


Abstract
Aim: The purpose of the present study was to compare microarray gene-expression profiling data between primary central nervous system (CNS) lymphoma and non-CNS lymphomas. Materials and Methods: We performed whole-genomic cDNA-mediated annealing, selection and ligation assay with 177 formalin-fixed paraffin-embedded tumor samples. Results: We identified 20 differentially expressed genes out of which 5 were predominantly expressed in CNS DLBCL compared to non-CNS DLBCL (C16orf59, SLC16A9, HPDL, SPP1, and MAG). SLC16A9 may be involved in aerobic glycolysis of malignant tumors. The alteration in gene expression of SPP1 in primary CNS lymphoma is involved in biological activity, such as CNS tropism, B-cell migration, proliferation, and aggressive clinical behavior. MAG may be an important adhesion molecule that contributes to perineural cancer invasion. Conclusion: Genomic differences between CNS and non-CNS DLBCL exist and the most prominent genes are SPP1 and MAG. SPP1 may play a key role in CNS tropism of primary CNS lymphoma.
Primary central nervous system (CNS) lymphoma refers to lymphoma presenting in the brain parenchyma, spinal cord, eyes, and cranial nerves in the absence of systemic lymphoma (1). Primary CNS lymphoma is rare and currently estimated to account for up to 1% of non-Hodgkin's lymphoma (2, 3). Morphologically, approximately 95% of these tumors are diffuse large B-cell lymphomas (DLBCL) categorized as high-grade non-Hodgkin's type, with only a small proportion comprising Burkitt's, lymphoblastic, marginal zone or T-cell lymphoma (2-5). However, primary CNS lymphoma has clinical features distinct from those of DLBCL, as well as worse prognosis than non-CNS DLBCL. High-dose methotrexate is the most active chemotherapeutic agent for PCNSL and is usually used as a single agent or as the backbone of combination chemotherapy (3-6). Several other chemotherapeutic agents have been studied in combination with high-dose methotrexate, but standard treatment of PCNSL has not yet been defined.
Since the improvement of detailed analysis of molecular pathogenesis, a variety of molecular and genetic abnormalities of DLBCL have been recognized (7). DLBCL has a wide range of clinical courses and prognoses within the same clinical stage, depending on the molecular and genetic abnormalities of each case (8-10). Previous studies have identified distinct sub-types of DLBCL based on their gene-expression profiling. Patients with germinal center B-cell-like DLBCL had significant by better survival than the of the activated B-cell-like subgroup (11, 12). During the past decade, research has also focused on molecular pathogenesis of primary CNS lymphoma. Studies of gene-expression profiling of primary CNS lymphoma are scarce owing to limitations of biopsy size, and the results were conflicting and differed between the various studies, mostly because of the differences in platforms and algorithms (13-17).
Pathogenesis, biology and molecular knowledge of primary CNS lymphoma are, therefore, still poorly understood regardless of the phenotypic similarities between CNS and non-CNS lymphoma. Thus, in the present study, we used formalin-fixed paraffin-embedded (FFPE) tumor samples for microarray gene-expression profiling to compare CNS and non-CNS lymphomas and identify specific differentially expressed genes which may be helpful to understand molecular pathogenesis and potential treatment targets of primary CNS lymphoma.
Materials and Methods
Patient cohort. We performed a retrospective analysis using tumor tissue taken at diagnosis. The purpose of this study was to use gene-expression profiles to identify the difference between CNS and non-CNS DLBCL. We collected data of all patients diagnosed with DLBCL at the Samsung Medical Center, Seoul, Korea from August 1995 to November 2009. All available tissue slides were centrally reviewed by a hematopathologist (YHK) and primary tumor blocks containing the largest amount of tumor were selected for this study. The immunohistochemistry (IHC) was performed for the diagnostic work-up for DLBCL including for CD10, Ki-67, BCL2, BCL6, and MUM1 at the time of initial diagnosis. The classification of germinal center type and non-germinal center type was carried-out according to a previously reported algorithm (18). Tumor samples were obtained by stereotactic biopsy or open craniotomy in primary CNS lymphoma and all tumor blocks of non-CNS DLBCL were obtained during excisional biopsy from involved lymph nodes or surgical resection of tumor from extranodal sites such as the intestine or spleen. Tissue blocks with poor quality were excluded from this study as were DLBCLs which had transformed from other types or post-transplantation lymphoma. This study was approved by the Institutional Review Board (2011-07-046) at the Samsung Medical Center.
