


Abstract
Background/Aim: Neuroblastoma (NB) is the most common and lethal extracranial solid tumor in children. The present study aimed to verify that the heparin-binding epidermal growth factor-like growth factor (HB-EGF) is a rational target in NB therapy. Material and Methods: We examined expression of EGFR ligands in four NB cell lines using 2-dimensional culture (DC) and 3DC conditions. To assess the anti-tumor effect of cross-reacting material 197 (CRM197), which is a specific inhibitor of HB-EGF, on NB cells, we also performed terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay to detect apoptotic cells. Results: HB-EGF was predominantly expressed in two out of four NB cell lines under 2DC and 3DC conditions. CRM197 significantly induced apoptosis of NB cells with high HB-EGF expression. Conclusion: HB-EGF plays an important role in neuroblastoma tumorigenesis and CRM197 showed an effective antitumor effect in neuroblastoma cells.
Neuroblastoma (NB) is a neuroblastic tumor derived from the primordial neural crest. It is the most common solid malignant extracranial tumor in children and infants accounting for 8-10% of all diagnosed paediatric malignancies and approximately 15% of paediatric cancer deaths (1). Stage, age, histology, MYCN gene status, changes in chromosome 11q and DNA ploidy in tumor cells are used to categorize NB as very low-risk, low-risk, intermediate-risk or high-risk. Although some tumors have good prognoses, about half of NB tumors are very aggressive. In particular, high-risk NB has a 5-year event-free survival rate of less than 50% (2).
NB reportedly shows several heterogeneous clinical behaviours (3). Some NB tumors can spontaneously regress or mature, even without therapy; others display a very aggressive phenotype that responds poorly to current multimodal therapy. New strategies and rapid development of targeted-therapies are needed to improve patient outcomes. Although several recent reports have associated the clinical behaviour of high-risk NB with some molecular factors (4-6), these studies lack clear evidence.
Heparin-binding epidermal growth factor-like growth factor (HB-EGF) is a member of the epidermal growth factor (EGF) superfamily produced mainly by activated macrophages, endothelial cells and smooth muscle cells (7-9). Membrane-anchored HB-EGF (proHB-EGF) is synthesized as an 87-amino acid chain that includes an EGF-like domain (10, 11). It is cleaved by proteases from the disintegrin and metalloproteinase families (12); membrane type 1-matrix metalloproteinase (13) affects this cleavage at the cell surface in a process called ectodomain shedding, which induces secretion of the soluble mature form of HB-EGF (sHB-EGF) (14, 15). sHB-EGF binds to the EGF receptor (EGFR) and can contribute to several biological processes, including wound healing (16), inflammation (17, 18) and carcinogenesis (19). Notably, sHB-EGF is essential for the development of some tumors, including ovarian (19, 20), gastric and breast cancer (21-24). In a previous report, we found HB-EGF to be a potential therapeutic target for those cancers (21).
ProHB-EGF also functions as a receptor of diphtheria toxin. Cross-reacting material 197 (CRM197) is a non-toxic mutant of diphtheria toxin that specifically binds to the EGF-like domain of both proHB-EGF and sHB-EGF (24), thus inhibiting binding of HB-EGF to EGFR and downstream mitogenic signalling by the HB-EGF pathway (25). CRM197 is a specific inhibitor of HB-EGF; it does not bind other EGFR ligands or inhibit their mitogenic activity (26). CRM197 is currently under evaluation in clinical cancer trials. The present study aimed to evaluate the validity of HB-EGF as a therapeutic target molecule for NB and the in vitro antitumor effect of CRM197 on NB cells.
