effect involved in phylogenetically-preserved innate immunity and is, therefore, useful in the industry.
Materials and Methods
Observation of mortality in broiler chicken farms. A total of 10,000 Ross 308 broiler chickens on commercial farms were randomly divided into two groups, control and one group that was administered feed containing LPSp (10 μg/kg of body weight (BW)/day) from 3 to 7 weeks of age. Two tests were conducted to examine the number of deaths from 0 to 10 weeks of age.
Cell adjustment for examination and evaluation of phagocytic function. A total of 150 female Ross 308 broiler chickens were divided into three groups. Two groups were orally administered sonde drinking water containing 200 μg/kg and 0.2 μg/kg of LPSp from 3 to 4 weeks of age. The remaining group was the control group. For breeding control, a free-range chicken coop of a mean of 4.32 m2 was used. Each experimental plot included 5 chickens. Peripheral blood was collected using a syringe with heparin every week after 3 weeks. Following blood collection, lymphocytes and monocytes were collected using Ficoll-Paque and the cell density was adjusted to 1×106 cells/ml in RPMI 640 medium containing 10% FBS. Thereafter, 10 ml of the cell suspension was added to a 60-mm collagen-coated dish and 200 μl was added to an 8-well chamber slide (Matsunami, Osaka, Japan) and left to incubate for 1 h, after which the suspended cells were removed and used in the tests.
Evaluation of phagocytic activity. Adherent cells were collected from the 60-mm dish using a scraper, adjusted to 5×105 cells/ml, and adherent monocytes were transferred to 8-well chamber slides to evaluate phagocytic activity. Fluorescein isothiocyanate (FITC)-labeled 2-μm latex beads were added to the preparations, which were then washed after 1 h to remove latex beads that were not phagocytosed. Subsequently, after staining with Wright-Giemsa stain (Diff-Quik, Sysmex, Hyogo, Japan), phagocytes were evaluated under a microscope.
Evaluation of NO-producing ability. Adherent cells were collected from the 60-mm dish using a scraper, adjusted to 5×105 cells/ml, and transferred to a 96-well plate at 100 μl per well. After 24 h, 1 ml of killed E. Coli (at a density of 107 cells/ml) was added to each well and the culture supernatants were collected after 24 h. Nitrite ion concentrations in the supernatants were measured using the Griess reagent assay (Promega, Tokyo, Japan).
Quantitative real-time reverse transcription (RT)-PCR analysis. A total of 74,000 female Ross 308 broiler chickens on commercial farms were divided into two groups. One group was administered feed containing LPSp (10 μg/kg BW/day) from 10 to 34 days of age. At 34 days of age, the broiler chickens were intraperitoneally administered LPS derived from E. coli (LPSe) at a dose of 1 mg/kg BW. Two hours later, 5 splenic specimens were collected from the 2 groups. Similarly, 5 splenic specimens were collected from the 2 groups not administered LPSe. The specimens were immediately preserved in RNAlater solution (Ambion, Austin, TX, USA). RNA was extracted from the spleens using TRIzol; then, the adjusted level of mRNA expression and cytokine mRNA expression calculated according to real-time PCR were compared. Expression of genes used in control analyses (IL-1β, IL-6, IL-8, IL-10 and IL-12) is shown as the rate per copy number of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), a housekeeping gene. Table I shows the primer information of the analyzed and housekeeping genes acquired from the literature (15-18).