Microarray gene-expression profiling. Total RNA was extracted from two to four sections of 4-μm thick FFPE sections using the High Pure RNA Paraffin kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's protocol. The FFPE sections contained a minimal 70% of tumor volume. Samples with RNA concentrations of <40 ng/μl, and absorbance A260/A280 ratios of <1.5 or A260/A230 ratios of <1.0 were considered inadequate and were excluded from the analysis. The whole-genome cDNA-mediated annealing, selection and ligation (WG-DASL) (Illumina, San Diego, CA, USA) assay was performed following the manufacturer's instructions using 200 ng RNA. We excluded samples based on the following quality control criteria: number of genes detected <4,905, gene intensity corresponding to the 95th percentile <1,000 and gene intensity corresponding to the 75th percentile <500. The quality control criteria were from our previous experience with WG-DASL assay of stomach cancer samples after their value was validated by the comparison of the results of WG-DASL assay with results of fluorescence in situ hybridization (FISH) and IHC. A total of 24,526 probes on the array were analyzed, we filtered out probes that were confirmed as “present” in fewer than 20% of the samples. Finally, filtering yielded 16,393 probes to be used in the subsequent analyses. The probe intensity was log2-transformed and then substituted after quantile normalization (19).
Statistical analysis. A two-sample t-test was used for analysis of comparing CNS with non-CNS DLBCL. A volcano plot was drawn using the negative log of the p-value of diferentially expressed genes between two group (CNS vs. non-CNS) and log of the fold difference for comparison of gene expressions. Rather than using a fold-change cut-off value to identify differentially expressed genes, we used the fold difference and false-discovery rate of the p-value (Q value) performed with Bonferroni correction on the p-value; a gene with a Q-value of <0.0001 and fold difference >2 was defined as a differentially expressed gene (DEG) in this study. Overall survival was estimated using a Kaplan–Meier curve.
Results
Patients' characteristics. A total of 177 samples (CNS DLBCL=10, non-CNS-DLBCL=167) were analyzed with WG-DASL assay, after quality control in this study (Table I). The baseline characteristics of the patients with non-CNS DLBCL were previously reported in a DASL study for prediction of treatment outcome of DLBCL (20). The patient characteristics of CNS DLBCL are summarized in Table II. The median age was 56.5 years (range=52-72 years), and there were five males. The majority of patients were in the low- or low-intermediate risk group, according to the international prognostic index (IPI) (80%). Among the 10 patients, seven were classified by IHC staining of previous reported model (18) all with non-germinal center type. Seven patients had died at the time of analysis, and median overall survival was 13.2 months (95% confidence interval=2.8-23.6 months).