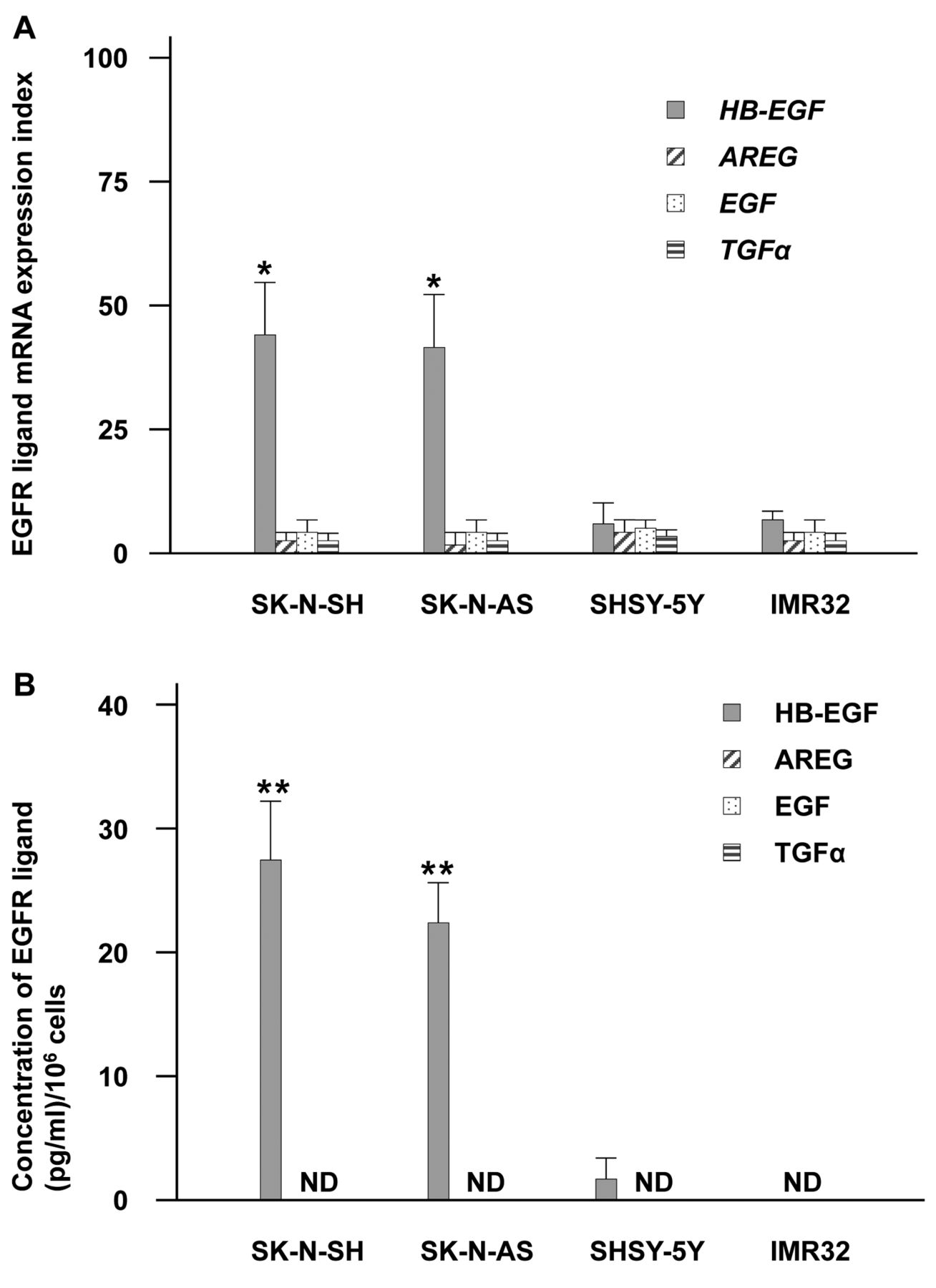
Differences in EGFR ligand expression in neuroblastoma cells under 2-dimensional culture (DC). (A) EGFR ligand mRNA expression indices (with mean and SD) in neuroblastoma cells. *p<0.05, versus other EGFR ligands. (B) EGFR ligand protein concentrations per 1×106 cells (with mean and SD) in 2DC media from neuroblastoma cells. **p<0.05, versus SHSY-5Y and IMR32 cells. ND: Not determined. For both panels, heparin-binding epidermal growth factor-like growth factor (HB-EGF): grey bar; amphiregulin (AREG): hatched bar; epidermal growth factor (EGF): dotted bar; transforming growth factor-alpha (TGFα): striped bar.
