


Abstract
Background/Aim: miR-21 is a common OncomiR in human cancer. The present study analyzed the distribution and expression of miR-21 in breast tumor tissues so as to examine the role of miR-21 in the carcinogenesis of breast cancer. Materials and Methods: Sixteen malignant and 10 benign breast tissue specimens were analyzed using a miRNA chromogenic in situ hybridization (CISH) assay. The locations of miR-21 CISH-positive cells in breast tissues were observed and its expression level was semi-quantified by ISH scoring. Results: Positive in situ staining of miR-21 was detected in the cytoplasm of malignant epithelial cells in most of the high-grade infiltrating ductal carcinoma specimens. miR-21-positive spindle-like cells were found to surround tumor cell islands. High miR-21 ISH scores were correlated with positive lymph node status. miR-21 expression was low in most types of benign breast tissues. Conclusion: miR-21 is a potential biomarker for breast cancer prognosis.
MicroRNAs (miRNAs) are a class of endogenous small RNAs that negatively regulate gene expression at the post-transcriptional level (1-4). In eukaryotes, miRNA-mediated gene regulation is found in many fundamental biological processes, including proliferation, differentiation, migration and apoptosis (5-7). Research has demonstrated that miRNAs play indispensable roles in disease pathogenesis (8). In recent years, miRNAs have been identified as effective diagnostic and prognostic biomarkers, and some miRNAs have become important therapeutic targets (9, 10).
Carcinogenesis is a multi-step process involving essential genetic alterations that drive the conversion of normal cells into malignant cells (11). miRNAs are involved in all critical alterations that lead to tumor formation (12, 13). miRNAs are pleiotropic gene regulators. The effect of one miRNA can be accumulated and amplified by targeting multiple genes (13). MicroRNA-21 (miR-21) is an oncomiR which is frequently up-regulated in breast cancer and many other types of human cancers (14-16). The target genes that miR-21 blocks include tropomyosin 1 (TPM1) (17), programmed cell death 4 (PDCD4) (18), phosphatase and tensin homolog (PTEN) (19), and tissue inhibitor of metalloproteinase 3 (TIMP3) (20). Blocking miR-21 inhibits tumor cell growth and migration (21). Overexpression of miR-21 induces tumor angiogenesis (22).
Gene de-regulation can result from either abnormal expression or irregular localization of miRNA. miRNAs are found in both extracellular and intracellular locations (23). It was recently reported in a renal a fibrosis study, that tubular epithelial cells can secrete miR-21 and deliver it into recipient tubules by microvesicles, where exogenous miR-21 promotes the phenotype change of the recipient by activating the serine/threonine-specific protein kinase (Akt) pathway through blocking PTEN (24). Furthermore, increasing evidence has revealed that miRNAs not only influence the process of carcinogenesis but also affect the microenvironment that promotes the progression of tumors (25, 26). It is very common that genetically divergent tumor cells co-exist within human tumors (27). However, the potential association between the heterogeneity of human tumor cells and the histological location of altered expression of miRNA is not well-understood. In breast cancer, the observations by different groups on the source cells of miR-21 overexpression are controversial; certain groups found that miR-21 was expressed in breast cancer epithelial cells (28, 29), while others found miR-21 to be predominantly expressed in cancer-associated fibroblast-like cells and myoepithelial cells (30, 31). A recent study also showed that the expression of miR-21 in tumor stroma was relevant to metastasis and cancer survival (32). miR-21-positive cells were detected occasionally in flat epithelial atypia and normal breast epithelial cells (28, 29), while the expression status of miR-21 in benign breast tissues has not been reported.
In the present study, we used an optimized chromogenic in situ hybridization (CISH) assay to investigate the expression and distribution of miR-21 in 26 formalin-fixed paraffin-embedded (FFPE) breast tissue samples representing different histological types. The distribution of cells with altered miRNA expression and their relative expression levels in these samples were analyzed. Multiple patterns of miR-21 distribution were revealed in the malignant samples.
