


Abstract
The electrical current exclusion (ECE) principle provides an alternative to common methods of cell diameter measurement and especially in atrophy and cancer associated cachexia research. C2C12 myoblasts were differentiated into myotubes and treated with 100 μM dexamethasone to induce atrophy in vitro. Subsequently, they were incubated for 24 h with media containing different concentrations of curcumin and/or branched-chain amino acids (BCAAs) in order to counteract atrophy. After treatment with curcumin, an increase in cell diameter was detectable; the highest increase with 13.9±0.4% was seen with 10 μM curcumin. The combination of curcumin and BCAAs showed an increase of 13.4±1.2 %. Cell diameter measurement via the ECE showed that curcumin, and curcumin in combination with BCAAs, were able to restore atrophic C2C12 myotubes. Therefore, the application of ECE in muscle atrophy and also cancer-associated cachexia research allows rapid screening of novel compounds in order to test their efficacy in vitro.
Screening for therapies and drugs that prevent muscle atrophy and cachexia have gained in importance. The multifactorial syndrome of cachexia is associated with decrease of body weight, emaciation and gradual changes in vital body functions, which leads to a distinctly reduced quality of life (1). Muscle atrophy and cachexia are side effects occurring especially in cancer patients (2). Cachexia is prevalent in 30 to 80% of cancer patients; however, its severity depends on variations in the tumour's phenotype and on the genotype of the cancer patient himself (3, 4). Fifteen percent of these patients suffer from an immense loss of more than 10% of their body weight (1). The decline in body weight results from chemotherapy, radiotherapy or surgery and is often accompanied by anorexia (5, 6). In vivo models showed that increased protein synthesis existed, although anorexia was absent (7). A study of cancer cachexia points out that weight loss is a prognostic factor with an important impact: with an increased progression of cachexia and a lower survival rate (8).
In addition to pain, fatigue and other physical and psychological implications, especially the catabolic loss of muscle mass itself has devastating consequences for the patients (1). While the amount of non-muscle proteins is relatively unchanged, the mass of muscle tissue itself decreases (9). This process, called atrophy, is described by sinking protein synthesis and increasing protein degradation; first of all, the ATP-dependent ubiquitin-proteasome pathway and the activity of lysosomes are involved (1, 10). The state of atrophy is connected to a high concentration of ubiquitin-protein conjugates and the transcription of components belonging to the ubiquitin degradation pathway is enhanced (10). For curing cachexia patients, the understanding of mechanisms leading to tissue wasting is a matter of importance. In order to discover appropriate drugs or agents against cachexia, many investigations target on several signalling pathways which are involved in the ubiquitin-proteasome system or contribute to protein synthesis (11, 12). Examples of these drugs are branched-chain amino acids (BCAAs), especially leucine. BCAAs are able to intensify protein synthesis due to initiating signal transduction pathways (13). Leucine activates the protein kinase activity of mTOR via interactions with other factors, for example Raptor (14). Furthermore, curcumin was tested as an anti-cachectic drug. The yellow pigment, ingredient of the rhizome from Curcuma longa, is well-known for its antioxidant, anti-inflammatory and wound healing effects (15). Oelkrug et al. have analysed the effects of curcumin in cells in vitro and pointed out that curcumin is thoroughly able to counteract muscle atrophy (16). The investigators used the method of measuring the electrical impedance of cells. Another method of investigation is provided by direct measurement of cell diameter. Until now, high-precision quantification of cell diameter changes was time-consuming and required the use of costly equipment. When working in vitro, the heterogeneity of cells plays an important role. In addition, changes in cell diameter are quite small and difficult to detect. Due to these facts, calculating results with low statistical error is extremely challenging.

Western blotting to detect changes in aquaporine-1 concentration: aquaporin-1 (30 g of total protein per lane) prepared from protein extracts of C2C12 cells cultivated in differentiation medium for 0 (A), 3 (B) or 7 (C) days.
In this article we present a new method to detect changes in the diameter of murine C2C12 myotubes. In contrast to primary muscle cells, the handling and cultivation of C2C12 cells is actually quite simple. The fact that C2C12 cells can provide an adequate in vitro model for muscle atrophy questions has already been demonstrated in many studies (17, 18). To enhance the growth of myotubes, cells received media supplemented with insulin-transferrin-selenium. Changes in cell diameter which occur during atrophy/cachexia were detected after treatment with dexamethasone. Experiments by Trevor et al. indicate that the response of C2C12 myotubes to dexamethasone reflects the response of skeletal muscle in vivo (10). The method for detection and quantification of the diameter changes is carried out with the use of the electrical current exclusion (ECE) principle. In most cell culture laboratories the CASY® Model TTC- Cell counter and analyser simplifies routine work. This system is based on the electrical current exclusion principle where a signal is generated when a cell or particle passes through the low voltage field in the CASY®'s measuring pore. Each particle is scanned with a frequency of one million measurements per second (19). Furthermore, the model uses pulse area analysis to maximize the gained information. It ensures that the properties of every single cell are included in the size distribution. Due to this technique, the CASY® Model TTC provides high-precise data of cell concentration, viability, volume, diameter, aggregation and debris. Important advantages are the short measuring intervals (10 seconds), the low volume sample (5-100 μl) and easy sample preparation.