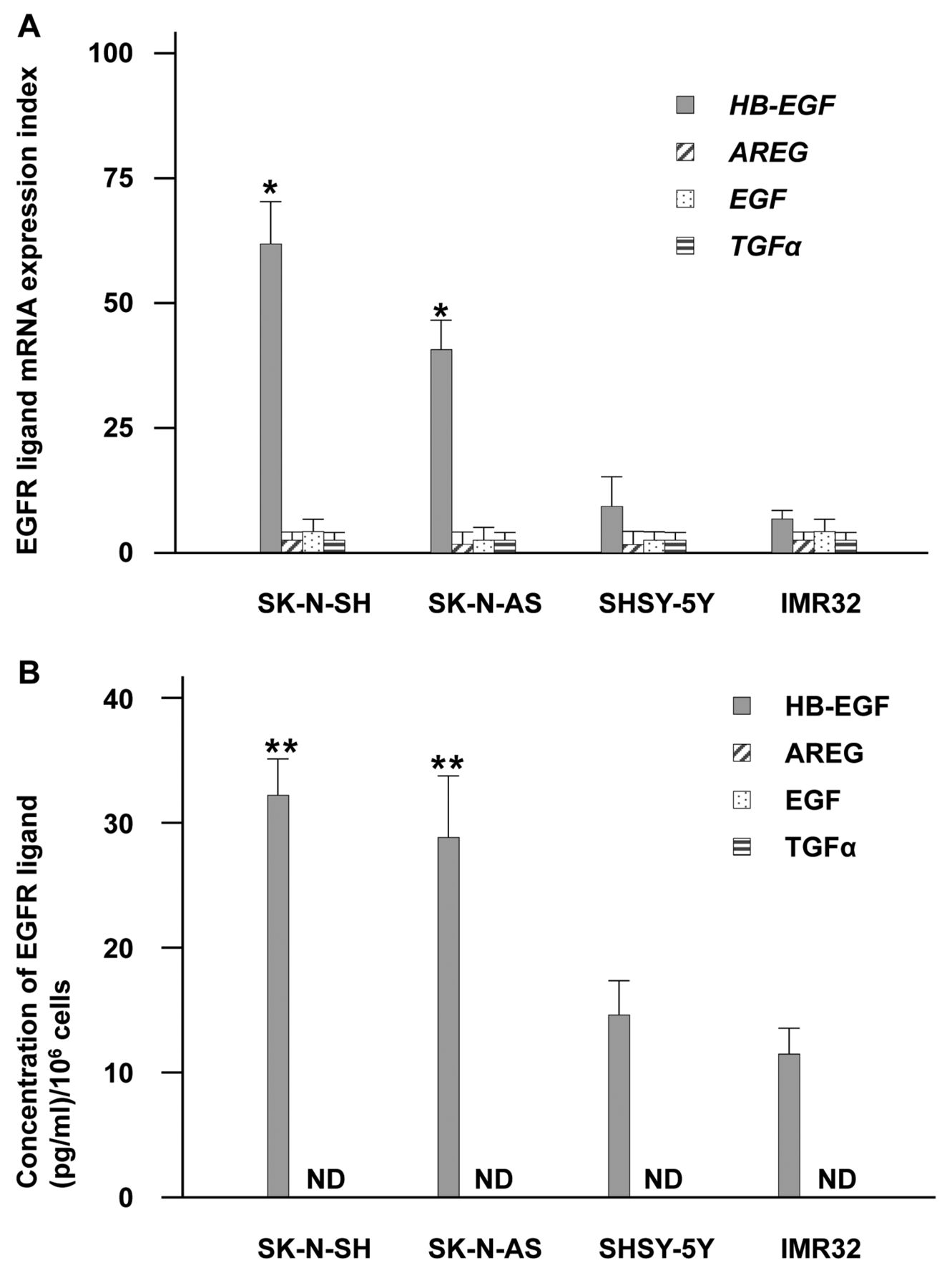
Differences in EGFR ligands in neuroblastoma cells under 3DC conditions. (A) mRNA expression (with mean and SD) in neuroblastoma cells.*p<0.05 versus other EGFR ligands. (B) EGFR ligand protein concentrations per 1×106 cells (with mean and SD) in 3DC media from neuroblastoma cells. **p<0.05, versus SHSY-5Y and IMR32 cells. ND: not determined. For both panels HB-EGF: grey bar; AREG: hatched bar; EGF: dotted bar; TGF-α: striped bar.