Materials and Methods
Clinical samples. Samples in this study included FFPE sections from 16 malignant and 10 benign or atypical breast tissue specimens. These specimens were obtained from the Samaritan Hospital in Troy, NY, USA, between October 2012 and June 2014. Tissues were fixed with 10% buffered formalin and paraffin embedded according to standard protocol. Sample blocks were cut into 4 μm-thick sections and placed onto positively charged slides. Hematoxylin and eosin (H'E) staining was performed for visualization of the tissues. Seven different histological types were identified including infiltrating ductal carcinoma (IDC), infiltrating lobular carcinoma (ILC), atypical ductal hyperplasia (ADH), fibrocystic disease (FD), fibro fatty breast tissue (FBT), fibroadenoma (FA) and stromal fibrosis (SF). The tumor grade, tumor-node-metastasis (TNM) stage, and breast cancer molecular subtypes of each sample were evaluated by board-certified pathologists using clinical pathological standards (33, 34). The sample lymph node stage (N) was determined based on the TNM system (34). The clinical pathological parameters of the breast FFPE samples in this study are listed in Table I. Institutional Review Board (IRB) approvals were obtained from the IRB committees of St Peter's Health Partners (IRB# 12-45) and Albany College of Pharmacy and Health Sciences, Albany, NY, USA.
CISH probes and reagents. The double digoxigenin (DIG)-labeled miRCURY LNA™ microRNA detection probes were purchased from Exiqon, Vedbaek, Denmark. The probe sequence for miR-21 was 5’-TCAACATCAGTCTGATAAGCTA-3’. The positive control small nuclear RNA (snRNA) U6 probe had the sequence of 5’-CACGAATTTGCGTGTCATCCTT-3’. The negative control scramble probe, which represents random sequences, had the sequence of 5’-GTGTAACACGTCTATACGCCCA-3’. The saline-sodium citrate (SSC) buffer, proteinase K, sheared salmon sperm DNA, yeast t-RNA, and 10× Denhardt's solution were purchased from Life Technologies, Grand Island, NY, USA. Sheep-anti-DIG-AP and NBT/BCIP ready-to-use tablets were purchased from Roche, Indianapolis, IN, USA. Bovine serum albumin (BSA) and Nuclear Fast Red™ were obtained from Vector Laboratories (Burlingame, CA, USA).
CISH. MCF-7 and MDA-MB-231 breast cancer cells (American Type Culture Collection, Manassas, VA, USA) cultured in Nunc™ Lab-Tek™ II Chamber Slide™ (Thermo Scientific, Waltham, MA, USA) were fixed with 10% buffered formalin for 30 min at room temperature and then stored in 70% ethanol overnight. Permeable treatment was carried out by treating slides with 5 μg/ml proteinase K for 10 min at 37°C. After dehydration, cell samples are ready for hybridization. For FFPE samples of breast tissue, 4 μm-thick sections were de-paraffinized in xylene and then rehydrated through an ethanol dilution series. After being rinsed with phosphate buffered saline (PBS), slides were treated with 20 μg/ml proteinase K for 15 min at 37°C. The slides were dehydrated in serial ethanol solutions and air dried. For each sample, three hybridization reactions including 50 nM of miR-21 probe, 50 nM of scramble probe, and 5.0 nM snRNAU6 probe in 20 μl of hybridization buffer (50% formamide, 0.5 mg/ml sheared salmon sperm DNA, 0.5 mg/ml yeast t-RNA, 1× Denhardt's solution, 10% dextran sulphate, 5× SSC) were prepared in serial sectioned slides. Hybridizations were performed at 55°C overnight in a Dako hybridizer (Dako North America, Carpinteria, CA, USA). After hybridization, stringent washing was performed with 5× SSC (10 min) and 1× SSC (10 min) at 65°C and then 0.5× SSC (5 min) at room temperature. Slides were treated with blocking solution (0.1% Tween, 2% Sheep serum, 1% BSA in PBS) for 60 minutes at room temperature. Sheep anti-Dig-AP was diluted 1:500 in PBS with 0.05% Tween, 1% sheep serum and 1% BSA and was used to incubate slides for 60 min at room temperature. Slides were washed three times for 5 minutes each with TBST (0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.5% Tween-20) and then incubated with NBT-BCIP working solution with 0.2 mM levamisol (Sigma, St. Louis, MO, USA) in the dark overnight at room temperature. Slides were stained with Nuclear Fast Red™ (Vector Laboratories) for 1 min, and finally mounted with HistoMount solution (Life Technologies).
Patients' clincopathological parameters.
Slides were observed under an Olympus BX41 microscope (Olympus Corporation of the Americas, Center Valley, PA, USA) Multiple fields of view were examined for each slide. Each miR-21 ISH slide was scored by two independent reviewers for intensity of staining reaction and distribution in positive cells using the following scale: 0=negative; 1=weak ISH staining intensity and a positive cell distribution percentage less than 10%; 2=moderate ISH staining intensity and a positive cell distribution percentage of 11-50%; 3=strong ISH staining intensity and a positive cell distribution percentage greater than 50%. When there was a discrepancy, a consensus score was determined by the two reviewers. Images representing overall specimen status were recorded with DP controller software (Olympus, Shinjuku Monolith, Japan).