In view of these facts, we analysed the effects of curcumin and BCAAs on dexamethasone-treated C2C12 myotubes with the innovative CASY® Model TTC.
Materials and Methods
Cell culture. C2C12 myoblasts (provided by Dr. Volker Adams, Clinic for Cardiology, Heart Center Leipzig, Leipzig, Germany) were cultured in growth media (RPMI-1640 (Gibco® by Life Technoogies, Darmstadt, Germany) supplemented with 10% fetal bovine serum (Sigma-Aldrich, Steinheim, Germany), 100 units/ml penicillin, 100 μg/ml streptomycin (Gibco® by Life Technologies) and incubated at 37°C with 5% CO2 until reaching 60% confluence. Subsequently, the C2C12 myoblasts were plated on 0.1% gelatine coated 12-well plates at a density of 5.6×104 cells/well and received growth media for 2 days until reaching 90-100% confluence. After aspirating growth media and washing with 1×Dulbecco's phosphate-buffered saline (DPBS), to achieve differentiation into C2C12 myotubes, the cells were fed with fusion media (RPMI-1640 including 2% horse serum (Sigma-Aldrich), 100 units/ml penicillin, 100 μg/ml streptomycin, 1% insulin-transferrin-selenium (Gibco® by Life Technologies). The medium was changed every 24 hours. The morphological changes and the present state of differentiation were checked microscopically. Additionally, the decreasing concentration of aquaporin 1 (AQP-1) was detected via Western blotting.
Three, 5 and 10 days after the initiation of the differentiation procedure, a fraction of C2C12 myotubes was directly measured with CASY® to receive the cells' mean diameter. Subsequently, the remaining myotubes were treated with 100 μM dexamethasone for 24 h to induce atrophy. A water-soluble dexamethasone solution (Sigma-Aldrich) was used which was easily dissolved in fusion media. To counteract atrophy, various concentrations of curcumin (Curcumin Solubilisate 6%, provided by AquaNova AG, Darmstadt, Germany), BCAAs (L-Leucin, L-Valine, Isoleucine; all from Sigma-Aldrich) and both supplements in combination were added for an incubation time of 24h (Table I). The supplements were dissolved in fusion media and stored at 4°C for a maximum of 3 days.
Diameter and volume measurement. For cell counting with the CASY® cell counter and analyser (Roche Applied Science, Mannheim, Germany) the establishment of a cell-specific setup for C2C12 myotubes was necessary. It involved the positioning of so called cursors that determined borders between cell debris and dead cells (normalization cursor) and between dead and viable cells (evaluation cursor). For this, samples of viable cells, dead cells and a mixture of dead and viable cells at a ratio of 30:70 were prepared. Cell suspensions were then diluted in a reaction tube 1:4 in freshly filtered CASY®ton, an electrolyte developed for cell counting. One hundred μl of these dilutions were mixed with 10 ml of additional CASY®ton in a CASY® cup to get the final dilution for the measurement of the viable sample. Another 100 μl of the 4-fold dilution was first mixed with 900 μl of CASY®blue, an alcoholic dilution, incubated for 2 min at room temperature and then mixed with an additional 9 ml of CASY®ton in a CASY® cup to get the final dilution for the measurement of the dead cell sample. Both samples were mixed by inversion three times prior to measurement, placed under the 150 μm measuring capillary and the measurement process was started. Results were displayed as size distribution. Leftovers of the viable sample and the dead cell sample were combined in a ratio of 70% viable and 30% dead cells, three times mixed by inversion and measured as well. This approach allowed to clearly determine the three different populations (debris, dead cells and viable cells) and set a normalization cursor and an evaluation cursor. All information was stored in a setup file with the name of the cell line and used for standardized cell counting.
