


Abstract
Background: The objective of the present study was to investigate how oxidative status influences the effective ness of cytotoxicity of artemisinin towards cancer cells. It is hypothesized that antioxidants would reduce, whereas pro-oxidants would enhance, cytotoxicity. Materials and Methods: Molt-4 human leukemia cells were incubated with vitamins C, E, D3, dexamethasone, or hydrogen peroxide alone or in combination with dihydroartemisinin (DHA). Concentrations of these compounds studied were similar to those achievable by oral administration. Viable cell counts were performed before (0 h) and at, 24 and 48 h after treatment. Results: Vitamin C, vitamin D3, dexamethasone, and H2O2 caused significant Molt-4 cell death. Vitamin E caused an increase in Molt-4 cell growth. Vitamin C and vitamin D3 significantly interacted with DHA at the 48-h time point and with H2O2 at both 24-h and 48-h time points. Conclusion: Cellular oxidative status could alter the potency of artemisinin in killing cancer cells.
Environmental factors, such as nutrition and nutritional supplementation, can explain some of the variability seen among treatment outcomes of cancer patients (1). To explore this possibility, we tested the effects of several nutritional supplements commonly taken by cancer patients on an experimental cancer drug artemisinin, which we have been investigating as an alternative to current chemotherapeutics (2). Artemisinin, a compound isolated from the plant Artemisia annua L (sweet wormwood), and its derivatives are currently the most effective anti-malarials (3). Artemisinin compounds have also been shown to be potent and selective anticancer agents both in vitro and in vivo (2). Artemisinin's anticancer mechanism has been hypothesized to arise from the interaction of its endoperoxide moiety with free ferrous iron, which is more abundant in cancer cells, generating cytotoxic reactive oxygen species (ROS) (4, 5). There are several case reports of cancer treatment using artemisinin (6-8).
In the present study, we investigated whether various nutritional supplements interact with the more hydrophilic artemisinin analog dihydroartemisinin (DHA) in killing cancer cells. Since artemisinin works by overloading cancer cells with ROS, we selected molecules that are known to affect the oxidative status of cells. For their antioxidant properties, we studied sodium ascorbate (vitamin C) and Trolox (vitamin E). For their pro-oxidant properties, in certain cancer cells, we studied vitamin D3, dexamethasone (dexamethasone 21-acetate), and hydrogen peroxide (H2O2). To evaluate the interactions of artemisinin with these agents, we used the commonly studied, rapidly growing and easily cultured Molt-4 human lymphoblastic leukemia cells.
Materials and Methods
Chemicals. All chemicals were purchased from Sigma Aldrich (St. Louis, MO, USA) unless mentioned otherwise.
Molt-4 cell culture. Molt-4 cells were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). They were cultured in RPMI-1640 media (Life Technologies, Grand Island, NY, USA) with 10% fetal bovine serum (ATCC) at 37°C with 5% CO2 and 100% humidity to reach a density of 6×105 cells/ml before diluting to a density of 6×104 cells/ml approximately 24 h prior to a treatment. Twenty-four hours of pre-incubation allowed the cells and media to be conditioned prior to exposure to supplements and experiments to begin at a density of approximately 1×105 cells/ml. At this time, cells were in the log phase. Viable cells were counted before drug treatment (0 h) and at 24 h and 48 h after treatment using a hemocytometer. To assess cell viability, we used trypan blue exclusion and identified dead cells based upon morphological criteria: chromatin condensation, formation of apoptotic bodies, shrinkage of the cytoplasm, and blebbing of the plasma membrane with irregular outline (9, 10). Only healthy cells were counted.
Results of two-way ANOVA showing significant differences of dihydroartemisinin (DHA), supplements (ascorbate, Vitamin D3, dexamethasone, Trolox and hydrogen peroxide) and interaction (DHA+supplements) effects at the 24-h and 48-h time points. Results showing significance are in bold.