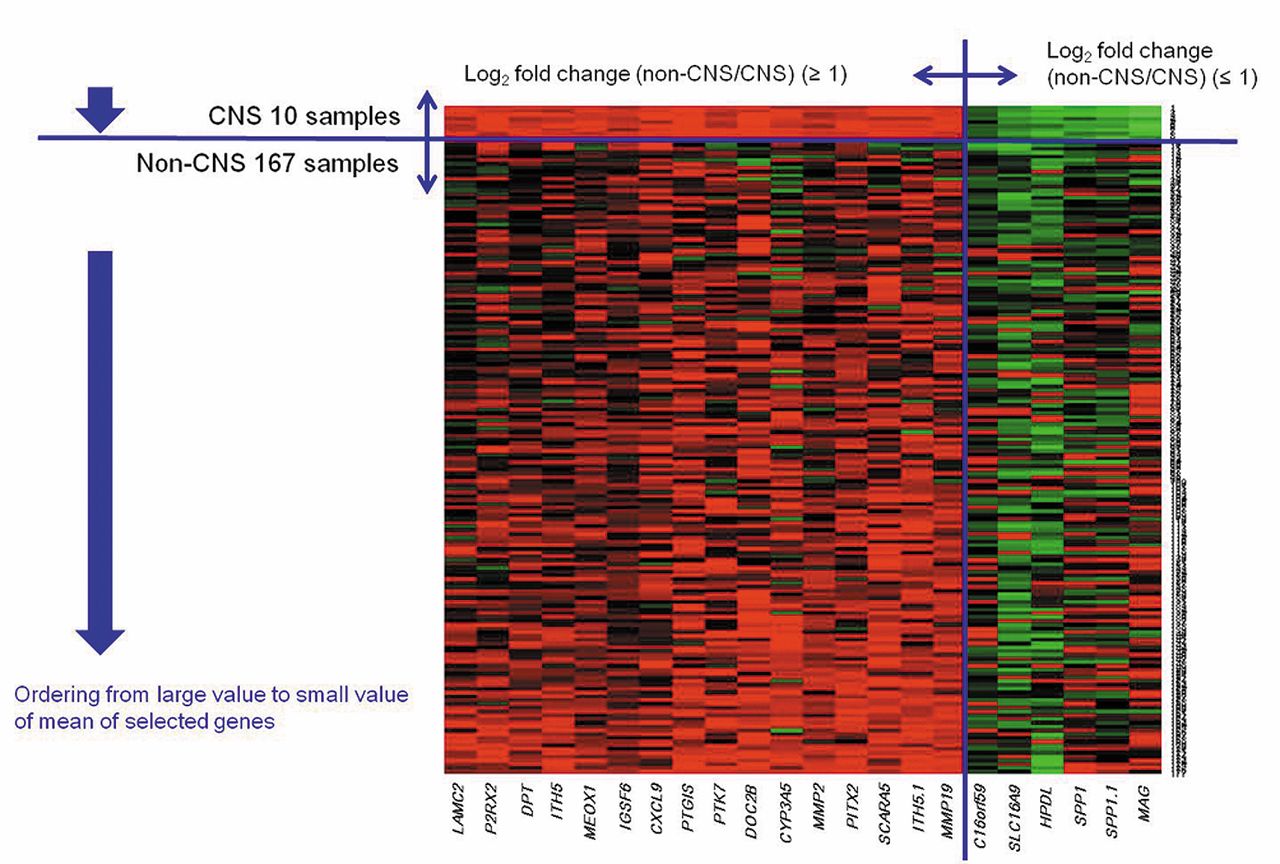
Gene expression profiles. Among of a total of 16,393 probes, the number of significant probes according to p- and Q-values after a two-sample t-test are shown in Table III. There are some interesting genes which were differentially expressed between CNS and non-CNS DLBCL, as shown by the volcano plot (Figure 1). According to definition of DEG, we finally selected 20 genes using 22 probes (Table IV). Among them, 15 genes were highly expressed in non-CNS DLBCL and five were predominantly expressed in PCNSL. SPP1 and the MAG of the differentially expressed 20 genes exhibited the most distinct fold difference. A heat map using the probes also showed differential gene-expression patterns of CNS and non-CNS DLBCL, especially in the genes predominantly expressed in CNS DLBCL (Figure 2). The left column of the heat map shows genes highly expressed in non-CNS DLBCL and the right column genes that were more predominantly expressed in CNS DLBCL. Although all 20 genes were statistical significant DEG, the heat map showed more striking differences for those which were highly expressed in CNS DLBCL.
Genes highly expressed in CNS DLBCL. The five genes which were predominantly expressed in CNS DLBCL compared to non-CNS DLBCL were as followed; chromosome 16 open reading frame 59 (C16orf59); solute carrier family 16, member 9 (SLC16A9); 4-hydroxy-phenylpyruvate dioxygenase-like (HPDL); secreted phosphoprotein 1 (SPP1); and myelin-associated glycoprotein (MAG). The C16orf59 is a protein coding gene which function is not identified yet. SLC16A9 is protein coding gene which function is proton-linked monocarboxylase transporter. HPDL, previously named glycoxalase domain-containing 1, may have dioxygenase activity. The protein encoded by SPP1 gene is involved in the attachment of osteoclasts to mineralized bone matrix. The encoded protein is secreted and binds hydroxyapatite with high affinity. SPP1 also has various functions including in inflammation, cytokine regulation, malignant transformation and cancer biology. The protein encoded by MAG gene is a type I membrane protein and member of the immunoglobulin superfamily. It is thought to be involved in the process of myelination. It is a lectin that binds to sialylated glycoconjugates and mediates certain myelin-neuron cell–cell interaction. Gene associations are summarized in Table V.
Distribution of samples used for analysis in the study.
Patient characteristics of primary central nervous system (CNS) lymphoma.