TaqMan® microRNA assay for breast cancer cells. Breast cancer cell lines MCF-7 and MDA-MB-231 were grown in Dulbecco's modified Eagle's medium (Life Technologies) supplemented with 10% fetal bovine serum (Life Technologies), 100 units/ml penicillin and 100 μg/ml of streptomycin (Life Technologies). Cells were cultured at 37°C in an incubator with 5% CO2. For the real-time polymerase chain reaction (PCR) assay, the miRNAs of MCF-7 and MDA-MB-231 cells were extracted using the mirVana™ miRNA Isolation kit (Life Technologies) by following the manufacturer's instruction. miR-21 and reference snRNA U6 TaqMan® MicroRNA Assays (Life Technologies) were performed. PCR was performed by 95°C for 20 seconds, followed by 40 cycles of 95°C for 1 sec then 60°C for 20 sec in a StepOnePlus™ real-time PCR System (Life Technologies). The relative expression of miR-21 normalized to that of snRNA U6 was calculated using StepOnePlus™ software (Life Technologies), and presented by values of -ΔCT.
Immunohistochemical analysis of smooth muscle myosin and p63. Immunohistochemical analysis was performed by using standard techniques. Briefly, after 4 μm paraffin-embedded sections were heated and de-paraffinized, Bond™ Epitope Retrieval Solutions (Leica Microsystems, Buffalo Grove, IL, USA) were used to expose binding sites for antibody. Endogenous peroxidase and biotin were blocked. Sections were incubated with smooth muscle myosin mouse monoclonal antibody (SMMS-1; Cell Marque, Rocklin, CA, USA) diluted 1:200, or with pre-diluted mouse antibody against p63 (Biocare Medical, Concord, CA, USA) according to the manufacturers' instructions. Slides were then washed and developed using Bond™ Polymer Refine Detection (Leica) and finally counterstained with hematoxylin, dehydrated and coverslipped. Slides were observed under an Olympus BX41 microscope (Olympus Corporation of the Americas)
Statistical analyses. The correlation between miR-21 ISH scores and clinical pathological parameters, including age, grade, tumor size, positive lymph node status, and molecular subtypes, was determined by the Fisher's exact test. p-Values less than 0.05 were considered significant.
Results
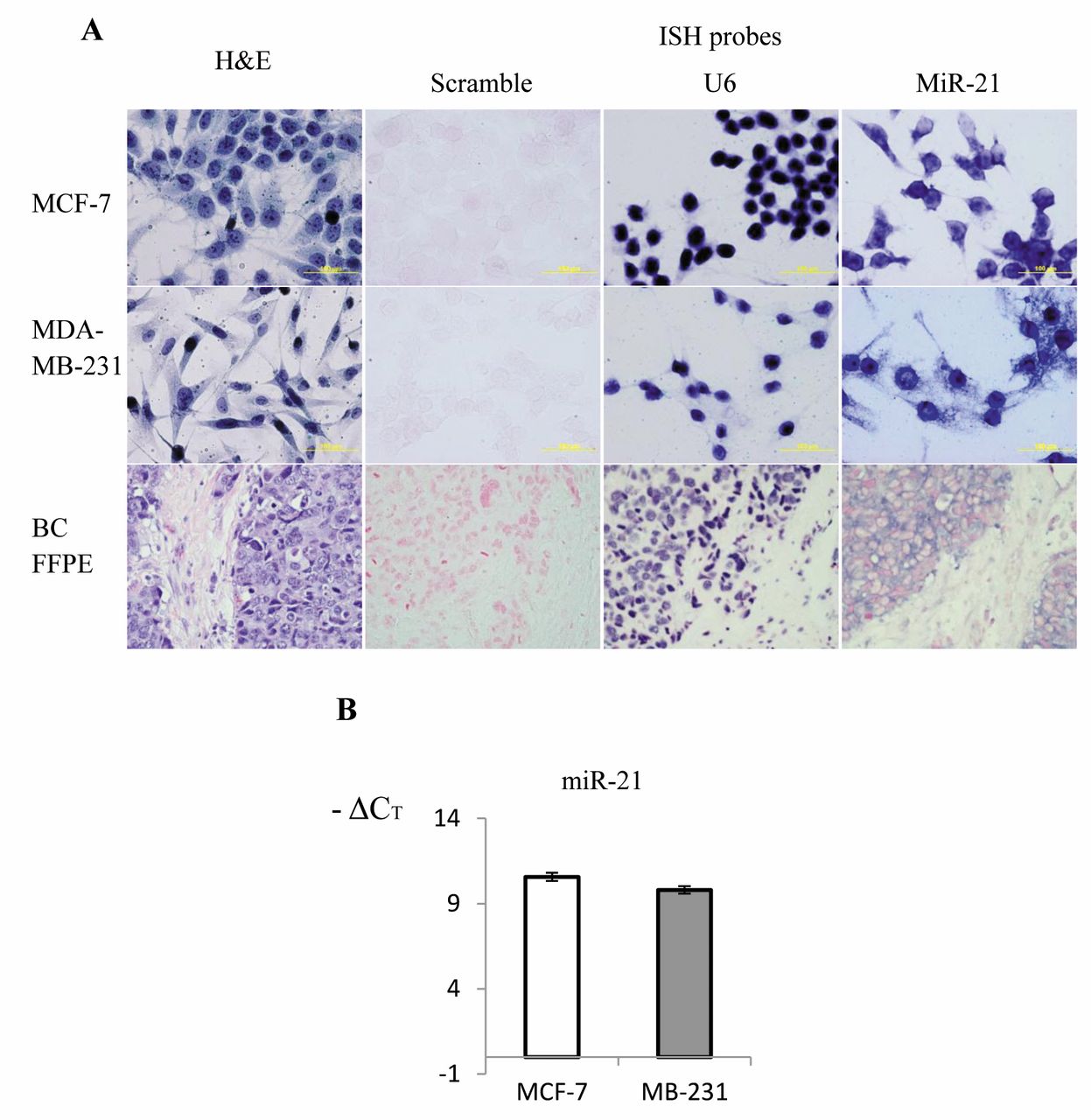
Detection of miR-21 in breast cancer cells and tissue samples by CISH. The miR-21 CISH assay successfully detected miR-21 in cultured breast cancer MCF-7, MDA-MB-231 cells and clinical breast cancer FFPE specimens. The negative control scramble probe did not produce any signals in any of the tests and the positive control U6 probe yielded very strong nuclear focused signals (Figure 1A). Both MCF-7 and MDA-MB-231 