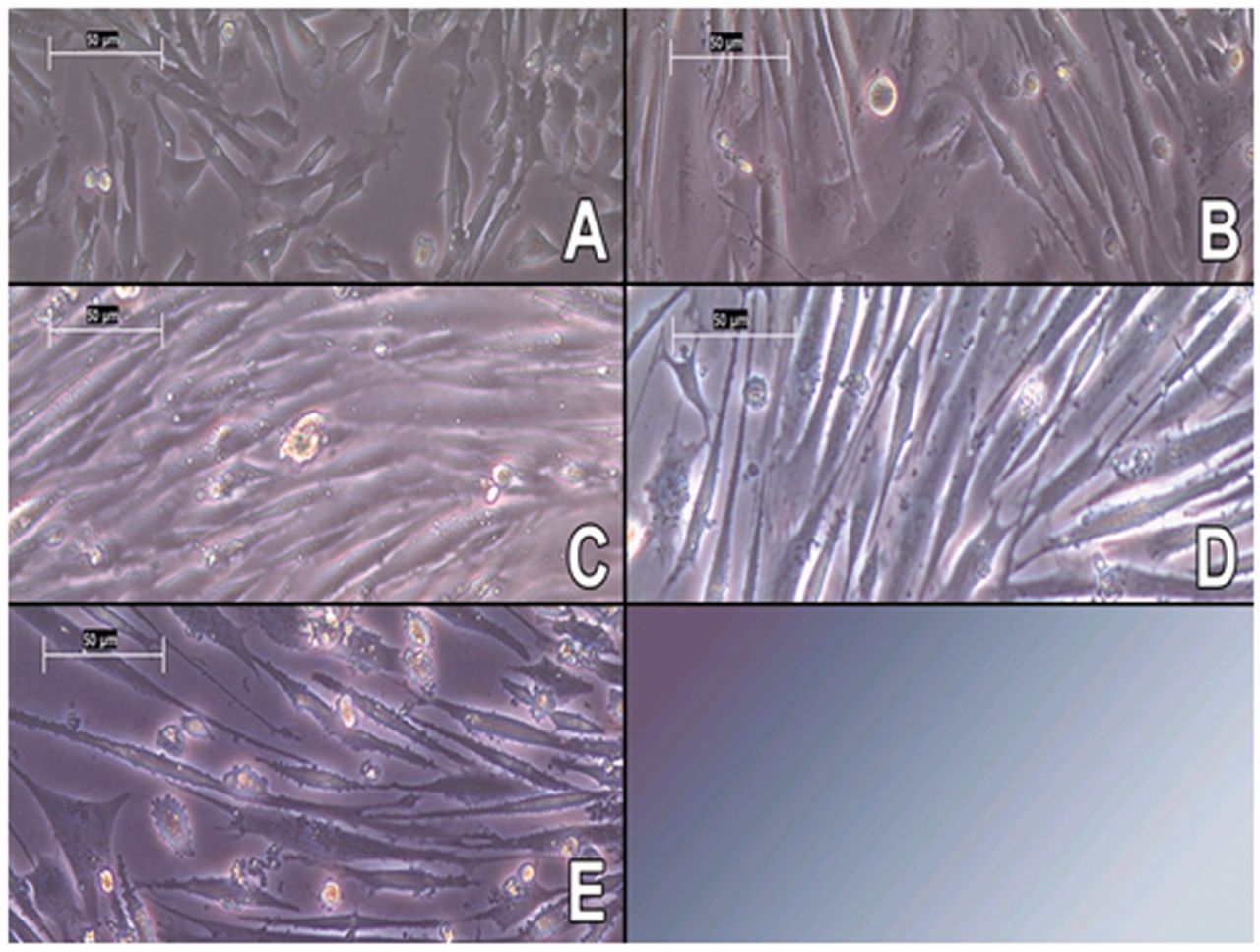
Microscope imaging of murine C2C12 myotube cultures: (A) myoblasts (B-E) treatment with fusion media for 3, 5, 7 and 10 days; 200× magnification.
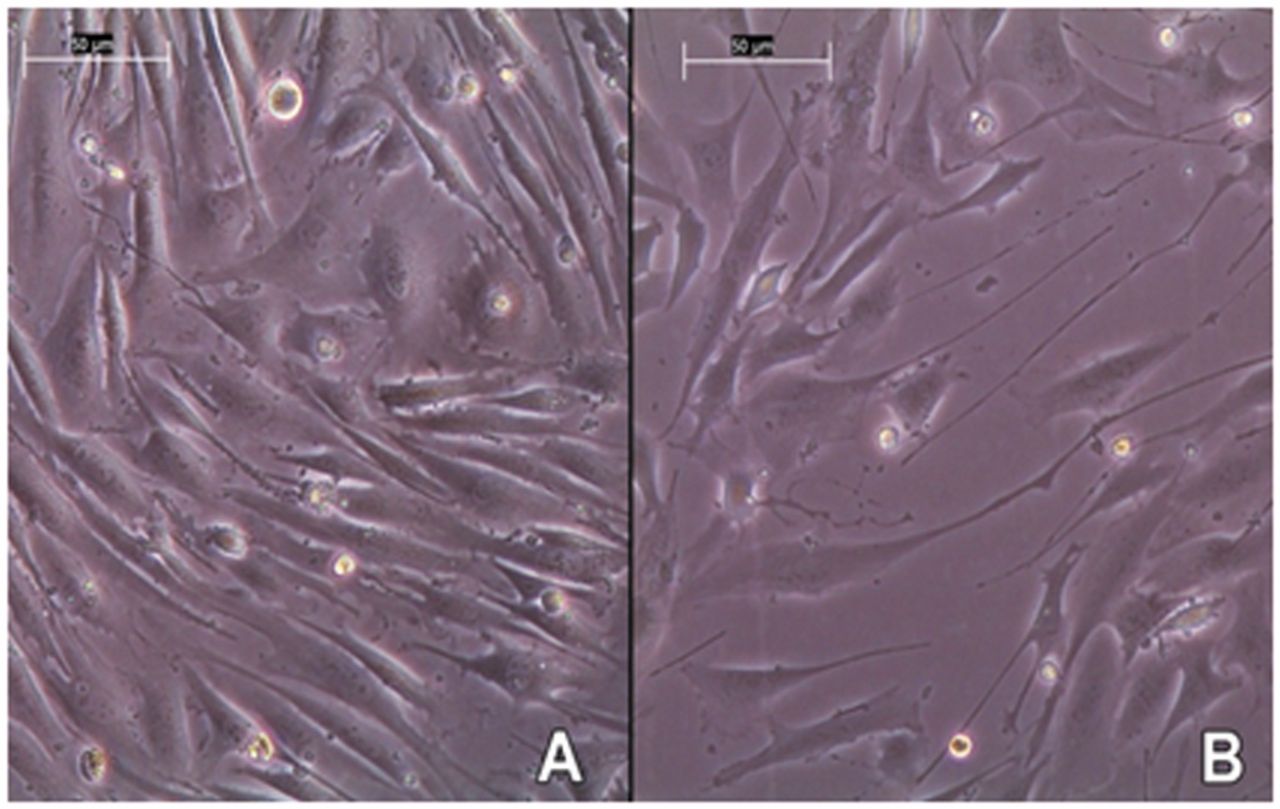
(A) image of differentiated C2C12 myotubes, (B) image of C2C12 myotubes treated with 100 μM dexamethasone for 24 h; 200× magnification.