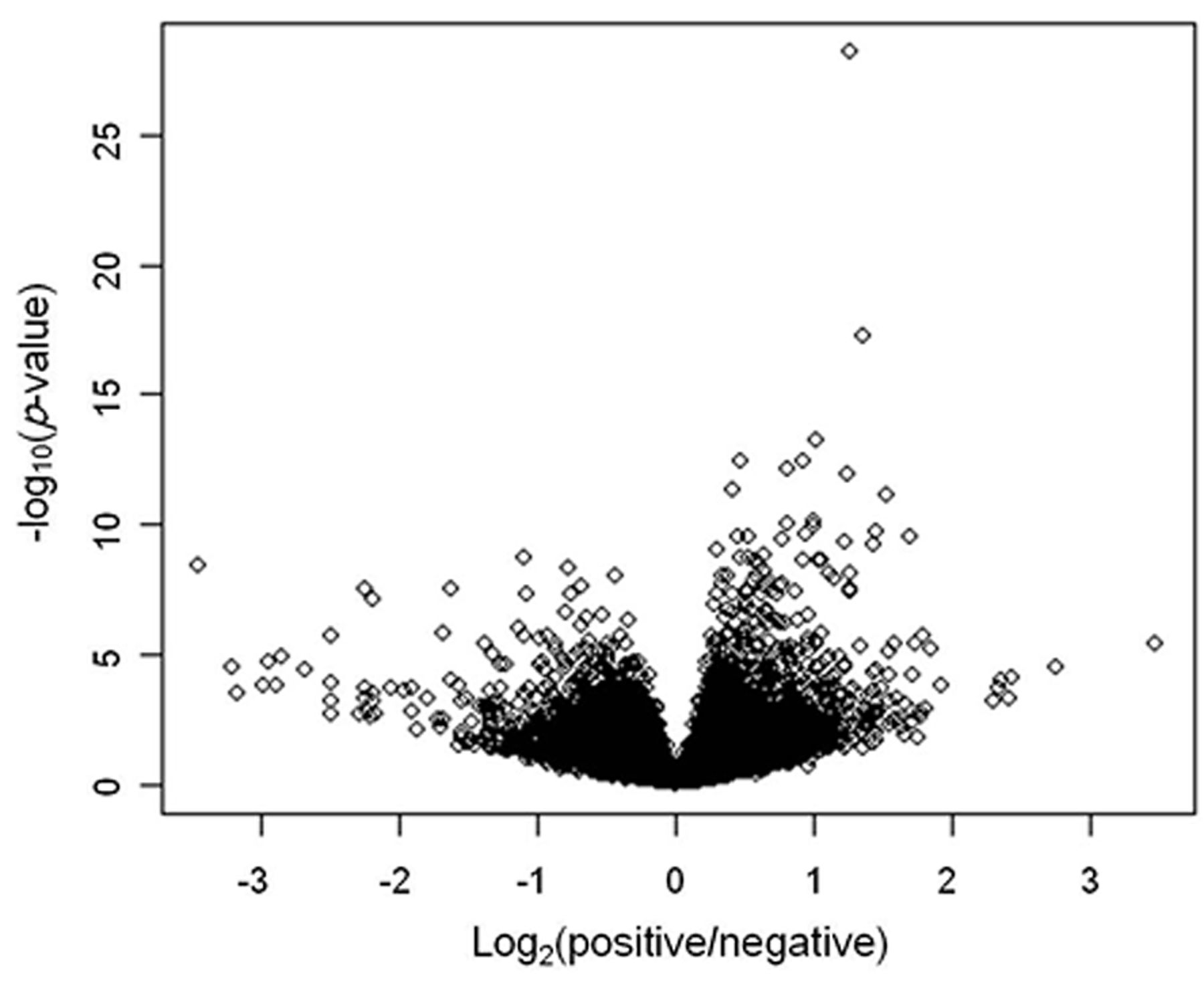
Volcano plot showing genes differentially expressed between CNS and non-CNS diffuse large B-cell lymphoma (DLBCL).