cells were positive for miR-21. The probe signals in both cells were located in cytoplasm, but were also seen in the nucleus because whole-cell samples were used in ISH. There was no obvious difference in miR-21 ISH staining intensity between MCF-7 and MDA-MB-231 cells. The malignant FFPE sample shown in Figure 1 was evaluated as histological grade III and clinical TNM stage T3pN1Mx. In the H'E-stained slide, malignant cells were highly invasive. ISH showed that miR-21 was mainly expressed in malignant epithelial cells. Dark blue miR-21 probe signals were detected in cytoplasm since the red nuclei stained by nuclear fast red were clearly visible.
The expression of miR-21 in MCF-7 and MDA-MB-231 cells was also analyzed by TaqMan real-time PCR. There was no obvious difference in miR-21 expression levels between MCF-7 and MDA-MB-231 cells (Figure 1B), which agrees with the miR-21 ISH results for these two cell lines. Both cell lines are epithelial cells derived from breast adenocarcinoma; MCF-7 is a luminal subtype and MDA-MB-231 belongs to a basal subtype (35). These data imply that miR-21 up-regulation exists in epithelia cells of both the cultured cell lines and the malignant breast tissue samples.
The intensity and distribution of miR-21 ISH-positive cells in breast cancer tissue. Positive ISH staining of miR-21 in the cytoplasm of epithelial cells was observed in most of the high-grade breast cancer cases. However, the color intensities varied among these cases. High color intensity miR-21-positive stromal cells were not observed. The miR-21 ISH-positive cancer-associated fibroblast-like cells presented medium or weak probe signals in some IDC cases. To test whether miR-21 is expressed in myoepithelial cells, we identified several residual ductal carcinoma in situ (DCIS)-like structures from IDC cases then performed IHC for SMM1 and p63 with serial sections of the same sample block. We found that the myoepithelial cells detected by markers SMM1 and p63 in IHC are not miR-21-positive cells and the epithelial tumor cells surrounded by this myoepithelial layer were much weaker or negative for miR-21 staining (Figure 2A). miR-21-positive cells were visible as epithelial cells in nearby typical invasive cancer regions. We also observed miR-21-positive spindle-like cells surrounding cancer islands in IDC cases (Figure 2B). These cells were not positive for either SMM1 or p63 IHC, and it was hard to determine if they were true stromal cells based on H'E staining. We also observed high intensity miR-21-positive cells on the edge of a tumor cell island in multiple IDC cases (Figure 2C).