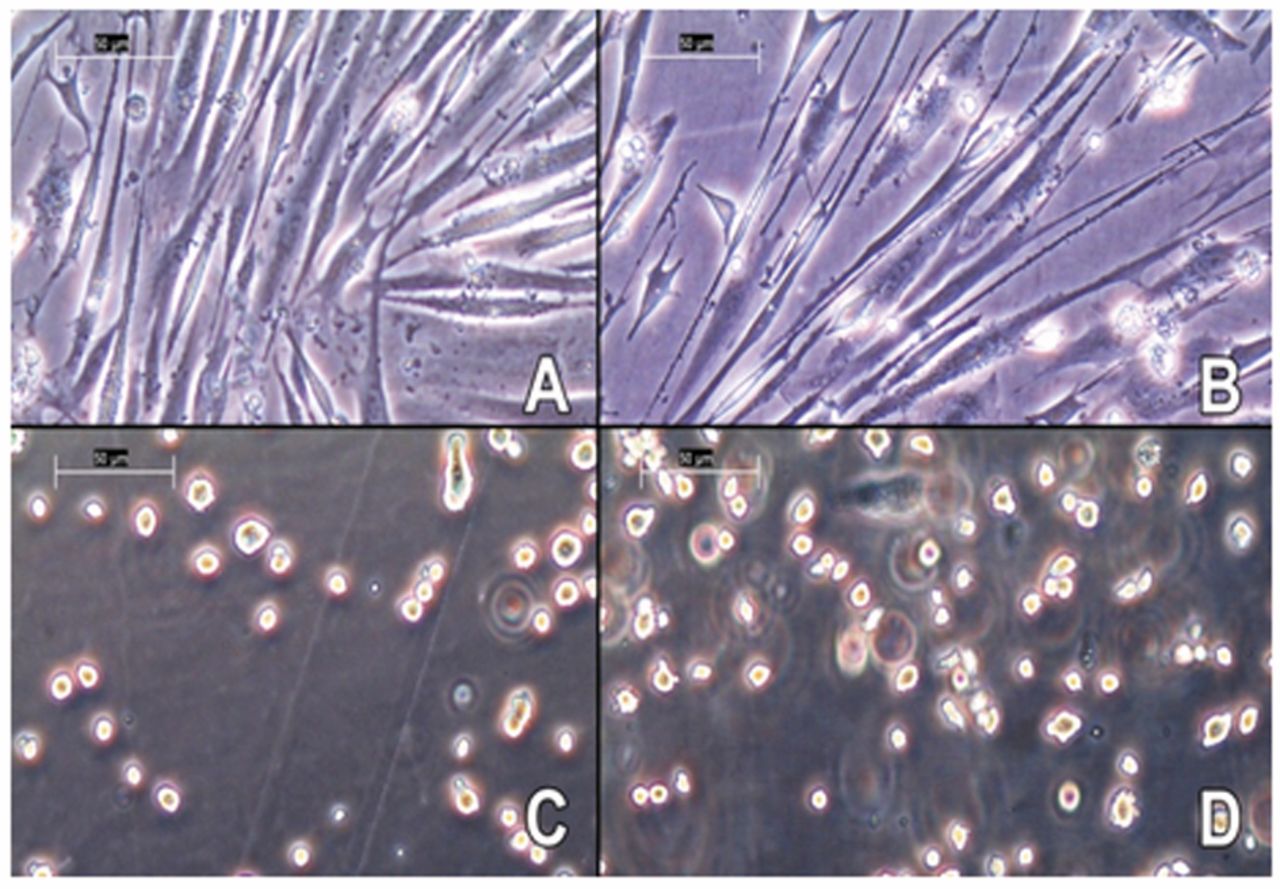
(A) untreated C2C12 mytobes, (B) cells treated with 100 μM dexamethasone, (C) untreated cells, incubated with TrypLE™ Express, (D) atrophic cells, incubated with TrypLE™ Express.