Discussion
Primary CNS lymphoma is a mature aggressive lymphoma confined to the CNS and morphologically corresponds to DLBCL. According to a gene-expression profiling study, DLBCL has been classified into three groups: germinal center B-cell type, activated B-cell type (ABC), and type 3 (11). Some studies have indicated that primary CNS lymphoma is of germinal center B-cell origin (21, 22). However, another study showed that primary CNS lymphoma has a uniform ABC-like immunophenotype which might explain its associated poor prognosis (23). These findings indicated that primary CNS lymphoma develops from a B-cell that has been exposed to a late germinal center and early post germinal center outside the CNS, which could explain the ABC phenotype observed. This suggestion was confirmed with similar results of gene-expression profiling study of primary CNS lymphoma (13) and we also found no germinal center type by IHC in our study. Therefore, understanding the mechanisms of B-cell migration and lymphoma adaptation to the microenvironment of the CNS are important goals in research on the biology of primary CNS lymphoma. At present, there exist certain molecular-based studies which compared primary CNS lymphoma with non-CNS DLBCL (14-16). A study using comparative genomic hybridization (CGH) array identified two candidate genes aberrantly expressed in primary CNS lymphoma compared to other DLBCL (14). These genes are MDM2 and YEATS4 that are involved in apoptosis, including the p53 pathway. Previous studies using microarray gene-expression profiling revealed angiotropism, extracellular matrix, and adhesion-related pathways to be involved in PCNSL (15, 16).
Analysis of results from comparison of CNS DLBCL with non-CNS DLBCL after two-sample t-test.
List of 22 probes and 20 genes selected according to the definition of a differentially expressed gene as given in the Materials and Methods section.
List of five genes predominantly expressed in CNS DLBCL compared to non-CNS DLBCL.
In the present study, we conducted gene-expression profiling of 177 FFPE tumor specimens from patients with DLBCL (CNS DLBCL=10, non-CNS-DLBCL=167) and identified alterations in gene-expression signature of primary CNS lymphoma in that five genes are predominantly expressed in CNS DLBCL compared with non-CNS DLBCL.

Heat map showed differential gene expression patterns between CNS and non-CNS DLBCL, especially in genes predominantly expressed in CNS DLBCL. LAMC2; Laminin subunit gamma-2 precursor, P2RX2; P2X purinoceptor 2, DPT; dermatopontin, ITIH5; inter-alpha-trypsin inhibitor heavy chain 5, MEOX1; mesenchyme homeobox 1, IGSF6; immunoglobulin superfamily, member 6, CXCL9; chemokine ligand 9, PTGIS; prostaglandin 12 synthase, PTK7; protein tyrosine kinase 7, DOC2B; double C2-like domains, beta, CYP3A5; cytochrome P450, family 3, subfamily A, polypeptide 5, MMP2; matrix metallopeptidase 2, PITX2; paired-like homeodomain 2, SCARA5; scavenger receptor class A, member 5, MMP19; matrix metallopeptidase 19, C16orf59; chromosome 16 open reading frame 59, SLC16A9; solute carrier family 16, member 9, HPDL; 4-hydroxyphenylpyruvate dioxygenase-like, SPP1; secreted phosphoprotein 1, MAG; myelin associated glycoprotein.
SLC16A9, also known as monocarboxylate transporter 9, belongs to a a family of proton-linked plasma membrane transporters (24). The monocarboxylate transporter family now comprises of 14 members, of which only the first four have been demonstrated to catalyse the proton-linked transport of metabolically important monocarboxylates such as lactate, pyruvate and ketone bodies across biological membranes. Malignant tumors rely heavily on aerobic glycolysis and thus need to efflux lactic acid via such transporters to the tumor microenvironment to maintain a robust glycolytic flux and to prevent poisoning themselves (25). These transporters have been successfully targeted in pre-clinical studies in brain tumors using small-interfering RNA and small molecule inhibitor (26, 27). Although the function of the SLC16A9 in primary CNS lymphoma still await characterization, this may be a potential treatment target.