Detection of miR-21 in breast cancer cells and tissue. A: miR-21 expression was detected in cancer MCF-7 and MDA-MB-231 cells and a clinical formalin-fixed paraffin-embedded (FFPE) sample. Cell morphology is shown using hematoxylin and eosin (H'E) staining. Negative control scramble probe, positive control snRNA U6 probes and miR-21 double DIG-labeled miRCURY LNA™ microRNA detection probes were used in chromogenic in situ hybridization (CISH) assays. NBT/BCIP was used to produce the colorimetric reaction and the positive signal is shown by a dark blue color. The slides were stained lightly with nuclear fast red. Nuclei are stained in red; magnification ×400. B: Real-time TaqMan polymerase chain reaction detection of miR-21 in MCF-7 and MDA-MB-231 cells.
In the present study, 50% of the cases were grade III disease; most cases exhibited miR-21-positive ISH staining in malignant epithelial cells. In approximately 25% of the cases, including grades I, II and III, a mixed pattern was found that contained miR-21-positive spindle-like cells wrapped around malignant epithelial cells or high intensity miR-21-positive cells surrounding the boundary of invasive tumor cell islands.
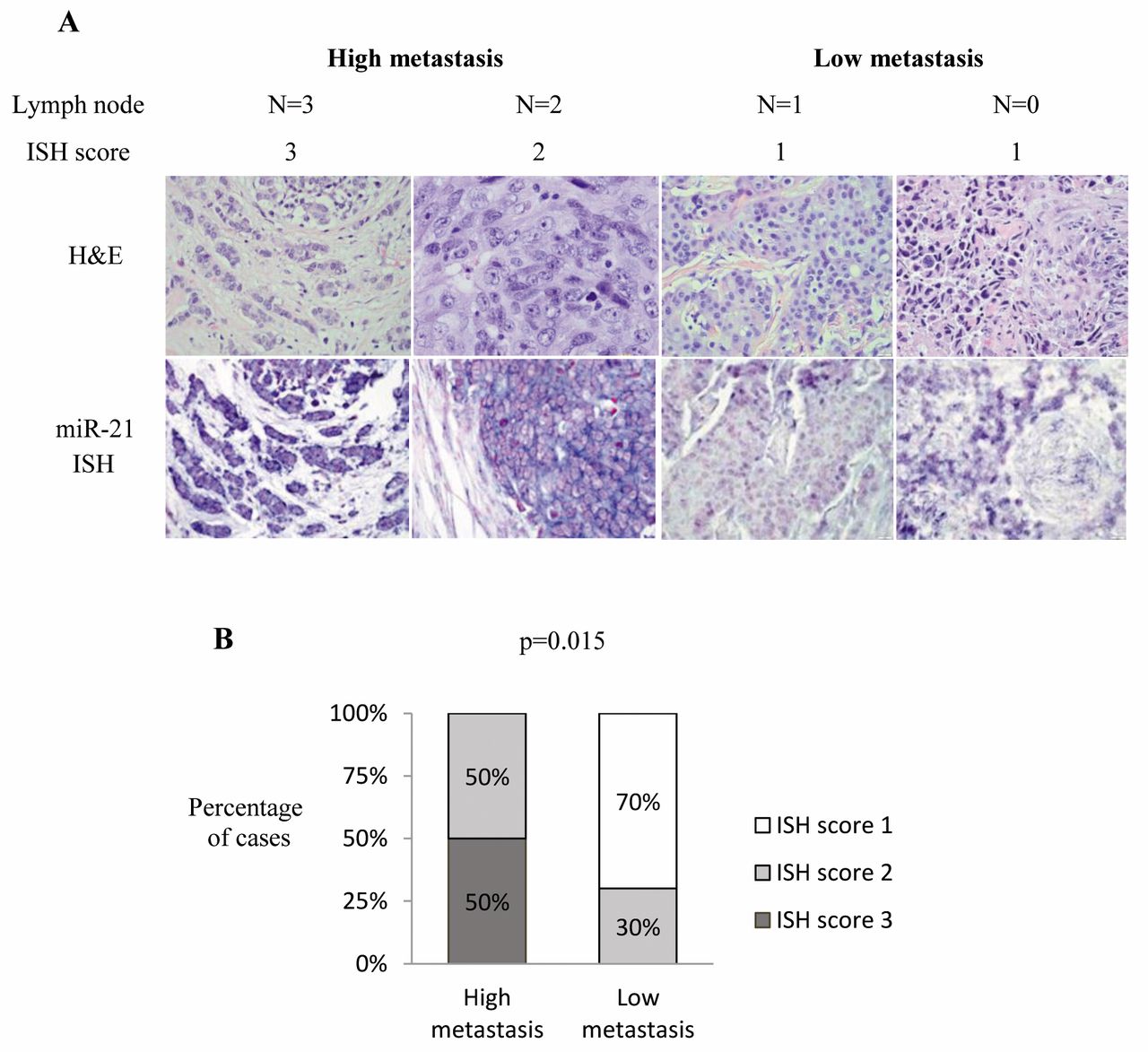
Correlation between miR-21 ISH score and clinicopathological parameters. The miR-21 ISH score correlated significantly with positive lymph node status. The high miR-21 scores (≥2) were highly correlated with advanced lymph node status (N≥2) in breast cancer (Figure 3), suggesting that miR-21 up-regulation is associated with cancer metastasis. No significant correlations were detected between miR-21 ISH scores and age, histological grade, tumor size, and any of the four molecular subtypes.