Materials and Methods
Cells and cell culture. Human NB cell lines, SK-N-SH, SK-N-AS, SH-SY5Y and IMR32, were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). SK-N-SH and IMR32 cells were maintained in Eagle's minimal essential medium (EMEM; ATCC, Manassas, VA, USA), SK-N-AS cells were maintained in Dulbecco's modified Eagle's medium (DMEM; ATCC, Manassas, VA, USA). SH-SY5Y cells were cultured with 1:1 mixture of EMEM and F12 Medium (Life technologies, Carlsbad, CA, USA). All media contained 10% foetal bovine serum (ICN Biomedicals, Irvine, CA, USA), 100 U/ml penicillin G and 100 μg/ml streptomycin (Invitrogen Corp., Carlsbad, CA, USA). Cultures were carried-out in a humidified atmosphere of 5% CO2 at 37°C.

HB-EGF mRNA and protein expression associated with the neuroblastoma tumorigenesis. (A) HB-EGF mRNA in SK-N-SH and SK-N-AS cell lines under 2DC or 3DC; 2DC: dotted bar; 3DC: grey bar. (B) ELISA shows amounts of EGFR ligand protein in culture media from SK-N-SH and SK-N-AS cell lines under 2DC or 3DC; 2D: dotted bar; 3DC: grey bar. For all panels: *p<0.05 vs. 2DC; data show mean and SD from triplicate experiments.
Reagents. CRM197 was obtained from the Research Institute for Microbial Diseases, Osaka University (Osaka, Japan).
Three-dimensional culture (3DC). Cultured cells were detached with trypsin-EDTA, washed three times with serum-free medium and suspended at a final concentration of 2×105 cells/1.5 ml. Aliquots (1.5 ml) were applied to the wells of 6-well plates precoated with 1.5 ml/well Matrigel (Becton Dickinson, Franklin Lakes, NJ, USA). Cells were then cultured under each experimental condition. Cells were retrieved from colonies using a BD Cell Recovery Solution (Becton Dickinson).
Real-time quantitative polymerase chain reaction (PCR). RNA extraction was performed using Trizol (Invitrogen); first-strand cDNAs were synthesized from equal amounts of total RNA (1 μg/reaction) with a PrimeScript II first-strand cDNA synthesis kit (Takara Bio, Shiga, Japan) each in a total volume of 20 μl as described by the manufacturer's protocol. Synthesized cDNAs were used for real-time PCR, that was performed using the Applied Biosystems 7500 Real-Time PCR Systems (Applied Biosystems, Foster City, CA, USA). The TaqMan quantitative PCR was carried-out using primer pairs; TaqMan probes for each EGFR ligand and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were detected with Assays-on-Demand primer and probe sets were HB-EGF: Hs00181813_m1; amphiregulin (AREG): Hs00950669_m1; transforming growth factor alpha (TGF-α): Hs00608187; epidermal growth factor (EGF): Hs01099999_m1; and GAPDH: Hs02758991_g1 (Applied Biosystems). Serial 1:10 dilutions of plasmid DNA containing each target cDNA (107-101 copies/μl) were analyzed and served as standard curves from which the rates of change for the threshold cycles were determined. Copy numbers of target cDNAs were estimated from the standard curves. Each PCR reaction was carried-out according to manufacturer's instructions. To evaluate mRNA levels, we used the mRNA expression indices, which reflect the relative mRNA expression level standardized to GAPDH and calculated as (in arbitrary units): mRNA expression index=(copy number of each mRNA/copy number of GAPDH mRNA) ×10,000 arbitrary units. All experiments were carried out independently three times.
Measurement of soluble HB-EGF, AREG, EGF and TGF-α in cell culture media. Cells were incubated for 48 h; levels of EGFR ligands in culture medium were determined using a commercially available sandwich ELISA (R&D Systems Inc., Minneapolis, MN, USA) according to manufacturer's instructions. Samples were normalized by cell numbers. Each mean value was considered representative of its corresponding culture media.