SPP1, also called osteopontin, is a secreted non-collagenous, chemokine-like glycoprotein and has numerous cellular functions, including cell communication, focal adhesion, immune cell activation, and immune cell migration (15). SPP1 also has an important role in malignant transformation and is involved in various aspects of cancer biology, including cellular proliferation, cell adhesion, inhibition of apoptosis, invasiveness, metastatic capacity, regulation of cytokine expression, and angiogenesis (28-41). Many human malignant cells express SPP1 and a high level of expression of SPP1 has been associated with aggressive disease and poor prognosis in human cancer (28, 29). Currently known cellular functions of SPP1 involved in cancer biology are summarized in Table VI. In the present study, we identified that SPP1 was highly expressed in primary CNS lymphoma compared to non-CNS DLBCL, and SPP1 might have an important role in oncogenesis and cancer progression in primary CNS lymphoma. SPP1 has been found to be highly expressed in brain tumor such as malignant glioblastoma and astrocytoma (28), and other de-myelinating diseases of the CNS such as multiple sclerosis (42). These findings suggest that SPP1 plays an important role in pathogenesis of CNS disease and primary CNS lymphoma overexpressing SPP1 might be adaptation to the microenvironment of the CNS. Indeed, the CNS does not contain tissue lymphocyte under normal conditions and lacks lymphatic vessels. Primary CNS lymphomas exclusively exist in the CNS and relapse is usually confined to the CNS. Some hypotheses explain this phenomenon (4). Firstly, B-cells having transformed in the periphery may spread throughout the body and may be eliminated outside the CNS by a specific antitumor immune response, whereas they escape from the immune response in the CNS. Secondly, after transformation, malignant B-cells may have a high affinity and tropism for the CNS, stimulating their homing to the CNS. Thirdly, non- or pre-malignant B-cells enter the CNS and by chance, a low number of B-cells may persist in the CNS and malignant transformation later. Currently, it is suggested that different inflammatory and homeostatic chemokines might contribute to B-cell tropism for the CNS (43, 44). A previous pathway analysis performed a genome-wide gene expression comparison between primary CNS lymphoma and non-CNS DLBCL, revealing that primary CNS lymphoma is characterized notably by significant differential expression of multiple extracellular matrix (ECM) and adhesion-related pathways (16). Tun et al. reported that the most significantly up-regulated gene in primary CNS lymphoma is ECM-related SPP1, which can be interpreted within several biological contexts with implications for primary CNS lymphoma, including CNS tropism (ECM and adhesion-related pathway), lymphoproliferation, aggressive clinical behavior, and aggressive metastatic cancer phenotype. In our study, we also identified up-regulation of SPP1 in CNS compared to non-CNS DLBCL, and SPP1 might play a key role in CNS tropism and aggressive cancer phenotype in primary CNS lymphoma. SPP1 normally acts through interaction with CD44 receptor or intergrin receptors and the same mechanisms are used by tumor cells in the process of metastasis (30). Therefore, SPP1 could be a candidate target for cancer therapy and this approach may also be applicable to primary CNS lymphoma.
Associations found for secreted phosphoprotein 1 (osteopontin) in cancer biology.
MAG is a cell membrane glycoprotein and member of the immunoglobulin superfamily. MAG regulates the interaction of myelin and axons, such as initiation of myelination and the maintenance of myelin integrity, by expression on periaxonal myelin membrane of the CNS and peripheral nervous system (45, 46). At present, studies of MAG are mainly focused on CNS regeneration and therapeutic targets in neurodegenerative disease and neurological injury. There are only limited data for MAG in patients with cancer. MAG is an important adhesion molecule which contributes to perineural invasion of pancreatic cancer (47). MUC1, a type I transmembrane mucin, is a counter receptor for MAG and increased expression of MUC1 in pancreatic cancer its found to enhance perineural invasion. Conversely, specific inhibition of MAG or MUC1 partially blocked adhesion. MAG may play another key role in CNS tropism of malignant B-cell lymphoma and may therefore be a potential candidate target for treatment of PCNSL. In this study, we observed that MAG was highly expressed in PCNSL compared with non-CNS DLBCL. Although, we included FFPE sections containing a minimal 70% of tumor volume, there was a possibility that the MAG gene signal might be from surrounding normal brain tissue not true lymphoma itself; further studies will be needed to confirm this data from pure lymphoma tissue from a large patient group. To the best of our knowledge, this is the first report of significant up-regulation of SLC16A9 and the MAG in primary CNS lymphoma. We do not yet know exactly what the cellular functions of SLC16A9 are and whether the MAG signal is truly from lymphoma itself; these may be a clue to understanding cancer biology of primary CNS lymphoma. SPP1 has an important role in malignant transformation and in cancer biology.
In conclusion, genomic differences between primary CNS lymphoma and non-CNS DLBCL do exist. We believe our results have a significant relevance for lymphoma research and the development of potential therapeutic targets to treat primary CNS lymphoma.
Acknowledgements
This study was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2014R1A2A1A11052626) and Samsung Biomedical Research Institute (Grant No. SMX1131921).
Footnotes
This article is freely accessible online.
- Received March 10, 2015.
- Revision received March 20, 2015.
- Accepted March 24, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.