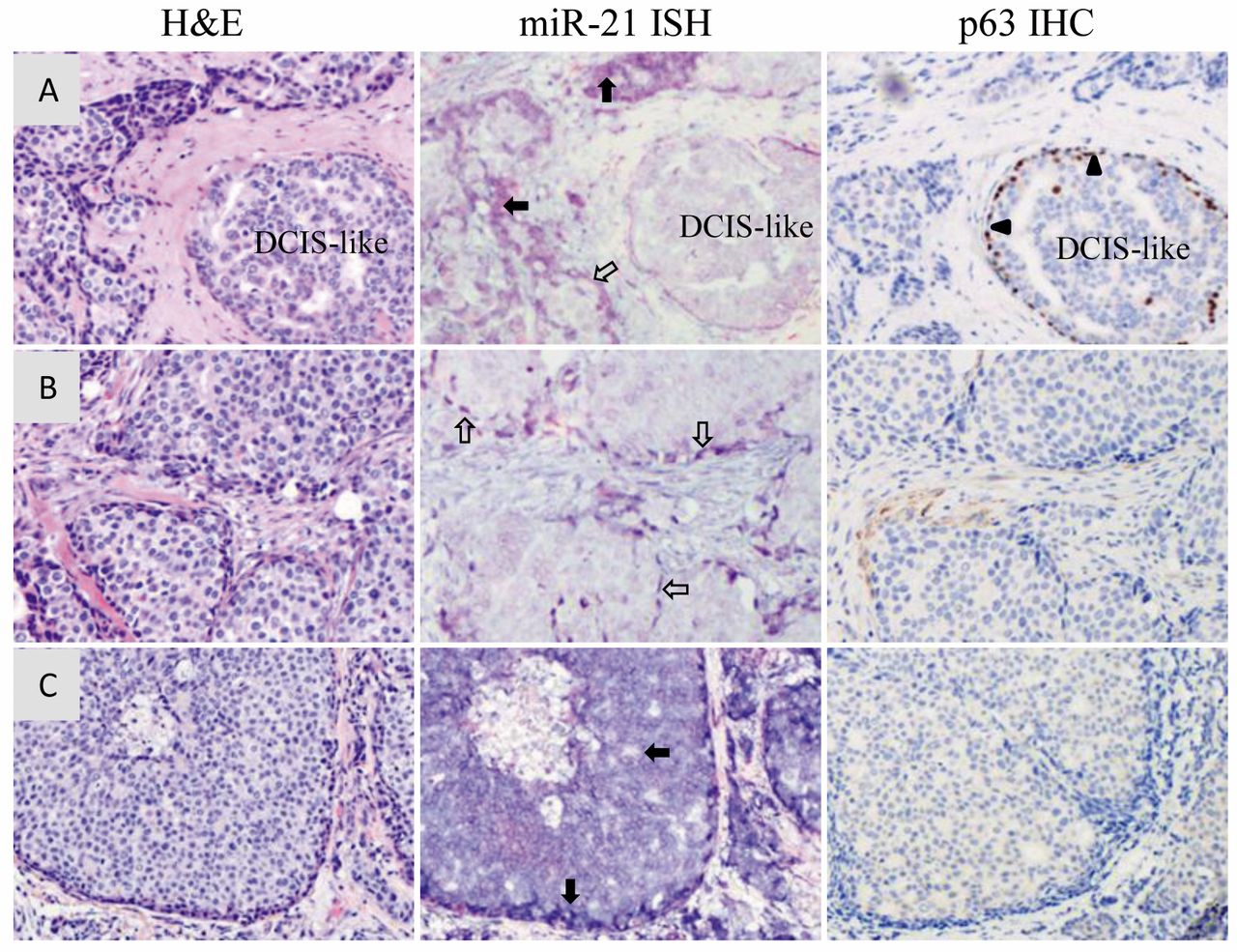
Distribution of miR-21-positive cells in breast cancer tissues by chromogenic in situ hybridization (CISH). A: Myoepithelial cells confirmed by p63 Immunohistochemistry (IHC) in residual residual ductal carcinoma in situ (DCIS)-like structure were not positive for miR-21 ISH staining. Epithelial cells surrounded by the myoepithelial layer were also miR-21-negative cells. Myoepithelial cells detected by p63 IHC stained dark brown, as indicated by triangles. miR-21-positive epithelial cells are indicated by closed arrows. B: Spindle-like miR-21positive cells as indicated by open arrows were found surrounding tumor cell islands. These spindle-like cells were negative for p63 by IHC. C: miR-21-positive cells with high intensity were detected at the edges of tumor cell islands, and these miR-21-positive cells were also p63-negative by IHC; magnification ×100.