To avoid contaminating particles affecting the accuracy of cell count and sizing, prior background checks were performed. If the background value was below 100 counts/ml, CASY® was ready for cell analysis. Before analysing cells, the wells were washed with warmed DPBS, trypsinized and re-suspended in warm fusion media. Subsequently, the samples were prepared according to the following procedure: an aliquot of cell suspension (20 or 50 μl) was mixed with 10 ml sterile-filtered CASY®ton in a CASY®cup. After mixing by inversion, the prepared CASY®cup was placed under the measuring capillary. The CASY® device regularly measured three aliquots of 400 μl and the software calculated the mean diameter.
Detection of AQP-1 via Western blot. For detection of decreasing AQP-1 concentration, cells with different differentiation timing were analyzed via Western blotting. After 0, 3 and 7 days of cultivation with fusion media, a fraction of murine C2C12 cells was treated with Triton™ X lysis buffer (final concentrations: 150 mM NaCl, 0.1% Triton™X, 50 mM TRIS HCl pH 8.0, supplemented with protease inhibitor (Sigma-Aldrich). The cells were gently detached with a cell scraper and the mixture was incubated for 30 min at 4°C. Subsequently, cells were centrifuged at 12,000 rpm for 20 min at 4°C. The supernatant was transferred into a fresh 1.5 ml tube and the sample was stored at −80°C for later applications.
Prior to performing sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) with a 14% polyacrylamide-gel, the total amount of protein within the samples was determined using the Pierce™ BCA protein assay kit (Thermo Fisher, Schwerte, Germany). A 96-multiwell plate was filled with triplets of a 10 μl sample and a 190 μl reaction mixture. The standard dilutions were handled in the same manner. Post incubation, 30 min at 37°C, absorbance at 560 nm was measured via a microplate reader (Tecan Sunrise™, Männedorf, Switzerland). Calculation of the total protein amount was performed using a calibration graph out of the standard samples data. To prepare for electrophoresis, 30 μg of protein was mixed with loading buffer (TBS, glycerol, SDS, mercaptoethanol and bromo phenol blue; dilution of 1:4) and the mixture was heated to 95°C. After separation via SDS-PAGE, proteins were transferred to a polyvinylidene difluoride (PVDF) membrane via semidry-blotting. Subsequently, non-specific binding sites were blocked. The PVDF membrane was incubated with AQP1 (B-11): sc-25287 (Santa Cruz, Texas, USA) at a dilution of 1:200 overnight at 4°C, followed by incubation with goat anti-mouse, horseradish peroxidise-labelled secondary antibody (Abcam, Cambridge, UK) at a dilution of 1:20,000 for 1 h at room temperature. For reducing background noise, 1 h blocking with 5% milk solution and long periods of washing with DPBS containing 0.05% Tween 20 were necessary. Exposure of x-ray film was accomplished via SuperSignal® WestPico Chemiluminescent Solution (Thermo Fisher, Schwerte, Germany).
Analysis of curcumin's cytotoxicity on C2C12 myotubes (WST-1 assay). Analysis of cytotoxicity and the calculation of half maximal inhibitory concentration IC50 of curcumin were tested via the WST-1 assay (Cell Proliferation Reagent WST-1; Roche, Mannheim, Germany). Murine C2C12 myoblasts were seeded in 96-multiwell plates at a density of 5×103 cells/well. The cells received 150 μl growth media and incubated at 37°C with 5% CO2 for 24 h to reach confluence. To induce differentiation into C2C12 myotubes, the medium was replaced by fusion media and the cells were cultivated for an additional 24 h. Subsequently, the cells were incubated again for 24 h but this time were fed with fresh fusion media containing 5 μM, 10 μM, 15 μM and 20 μM curcumin or remained untreated. The samples and the blanks (media without cells) were prepared in triplets for calculation of the arithmetic mean. 15 μl WST-1 per well were added and the plate was incubated for 4 h at 37°C, with 5% CO2. The absorbance values at 450 nm were generated using a microplate reader (Tecan Sunrise™, Männedorf, Switzerland). Viability of cells was expressed relative to untreated cells. To determine cytotoxicity, 100 % was subtracted from these viability values. Cytotoxicity [%] was plotted against curcumin concentration [μM]. For calculation of curcumin's IC50 value, non-statistical regression analysis and the SigmaPlot software (Systat Software, San Jose, CA, USA) were used.

Response of atrophic C2C12 myotubes to curcumin: (A, B) after 3 days of differentiation, (C, D) after 4 days of differentiation (***p<0.001).

Cytotoxicity of curcumin towards differentiated C2C12 cells via the WST-1 assay.