Cell treatment. There were four treatment groups: control (no-drug treatment), Dihydroartemisinin (DHA) alone, supplement alone, and DHA+supplement. DHA (Holley Pharmaceuticals, Chongqing, China) was used at a concentration of 12.4 μM. This concentration was used because it is similar to the maximum blood concentration in rats after an oral dosage of 10 mg/kg (11). It was also the concentration used in our previous studies. The supplement samples were added with the following concentrations: ascorbate 256 μM, vitamin D3 75 nM, Trolox 45.8 μM, dexamethasone 21-acetate 10 μM, and H2O2 8.82 μM, according to standard achievable in vivo concentrations of these compounds. Dimethyl sulfoxide (DMSO) was the solvent for DHA, dexamethasone, Trolox, and vitamin D3. All other compounds were dissolved in RPMI-1640 media. All groups were controlled at 1% DMSO by adding the appropriate amount of DMSO directly to the suspension culture at the experimental initiation. Each experiment was performed at least three times.
Data analysis. Data were expressed as the ratio of cell counts at the 24-h or 48-h time point relative to time zero. The GraphPad Prism 6.03 software (La Jolla, CA, USA) was used for statistical analysis. A two-way ANOVA was performed to test for significances of main effects of DHA and supplements, as well as their interaction effect. Pair-wise comparisons were analyzed using the Newman Keul's test. A p<0.05 was considered statistically significant.
Results
Data of effects of DHA and different supplements on Molt-4 cells are presented in Figure 1. Results of two-way ANOVA showing significances of main and interaction effects are presented in Table I.
In all cases, DHA-alone significantly decreased cell counts relative to untreated cells at both 24-h and 48-h time points. All supplements studied also affected cell counts. Compared to control, ascorbate and H2O2 both caused a significant decrease in cell count both at 24-h (p<0.05 and p<0.0001 for ascorbate and H2O2, respectively) and 48-h (p<0.0001 for both ascorbate and H2O2) time points. Dexamethasone and vitamin D3 caused a significant decrease in cell count only at the 48-h time point (p<0.01 for both supplements). A significant increase in cell count was observed at the 24-h time point for Trolox (p<0.05). Thus, all supplements, except for Trolox, caused a decrease in cell counts.
Hydrogen peroxide in combination with DHA caused significantly greater cell death than DHA-alone at both time points (p<0.01 and p<0.05 for 24-h and 48-h time points, respectively). Also, the effect of dexamethasone combined with DHA was found to be significantly greater than DHA alone at the 48-h time point (p<0.05). However, cell counts of ascorbate, vitamin D3 or Trolox in combination with DHA are not significantly different from DHA alone at both 24-h and 48-h time points. This is reflected in the results of the two-way ANOVA (Table I) indicating that H2O2 significantly interacted with DHA at both time points, whereas ascorbate and Vitamin D3 were found to interact at the 48-h time point. No significant interaction effect was observed between DHA and Trolox or dexamethasone.
Discussion
The effectiveness of artemisinin may be due to its free radical generation capabilities in the presence of Fe2+ causing widespread effects on cancer cells (12, 13). If patients are undergoing cancer therapies with drugs like artemisinin, which rely on oxidative stress, then it is important to know how supplementation with oxidative-state modulating compounds will affect the therapy. It may be possible to improve therapeutic outcomes by compounding the oxidizing effect of artemisinin and certain supplements, pushing cancer cells toward oxidative overload (14).
Vitamin C, a common supplement, has been shown to act as both a ROS generator in the presence of iron and copper (15) and as an antioxidant (16). The normal blood concentration of vitamin C is approximately 64 μM in an adult man (17). We hypothesize that ascorbate acts more like a pro-oxidant in cancer cells due to their higher cytoplasmic iron levels. From our results, we can postulate that ROS generation is causing cell death independently and in combination with DHA. Variation in cancer therapy out comes with drugs targeting oxidative stress may be caused, in part, by vitamin C levels, since it is a commonly used supplement.