Detection of miR-21 expressions in other histological types and benign breast tissues. We also investigated miR-21 expression and distribution in other types of breast tissues. The ILC case was diagnosed as breast cancer grade II and had no positive regional lymph nodes detected (Figure 4). The single-file pattern of malignant cells was clearly seen with H'E staining. A moderate level of miR-21 expression was detected in malignant epithelial cells by miR-21 CISH. Besides IDC and ILC cases, miR-21 distribution was also detected in other histological types of breast tissues including ADH, FD, FBT, SF and FA. miR-21 expression in ADH was very weak (Figure 4). In the benign samples, most cases showed a low level of miR-21 expression; however, a high level of miR-21 was detected in SF. The majority of altered miR-21 expression was found in ductal epithelial cells, but up-regulated miR-21 expression was also located in stromal cells in SF cases.
Discussion
Research in cell and animal model-based studies, as well as clinical investigations, has demonstrated that miR-21 plays an indispensable role in tumor proliferation, invasion, angiogenesis and metastasis (15, 16, 21, 22). However, the upstream regulation of miR-21 expression is not well-understood. In breast cancer, different locations of altered miR-21 expression have been reported, including tumor epithelial and stromal cells. These observations suggest that the expression of miR-21 can be induced by different factors, and activated miR-21 may play various roles in different cell types during the process of carcinogenesis.
In the present study, a high level of miR-21 was expressed in breast carcinoma cell lines MCF-7 and MDA-MB-231, as evidenced by TaqMan real-time PCR. The sub-cellular location of miR-21 was cytoplasmic as identified by miR-21 ISH. Yan et al. reported a similar observation (21). This finding supports the conclusion that the target cells of miR-21 expression can indeed be cancer epithelial cells. The tissue distribution of up-regulated miR-21 cells in cancer have been reported. In situ miR-21 staining was found in cancer epithelial cells in lung, pancreas and prostate (28, 36), while others found a high level of miR-21 in the stroma of colorectal cancer (37, 38). In breast cancer, several groups had reported similar observations that miR-21 was overexpressed in breast cancer epithelial cells (22, 36, 37). Contradictorily, Rask et al. showed that miR-21 was predominantly seen in cancer-associated fibroblast-like cells in grade I and grade II breast carcinomas (30) and Sempere et al. found a high level of miR-21 expression within tumor-associated fibroblasts (31). In a recent study based on tissue microarrays, MacKenzie et al. found that most breast carcinomas up-regulate miR-21 expression in the stroma, while a small sub-group with high miR-21 expression exclusively within epithelia were associated with poor clinical outcome (32). These conflicting findings may reflect either the complexity of genetic background of samples or differences in the sensitivity and specificity of ISH methods. There currently exists no standard miRNA ISH method for anatomic laboratory use. In our study, the majority of cases were grade II and grade III disease, with high level miR-21 expression mainly detected in invasive cancer epithelial cells. There were some in situ miR-21-positive stromal cells detected in various cases, but the probe signal intensities were weak compared to that in nearby positive epithelial cells.

The correlation between miR-21 in situ hybridization (ISH) score and lymph node status. A: Four cases with their lymph node status (N=0, 1, 2 and 3) and miR-21 ISH scores are shown as examples. Both hematoxylin and eosin (H'E) staining and miR-21 ISH images are presented for each case. B: miR-21 ISH score significantly correlated with lymph node status. Highly metastatic (N≥2) cases had higher miR-21 scores. p=0.015 by Fisher's exact test (total of 14 cases).