Statistical analysis. All data are presented as mean±standard deviation. Statistical analysis and graphic presentations were made using SigmaPlot 10.0/SigmaStat 3.5 software (Systat Software, San Jose, CA, USA). p-Values were calculated using student t-test (***p<0.001).
Results and Discussion
Detection of AQP-1. Grundry et al. have reported that AQP-1 concentration decreases as differentiation of C2C12 progresses to myotubes (20). This decreasing concentration was detected via Western blotting showing the glycosylated form of the protein having a molecular weight of 35-45 kDa. Our results confirm that the intensity and size of AQP-1 decreases over differentiation time (Figure 1).
Initiation of atrophy and treatment with curcumin or branched-chain amino acids. The analysis with the electrical current exclusion principle provides direct data on cell diameter and cell volume. There is no need of converting values, which is the case when using impedance measurement or other methods. One sample included an average of 2,459±1,099 cells. The diameter of every single cell was detected and the CASY® software calculated the arithmetic mean out of this high-precise data, whereat the data of debris and dead cells was already excluded. Due to the high number of analysed cells in one sample, statistical errors are small.

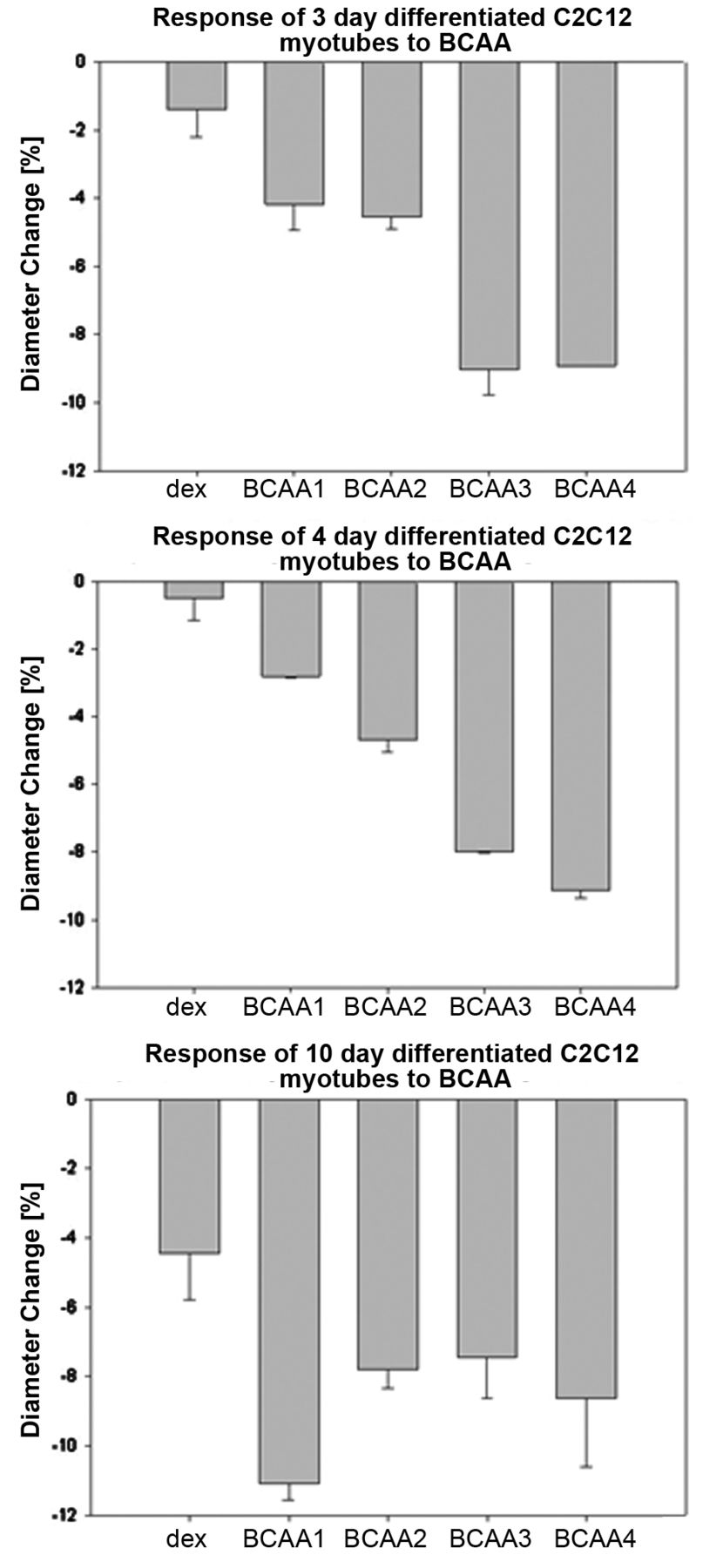
Diameter changes [%] of C2C12 myotubes in response to BCAA treatment.