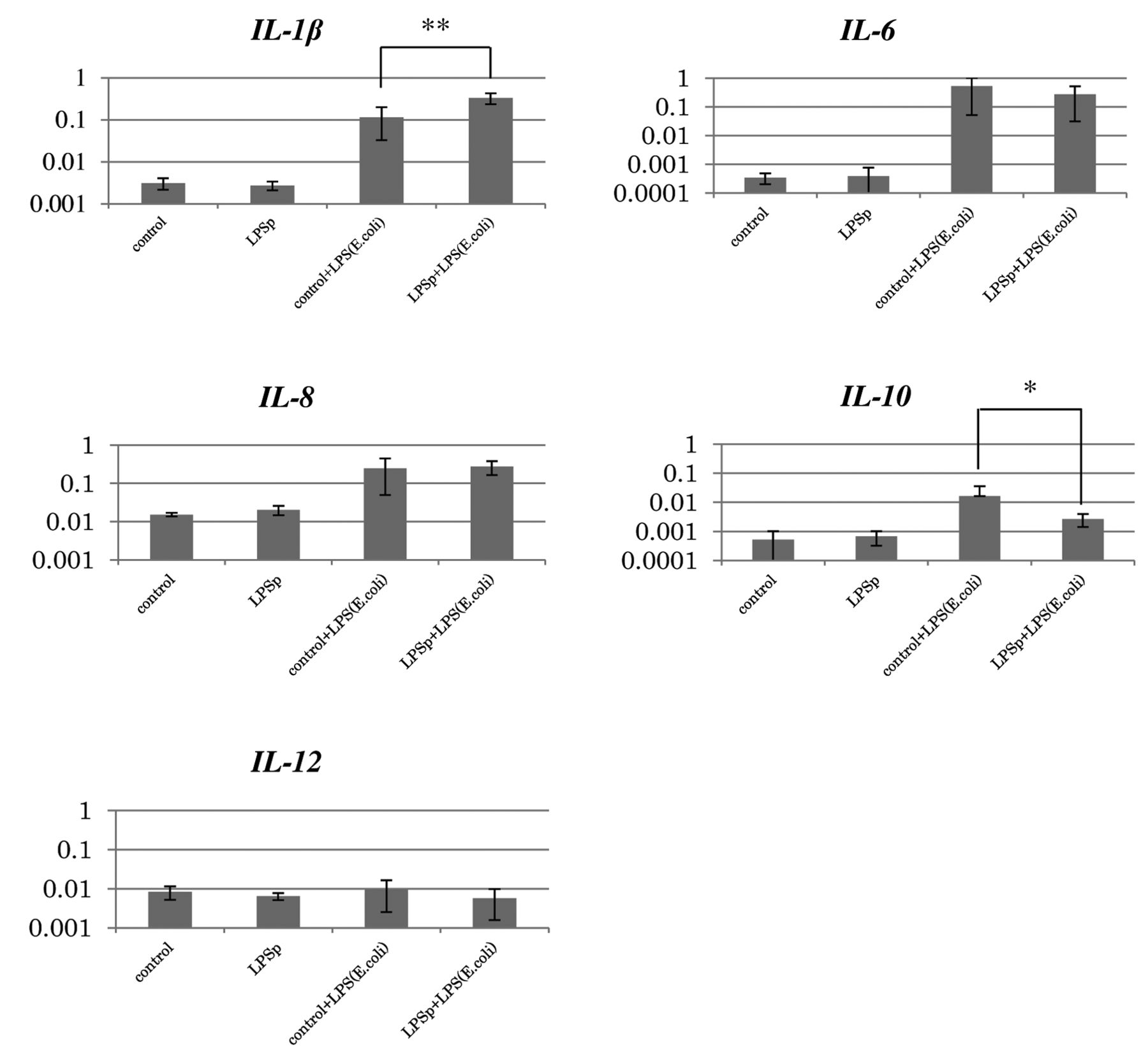
Gene expression level in the spleen after oral administration of LPSp. Female Ross 308 broiler chickens were administered feed containing LPSp (10 μg/kg BW/day) from 10 to 34 days of age. At 34 days of age, the chickens were intraperitoneally administered LPSe at a dose of 1 mg/kg BW as a trigger. Then, 2 h later, samples were collected from the spleen. Expression of each cytokine (IL-1β, IL-6, IL-8, IL-10, and IL-12) was measured by qRT-PCR. Five birds were used for each experimental plot. Statistical analysis was performed by the Dunnett test. Significant differences (**p<0.05, *p<0.1) were obtained between control+LPS (E. coli) and LPSp+LPS (E. coli).
Statistical analysis. Statistical analysis was performed using Excel 2008 ver.1.07. The Dunnett's multiple comparison test was used to compare relative gene expression levels.
Results
Prevention of mortality in broiler chicken farms by oral administration of LPSp. The results showing the number of deaths from 0 to 10 weeks of age are presented in Table II. In the 2 tests, mortality was significantly lower in the LPSp administration group with 67 and 69 deaths (162 and 155 in the control group) (p<0.01). Furthermore, it may be noted that mortality is commonly caused by sudden death syndrome (SDS) (19) and leg disorder (leg weakness) (20).
Summary of primer sequences used for chicken PCR analysis.
Enhanced phagocytic activity in broiler peripheral blood monocytes by oral administration of LPSp. The results of phagocytic activity in broiler peripheral blood monocytes evaluated after 3 weeks of age is shown in Table III. With LPSp administered at a dose of 200 μg/kg, the phagocytic activity was significantly increased at 6 weeks of age (p<0.01).
Enhanced NO production in broiler peripheral blood monocytes by oral administration of LPSp. The transition in NO-producing ability with or without stimulation with killed E. coli is shown in Table IV. There was no difference observed related to the dose in the non-stimulation group. However, similar to phagocytic activity, NO production was significantly increased at 6 weeks of age in the stimulation group when LPSp was administered at a dose of 200 μg/kg (p<0.01).
Analysis of gene expression level in the spleen after oral administration of LPSp and stimulation with killed E. coli. Cytokine mRNA expression levels in the spleen according to the presence or absence of intraperitoneal LPSe administration are shown in Figure 1. We found that intraperitoneal LPSp administration induced significant changes in IL-1β, IL-6, IL-8 and IL-10 gene expression. IL-12 gene expression was not affected. Furthermore, the LPSp-administration group exhibited significantly higher IL-1β expression compared with the group stimulated using LPSe (p<0.05), whereas IL-10 tended to decrease (p<0.1). No significant changes in IL-6, IL-8 or IL-12 were observed.