Cell apoptosis assay. NB cells (1×105) were seeded on poly-lysine-coated 6-cm dishes and their respective media then replaced with media containing 1% foetal bovine serum at 37°C for 48 h in the presence of CRM197 (10 μg/ml) after assuring the complete adherence of the cells to the poly-lysine-coated 6-cm dishes. For cell imaging, cells were fixed for 10 minutes in 4% (w/v) paraformaldehyde in phosphate-buffered saline (PBS) and photographed using a phase-contrast microscope (Leica-DM 2500; Leica, Wetzlar, Germany). For TUNEL assay, cells were harvested, pooled and fixed with 4% paraformaldehyde at 4°C for 30 min and re-suspended in 70% ethanol at −20°C for 30 min. After washing in PBS, cells were incubated with TdT reaction reagent (MEBSTAIN Apoptosis Kit Direct; MBL Co. Ltd., Nagoya, Japan) for 1 h at 37°C according to manufacturer's instructions. The TUNEL+ cells were quantified as apoptotic cells by flow cytometry analysis using a FACSCalibur (Becton-Dickinson, Franklin Lakes, NJ, USA).

Flow cytometry of apoptotic cells in neuroblastoma. Rate differences for apoptotic (A) SK-N-SH cells (A) and IMR32 cells (B) after treatment with CRM197 (grey line); control: black line. Each histogram represents three independent experiments. (C) Alteration in the percentage of apoptotic cells after incubation with the CRM197. Bars: Mean values and standard errors. *p<0.05 vs. control.
Statistical analysis. The statistical significance of differences among the groups was assessed using the Mann-Whitney U-test. All p-values <0.05 were considered significant.
Results
Validation of HB-EGF as a target for NB cancer cells. To address the validity of HB-EGF as a target for NB therapy, we examined mRNA expression and supernatant protein levels of EGFR ligands from NB cell lines (SK-N-SH, SK-N-AS, SHSY-5Y and IMR32) under 2DC conditions. SK-N-SH and SK-N-AS cells had greater HB-EGF mRNA (Figure 1A). In addition, SK-N-SH and SK-N-AS cells showed prominent secretion of HB-EGF protein in 2DC medium supernatant, although other EGFR ligands were not determined (Figure 1B). Meanwhile, SHSY-5Y and IMR32 cells did not show significant greater expression of HB-EGF mRNA expression compared to other ligands (Figure 1A) nor soluble HB-EGF protein expressions (Figure 1B). These results imply that HB-EGF is a valid molecular target for NB therapy.
Significance of HB-EGF in NB tumorigenesis. To determine whether HB-EGF contributes to NB cell proliferation in three-dimensional microenvironments, we tested HB-EGF mRNA and protein expression levels of the four cell lines in 3DC condition. As with 2DC, HB-EGF protein and HB-EGF mRNA were dominantly expressed in SK-N-SH and SK-N-AS cells and much more abundantly than in SHSY-5Y and IMR32 cells (Figure 2A, B). We also saw no significant change in EGFR ligand mRNA expression in SHSY-5Y and IMR32 cells and could detect only secreted HB-EGF protein by ELISA (Figure 2B). Compared to the 2DC results (Figures 1 and 2), HB-EGF mRNA and protein HB-EGF expression were both significantly enhanced in SK-N-SH and SK-N-AS cells under 3DC (Figure 3A, B). These data indicate that HB-EGF has a central function in NB tumorigenesis compared to other EGFR ligands.
Neuroblastoma cell apoptosis mediated by CRM197. To ascertain whether HB-EGF could be a therapeutic target for NB, we investigated the in vitro antitumor effects of CRM197 on NB cells using the TUNEL assay. CRM197 caused significant apoptosis in SK-N-SH cells (Figure 4A) but not in IMR32 cells with low HB-EGF expression (Figure 4B). Percentages of apoptotic cells were SK-N-SH: 43.3±4.6%; SK-N-AS: 38.4±5.2%; SHSY-5Y: 3.2±4.6%; and IMR32: 5.6±2.2% (Figure 4C). Phase-contrast microscopy also showed SK-N-SH cells treated with CRM197 to form apoptotic bodies (Figure 5A). In contrast, IMR32 cell morphology was almost unchanged by CRM197 treatment (Figure 5B). These findings imply that CRM197 could be effective against HB-EGF as an NB therapeutic target.