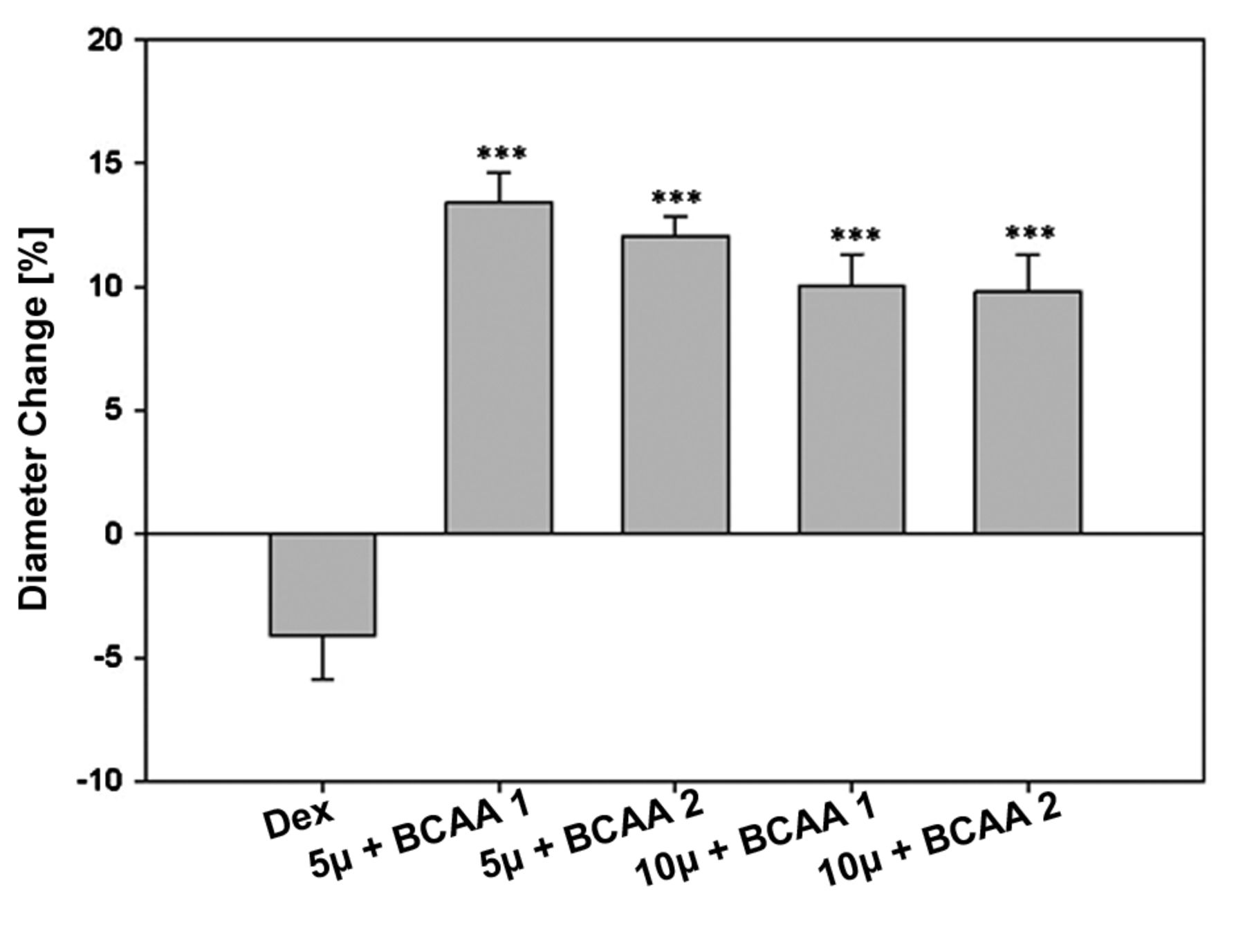
Diameter changes [%] in response to curcumin supplemented with BCAA (***P<0.001).
In addition to the diameter measurement with CASY®, microscope pictures were taken to check the viability and conditions of the cultured cells. As expected, the cell diameter and volume increased as a function of culture length as seen in Figure 2A-E. In Figure 2 A, the typical phenotype of mononucleated myoblasts is visible. During differentiation (Figure 2B-E), the myoblasts fuse and develop onto polynucleated myotubes. The microscope images coincide with data created via the CASY® measurement. The average diameter on the 3rd day of differentiation was (16.27±0.6) μm and on day 10 the average diameter was (17.56±0.4) μm.
Treatment with 100 μM dexamethasone for 24 h caused a decrease in cell diameter, which was detectable via the CASY measurement. The highest decrease from average (17.56 μm to 16.78 μm; 4.4%) is seen on C2C12 myotubes with a differentiation duration of 4 days. This data coincides with microscope imaging. In contrast to untreated cells (Figure 3A), the appendices of atrophic myotubes (Figure 3B) are obviously thinner and show a distinct atrophic phenotype. Although this change in diameter is quite difficult to detect via a microscope if the cells are trypsinized (Figure 4), the different diameters of untreated and atrophic cells are clearly measurable with the CASY® System. Other studies claim that treatment with dexamethasone induces a decline in cell diameter up to 50% or a change in the cell index by 20% (21, 10). Such major changes were not reached in this experiment; however, our analysis demonstrates the high sensitivity of cell measurement with the electrical current exclusion principle that can detect even a small change in cell diameter due to a moderate state of atrophy.