Mortality of Ross 308 broiler chickens on commercial farms. Broilers were randomly divided into 2 groups and administered feed containing LPSp (10 μg/kg BW/day) from 3 to 7 weeks of age compared with controls. Thereafter, 2 tests were performed to examine broiler mortality.
Discussion
In the present study, we evaluated whether oral administration of LPSp induced the same breeding results and priming effect in chickens (Ross 308 broiler) as seen in mice on the basis of phagocytic activity in peripheral blood monocytes, NO-producing ability and cytokine mRNA expression in the spleen. We found that oral administration of LPSp significantly reduced mortality in Ross 308 broiler chickens (p<0.01), whereas phagocytic activity and NO-producing ability were significantly increased upon stimulation with killed E. coli (p<0.01).
It has been found that both phagocytic activity and NO production are enhanced by priming induced by administration of interferon-γ in mice; in the present study, we confirmed that priming is induced by oral administration of LPS even in birds. It appears that enhanced phagocytosis effectively removes pathogens that have invaded the body, which aids in prevention of infection. This is supported by our observation that oral administration of LPS reduced broiler mortality.
Oral administration of LPS has been found to enhance phagocytic activity of hemocytes in invertebrates, such as kuruma prawn (Penaeus japonicus) (13), and fish, such as carp (Cyprinus carpio) (21). The fact that enhanced phagocytic activity was observed in birds also suggests that it is a function that has been preserved in higher-order animals. With regard to dosing, we observed these effects with an oral dose of 200 μg/kg of LPS for 7 days. On the other hand, enhanced phagocytosis was observed in kuruma shrimp at a dose of 20-40 μg/kg BW for 7 days and in carp at a dose of 10 μg/kg BW for 7 days. Although the optimal dose has not been examined for chickens, the fact that infection was prevented at a dose of 10 μg/kg BW suggests that phagocytic activity is enhanced at a dose of several μg/kg BW for 7 days. However, this should be examined further.
Phagocytic activity of peripheral blood monocytes after LPSp administration in female Ross 308 broilers. LPSp (200 μg/kg or 0.2 μg/kg) was force-fed to broiler chickens from 3 to 4 weeks of age. Thereafter, peripheral blood monocytes were collected and the rate of phagocytosis was measured under a microscope using fluorescent latex beads. Five birds were used for each test.
NO-producing ability of peripheral blood monocytes after LPSp administration in female Ross 308 broiler chickens. LPSp (200 μg/kg or 0.2 μg/kg) was force-fed to broiler chickens from 3 to 4 weeks of age. Thereafter, peripheral blood monocytes were collected and killed E. coli was added as a trigger. Then, 24 h later, nitrite ion concentration in the supernatants was measured using the Griess reagent assay. Five birds were used for each test.
Macrophages are broadly divided into M1 type (conventional activation) and M2 type (complement activation) (22). In our analysis of cytokines in the spleen, we evaluated mRNA expression in 3 types of M1 cytokines (IL-1β, IL-8 and IL-12), and 2 types of M2 cytokines (IL-6 and IL-10). Although no difference was observed in gene expression in terms of dose on the basis of LPSp oral administration alone, intraperitoneal stimulation with LPSe significantly increased the expression of the M1 cytokine IL-1β (p<0.05) and reduced the expression of the M2 cytokine IL-10 (p<0.1). It is commonly known that M1 macrophages defend against infection upon acute infection (22). This suggests that administration of LPSp may have strongly activated M1 macrophages, thus inducing priming that increases reactivity by means of secondary stimulation.
The main role of innate immunity involves stimulation of phagocytes including neutrophils, macrophages and dendritic cells, which utilize signals from toll-like receptors (TLRs) to differentiate pathogens and self-antigens (23). TLRs induce reactive oxygen and nitrogen intermediates (ROI and RNI), as well as inflammatory cytokine production, which increases the expression of mutually stimulating molecules and subsequently activates the optimal immune system (24). In chickens, 10 TLRs have been identified; among these, TLRs 3, 4, 5 and 7 are directly orthologous to those found in other vertebrates (24). TLR4 is known as a specific receptor of LPS derived from Gram-negative bacteria (25). Further examination is needed, including analysis of the expression of immune-related molecules in chickens; however, the priming effect of LPSp administration confirmed in the present study is assumed to be mediated through TLR4 and is a promising finding that could be applied in the activation of innate immunity and infection prevention in broiler chickens from commercial farms.
Footnotes
Conflicts of Interest
The Authors have no financial conflicts of interest.
- Received April 3, 2015.
- Revision received May 13, 2015.
- Accepted May 14, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}