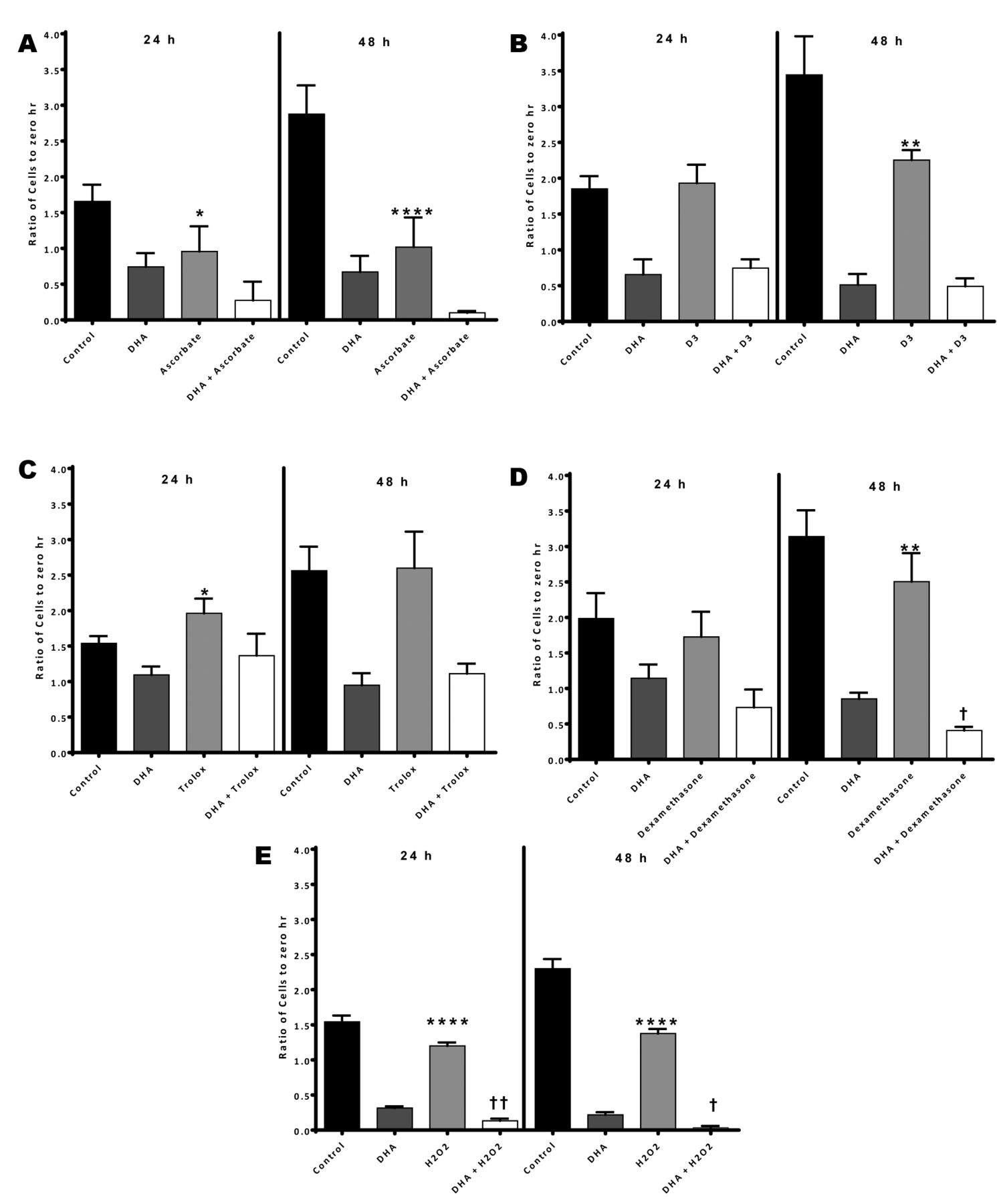
Effects of dihydroartemisinin (DHA), supplements, and DHA+supplements on Molt-4 cells at 24-h and 48-h time points. Error bar denotes standard deviation. *: p<0.05, **: p<0.01, ****: p<0.0001 comparing supplement with control. †: p<0.05, ††: p<0.01 comparing DHA+ supplement with DHA.