The compositions of branched-chain amino acids (BCAAs) and curcumin. Final concentrations in RPMI-1640 medium are shown.
Several studies verify the positive effects of curcumin on health issues due to the acceleration of wound healing and anti-inflammatory properties (22). Thus, our questions were if curcumin could help to overcome atrophy and if a significant increase in the cell diameter is detectable via the CASY® system. Therefore, the atrophic C2C12 myotubes were cultured in media with various curcumin concentrations for 24 h. Changes in myotubes' diameter are shown in Figure 5. An increase in the cells' diameter was reached with 5 μM, 10 μM and 15 μM curcumin regardless of the differentiation time. The largest increase from 16.62±0.7 μm to 18.93±0.7 μm (nearly 14%) was reached with 10 μM curcumin on 7 day differentiated cells. On the contrary, treatment with 20 μM curcumin provokes cell death and reduced viability to 43%. The WST-1-assay was used to calculate an IC50 of 20.6 μM curcumin for differentiated C2C12 (Figure 6), which was shown to correlate with the microscope images obtained and relevant literature (16). These results show that treatment with curcumin is a possibility to counteract atrophy, but there is just a small therapeutic window. Another challenging issue is the poor bioavailability of curcumin. In our experiments, we used NovaSOL® Curcumin (AQUANOVA AG, Darmstadt, Germany) which, according to the manufacturer, exhibits high bioavailability, full water solubilisation and special micelle structure for superior absorption (23). Seemingly, these qualities contribute to the immense increase in diameter of atrophic cells. However, attention must be paid on the instability of curcumin in an aqueous solution as it is rapidly degraded to trans-6-(4’-hydroxy-3’-methoxyphenyl)-2,4-dioxo-5-hexenal, ferulic acid or feruloyl methane (24). For use as a nutritional health supplement, the extract of Curcuma longa is often enclosed in capsules, combined with piperin, the extract of black pepper. Piperin has been shown to increase the bioavailability of curcumin (25).
We further investigated whether the addition of BCAAs: (L-leucine, L-valine and Isoleucine) could prevent atrophy. These BCAAs make up 35% of the essential amino acids in mammalian muscle proteins (26). In contrast to other essential amino acids which are oxidized in the liver, BCAAs are oxidized locally in skeletal muscle cells (27). Furthermore, Kimball et al. found out that BCAAs play an important role in promoting muscle protein synthesis in vivo (28). Treatment of mice bearing the MAC16 tumour with BCAAs caused an increase in muscle mass due to enhanced protein synthesis and moderated loss of body weight (29).
The results on the diameter change due to BCAA treatment are shown in Figure 7. The data indicate that no concentration of BCAA was able to block or reverse atrophy in cells. The diameter was lower than in the non-atrophic control and no clear trend between BCAA concentration and diameter change was visible. These results are contrary to several issues of BCAA studies, which indicate anti-atrophic effects of BCAA (1, 13, 27). In contrast to our protocol, BCAAs were administered over a longer period of time. Nevertheless, the overall opinion is that BCAAs, especially leucine, are useful nutritional supplements for sport and exercise or for treatment of muscle atrophy (1, 13, 27). However, Shimomura et al. presented an article pointing out that the most effective ratio for BCAA is not yet known (13). Here, the use of the electrical current exclusion principle (ECE) for direct measurement of cell diameter and cell properties provides a good solution. Screening different drug concentrations, as well as different processing times, could be tested easily.
The third part of our project was to test the effects of curcumin combined with BCAA on atrophic C2C12 myotubes. Myotubes were incubated with a mixture out of various concentrations of curcumin and BCAA. The highest increase in cell diameter was seen when cells were treated with 5 μm curcumin and BCAA1 after 24 h. On the whole, atrophy could be overcome at each combination of curcumin and BCAA which means that the cell diameter, prior to dexamethasone treatment, was reached (Figure 8). The positive effects of curcumin and BCAAs have been described previously (13) and it is possible that longer treatment with curcumin and BCAA may enhance the growth of atrophic cells.
In conclusion, the results clearly show that the use of the electrical current exclusion principle for direct measurement of cell diameter performed by the CASY® model TTC is beneficial. Due to the quick analysis of thousands of cells, an immense amount of high-precise data is created. It also enables to calculate results with low statistical error. By comparison to other methods that measure cellular diameter, such as the electric impedance measurement, electron tomography or optical coherence tomography, no high priced equipment is needed. The CASY® Model TTC enables every researcher in every laboratory to gain information about viability and diameter of cells in a high- throughput format.
Acknowledgements
The work presented here was made possible by funding from the Federal Ministry of Economics and Energy (03EGSSN113, 01.03.12-28.02.13).
Footnotes
-
↵* These authors contributed equally to this work.
- Received December 7, 2014.
- Revision received January 21, 2015.
- Accepted January 23, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.