Discussion
In the present study, we confirmed that HB-EGF is the predominant EGFR ligand in NB cells and essential for their three-dimensional proliferation. We also found that inhibiting HB-EGF expression by CRM197 had a tumor-suppressing effect in vitro.

Photomicrographs of SK-N-SH cells (A) and IMR32 cells (B) treated with CRM197. Left panels: control; Right panels: CRM197.
Chemotherapy combinations, including cisplatin, cyclophosphamide, vincristine, doxorubicin and etoposide, as well as surgery, followed by myeloablative therapy using hematopoietic stem cells, brought the best outcome in high-risk NB (27). However, the overall survival of the high-risk group of NB remained poor. Recently, novel therapeutic approaches for PI3 kinase, mTOR, IGF1R and other pathways that are implicated in NB proliferation have been used in clinical trials. Other specific inhibitors for NB, including 131I-metaiodobenzylguandine (targets the norepinephrine transporter, expressed in 90% of NB) (28), anti-GD2 antibody (targets GD2 ganglioside, expressed on >98% of NB) (29) and ALK small molecule inhibitor (targets ALK gene, which is mutated in 8-12% of NB at diagnosis) (30, 31) have emerged in pre-clinical studies and early-phase clinical trials for NB. As these agents are effective for only certain NB types, development of therapies for high-risk NB is still needed. Our results herein show that HB-EGF is a potential target in NB and that CRM197 could be useful in NB therapy. Previously, we reported that combination chemotherapy of CRM197 with conventional agents, such as paclitaxel and irinotecan, had a synergistic anti-tumor effect on ovarian and gastric cancers (32-34). Along with our current results, they suggest that adding CRM197 to chemotherapeutic agents could improve clinical outcomes of high-risk NB patients.
Neuroblastoma has heterogeneous properties meaning that tumors can spontaneously regress or mature into extremely malignant, chemotherapy-resistant cancers. Many factors responsible for this heterogeneity have been identified; they indicate that NB's biological and molecular features are highly predictive of clinical behaviour. Previous reports suggest that the most important biological factors were MYCN copy number (35, 36), DNA content (ploidy) (37, 38) and deletion or loss of heterozygosity of 1p and gain of 17q. However, identifying the most useful of these prognostic markers is difficult because NB's clinical behaviour is complex and large prospective clinical studies are lacking. Aside from amplification of the MYCN gene, risk classification for these factors is not confirmed. The overall prevalence of MYCN amplification is about 22% in NB (39) and thought to be a powerful predictor of a poor prognosis. However, whether MYCN mRNA or MYCN protein has prognostic significance in tumors is controversial as high levels of MYCN mRNA or MYCN protein show no relation to MYCN gene amplification in some NB cell lines as some NB cells lack MYCN amplification (40, 41). Therefore, development of a reliable prognostic and biologic marker for NB is needed to guide judgement of appropriate chemotherapy, as well as timing and extent of operative procedures for NB. We previously reported that high serum HB-EGF was associated with poor prognosis in ovarian cancer and triple-negative breast cancer (42, 43). In this study, SK-N-SH and SK-N-AS cells (derived from metastatic NB tumors) showed an increasing expression of HB-EGF. We, therefore, considered that HB-EGF could be a marker for poor prognosis in NB.
In conclusion, we verified that the HB-EGF is a key molecule in NB tumorigenesis and could be a target for NB therapy. We had previously shown, by a phase I study in the Fukuoka University Hospital (Japan), under the approval of the Institutional Ethical Committee, that CRM197 can be safely used in patients with recurrent ovarian cancer. The development of an HB-EGF inhibitor, such as CRM197, to target neuroblastoma would be a logical next step leading to better clinical outcomes for young NB patients.
- Received April 3, 2015.
- Revision received May 10, 2015.
- Accepted May 12, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}