However, one must be careful in taking vitamin C in combination with DHA. Ascorbate can convert ferric iron into ferrous iron (18), the active form that reacts with artemisinin, generating short lived free radicals. If this happens in the stomach of a person who is consuming artemisinin along with ascorbate, ascorbate will convert ferric iron in foods to the ferrous form, which may react with artemisinin locally, making the therapy less effective. To avoid this issue, ascorbate may be administered intravenously or taken orally at a different time of day as artemisinin.
Vitamin D3 has been shown to act as a pro-oxidant and an antioxidant in malignant and non-malignant human prostate epithelial cells, respectively (19).Vitamin D3 concentrations range between 75 and 190 nM in the blood (20), varying with an individual's age, sunlight exposure, race and other supplement intake (21). Our results indicate that vitamin D3 reduces leukemia cell growth independently but has no significant interaction effect with DHA. The reason of the lack of an interaction effect is unknown. Perhaps, vitamin D3 becomes an antioxidant in cancer cells at longer time points. It may be of interest to study the interaction of vitamin D3 levels with artemisinin in vivo, as vitamin D3 is involved in regulation of calcium and phosphorus intake (22), immune function (23) and, thus, may be involved in more complex interactions with DHA in cancer cells.
The normal blood concentration of vitamin E is 22.9 μM, with a range of 12 to 59 μM (25). Our results with the water-soluble vitamin E analog, Trolox, alone show an increase in leukemia cell growth at the 24-h time point, with no significant interaction with DHA. Vitamin E has been shown to decrease the anti-angiogenic effects of artemisinin (26); thus, it is pertinent to study how it interacts with DHA in vivo, where angiogenesis is a major factor in tumor growth.
Dexamethasone is shown to induce oxidative stress-mediated apoptosis in lymphoid tissue (27). Our results demonstrate that dexamethasone reduces leukemia cell growth. There was no significant interaction with DHA. Due to the anti-inflammatory nature of dexamethasone, it will be important to further study its interaction with DHA in vivo, where inflammation is a key factor in tumor modulation by the immune system.
H2O2 is a known signaling molecule and strong oxidizing agent. Normal human plasma concentration of H2O2 is approximately 11.1 μM (31) and can be increased with exercise (32). Our H2O2 results indicate that H2O2 kills leukemia cells alone. Furthermore, the interaction between H2O2 and DHA was found to be additive, which may be due to the similar methods of apoptosis mediation by ROS generation employed by both compounds.
We have explored the interaction effects of several nutritional supplements and natural compounds with antioxidant and pro-oxidant properties with DHA on Molt-4 cells. Some of these compounds could increase or decrease the anticancer effectiveness of artemisinin in vitro. We do not yet know the therapeutic values of our in vitro findings; however, implications of this work could pave the way for setting new treatment regiments for patients undergoing artemisinin therapy for cancer. Our goal is to understand variability between patients and, hopefully, use this knowledge to reduce the easily achievable dosage of artemisinin required for successful cancer treatment. Our strategy presents an attractive option for patients who are looking to optimize their treatments through nutritional modulation. Future in vivo research is needed to confirm our findings in animals and humans and to study more complex systemic interactions with the supplement molecules.
Footnotes
-
Conflicts of Interest
The Authors declare no conflicts of interest with respect to this study.
-
Funding
Funding for this research was provided in part by the Dean of the College of Engineering and Associate Vice Provost for Research of the University of Washington and the Washington State Life Science Discovery Fund (LSDF).
- Received December 15, 2014.
- Revision received January 14, 2015.
- Accepted January 16, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.