Detection of miR-21 expressions in different histological types of breast tissues by using chromogenic in situ hybridization (CISH). ILC, Infiltrating lobular carcinoma; ADH, atypical ductal hyperplasia; FBT, fibrofatty breast tissue; FD, fibrocystic disease; FA, fibroadenoma; SF, stromal fibrosis.
It is very interesting to recognize a mixed pattern of positive miR-21 cell distribution in about 25% of cases in this study, in which up-regulated miR-21 expression was found in both spindle-like cells surrounding local tumor cell islands and nearby invasive tumor epithelial cells (Figure 2). It is difficult to identify these spindle-like flat cells as stromal cells because malignant cells are so invasive in these areas, and cell morphology on H'E staining does not fully support their identification as stromal cells. We also found an increased miR-21 in situ staining at the boundary of tumor cell islands. Not all cells were myoepithelial in origin, based on the negative staining reaction for IHC biomarkers SMM1 and p63. Recent research has strongly suggested that miR-21 overexpression is involved in the early induction of epithelial–mesenchymal transition (EMT) (39, 40), which is a process highly associated with cancer aggressiveness and metastasis. The detection of these miR-21-positive spindle-like cells located in the boundary of cancer cell islands suggests a role of miR-21 in EMT and metastasis; however, further research is required to investigate the molecular evidence that supports this observation.
Normal myoepithelial cells act as natural tumor suppressors (41). As a result of malignant cell migration and invasion, myoepithelial cells are commonly absent from high-grade breast cancer. However, more recent evidence suggests that myoepithelial cells in a subset of pre-invasive residual ductal carcinoma in situ (DCIS) lose normal levels of tumor suppressors such as maspin and switch on tumor-promoter activity, creating a tumor microenvironment more permissive for invasion (42). Maspin has been identified as an miR-21 target (43). We found several residual ductal carcinoma in situ (DCIS)-like structures in invasive breast cancer; the myoepithelial cells identified by SMM1 and p63 markers were actually negative for miR-21 staining. This observation does not support the role of miR-21 in the transition of myoepithelial cells during cancer progression.
The expression of miR-21 is influenced by many factors in the context of malignancy. Bornachea et al. reported that miR-21 expression depends on mammalian target of rapamycin and Signal transducer and activator of transcription 3 activity and is required for metastasis in p53-deficient tumors (39). miR-21 expression is influenced by epidermal growth factor receptor activity (44). Echevarría-Vargas et al. reported that the binding of oncoprotein c-Jun to miR-21 DNA promoter regions leads to the aberrant increasing of miR-21expression through the c-Jun N-terminal kinases-1/c-Jun pathway (45). miR-21 can also be induced by transforming growth factor-beta (TGFβ) (46). miR-21 expression induced by TGFβ facilitated invasion and metastasis of colon carcinoma cells (47). The variations in the source cells of miR-21 expression observed by us and other groups may reflect the consequence of multiple miR-21 up-stream regulation pathways that function during different phases in cancer progression.
The findings of many groups have demonstrated that miR-21 plays an important role in EMT and cancer metastasis (21, 39, 40, 47). Based on their microarray result, Yan et al. found that miR-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and a poor prognosis (16). The primary finding in this study, that high miR-21 expression is correlated with advanced lymph node status, indicates that miR-21 is a valuable prognostic biomarker.
Low levels of miR-21 expression in normal breast epithelial cells has been reported by others (28, 29, 31). In this study, we demonstrate the miR-21 expression and distribution in ADH and four benign types of breast tissues including FD, FBT, SF and FA. miR-21 expression in most of the benign cases was undetectable or at a very low level, except for an increased level of miR-21 in SF. This observation implies that certain functions of miR-21 may be responsible for abnormal cellular proliferation in benign breast samples. It will be very interesting to investigate whether such irregular miR-21 expression in benign samples is temporary or persistent in nature.
Acknowledgements
The Authors would like to thank Dr. Indra Balachandran at Albany College of Pharmacy and Health Sciences, for the discussion and suggestion of this work. We thank Nicole Valade and Sara Laviska at Albany Memorial Hospital for technique support in clinical specimen preparation. They also thank Michael Racz, Albany College of Pharmacy and Health Sciences, for assistance in statistical analysis. This study was supported by Albany College of Pharmacy and Health Sciences Start Up Funds for Binshan Shi.
Footnotes
↵# Current address: Danbury Hospital, 24 Hospital Avenue, Danbury, CT 06810, U.S.A.
- Received March 7, 2015.
- Revision received March 18, 2015.
- Accepted March 20, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.