


Abstract
Background: Active hexose-correlated compound (AHCC) is an extract of a basidiomycete mushroom that enhances the therapeutic effects and reduces the side-effects of chemotherapy. Our previous studies demonstrated that heat-shock protein 27 (HSP27) was involved in gemcitabine-resistance of pancreatic cancer cells and it was down-regulated by AHCC-treatment. However, how AHCC down-regulated HSP27 is unknown. In the present study, we focused on two transcription factors reported to induce HSP27, heat shock factor 1 (HSF1) and high-mobility group box 1 (HMGB1) and investigated the effect of AHCC on their expression. Materials and Methods: KLM1-R cells were treated with AHCC and the protein expression of HSF1 and HMGB1 were analyzed by western blotting. Results: The protein expression of HSF1 in KLM1-R was down-regulated by AHCC treatment. On the other hand, the protein expression of HMGB1 was not reduced in KLM1-R cells after AHCC treatment. Conclusion: The possibility that AHCC down-regulated HSP27 through down-regulation of the HSF1, was herein shown.
Pancreatic cancer has a poor prognosis, and the 5-year overall survival rate is less than 5%. It is the fifth leading cause of cancer death (1, 2). Surgical resection is the only radical treatment, but most patients are not diagnosed until an advanced, unresectable stage (3). Gemcitabine (2′-deoxy-2′,2′-difluorocytidine monohydrochloride) improved the quality of life in many patients and moderately extended survival, and thus gemcitabine is currently one of the standard drugs for patients with pancreatic cancer (4). However, usually gemcitabine therapy is interrupted by intrinsic or acquired resistance of pancreatic cancer (5).
In our previous studies, we compared protein expression of the gemcitabine-resistant human pancreatic cancer cell line, KLM1-R, against those of sensitive parental KLM1, revealing that heat shock protein 27 (HSP27) was up-regulated in KLM1-R compared to KLM1 (6, 7). Furthermore, since down-regulation of HSP27 in KLM1-R elevated the cytotoxic effect of gemcitabine, HSP27 is thought to be involved in gemcitabine resistance (8-10).
Active hexose-correlated compound (AHCC) is derived from the basidiomycete mushroom Lentinula edodes that is composed of polysaccharides, amino acids, lipids and minerals. The principal components of AHCC are oligosaccharides and α1, 4-glucans occupy the major portions of these. AHCC has been reported to have immunomodulatory and antitumor effects (11). We previously showed that AHCC down-regulated HSP27 in KLM1-R cells in vitro (12).
Heat shock factor 1 (HSF1) is a transcription factor that is a regulator of heat-shock proteins, including HSP27 (13). Recently, several studies have reported that HSF1 has been found increased in several human cancers and was essential for the proliferation of cancer cells (14-16). High-mobility group box 1 (HMGB1) is also involved in the regulation of HSP27 (17, 18). HMGB1 is a nuclear factor that is important for transcription (19). HMGB1 is expressed following chemotherapy and depletion of HMGB1 increased sensitivity to anticancer chemotherapy (20, 21). However, whether AHCC-treatment affect HSF1 or HMGB1 in KLM1-R cells is unknown. To investigate the effect of AHCC on HSF1 and HMGB1 in KLM1-R cells, we analyzed the expressions of HSF1 and HMGB1 in AHCC-treated or untreated KLM1-R cells by western blot analysis.
Materials and Methods
Cancer cell line and conditions. The gemcitabine-resistant pancreatic cancer cell line, KLM1-R, was kindly provided by the Department of Surgery and Science, Kyushu University Graduate School of Medical Science. KLM1-R cells were established by exposing gemcitabine-sensitive KLM1 cells to gemcitabine (22). KLM1-R cells were incubated in RPMI-1640 medium added 10% fetal bovine serum (inactivated at 56°C for 30 min), 2 mM L-glutamine, 1.5 g/l sodium bicarbonate, 10 mM N-2-hydroxyehylpiperazine-N'-2-ethanesulfonic acid (HEPES), and 1.0 mM sodium pyruvate, and maintained in a humidified 5% carbon dioxide-95% air mixture at 37°C.
Agents. Active hexose-correlated compound (AHCC) was kindly given by the Amino Up Chemical Co., Ltd. (Sapporo, Japan). AHCC was dissolved in RPMI-1640 medium and filter-sterilized.
Sample preparation. KLM1-R cells were treated with or without AHCC (10 mg/ml) for 48 h in vitro. After treatment, the cells were homogenized on ice in lysis buffer [50 mM Tris-HCl, pH 7.5, 165 mM sodium chloride, 10 mM sodium fluoride, 1 mM sodium vanadate, 1 mM phenyl methyl sulfonyl fluoride, 10 mM ethylendiaminetetra-acetic acid, 10 μg/ml aprotinin, 10 μg/ml leupeptin, and 1% nonylphenocypolyehoxylethanol-40]. After centrifugation at 15,000 rpm for 30 min at 4°C, the supernatant was collected and used for western blotting. Protein concentrations were measured by the Lowry method (23).
Western blot analysis. In order to analyze the expression levels of intracellular proteins from KLM1-R cells, 15-μg protein samples were subjected to western blot analysis. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed in pre-cast gels (4-20% gradient polyacrylamide gels; Mini-PROTEAN TGX Gels, Bio-Rad, Hercules, CA, USA). After the samples were separated by electrophoresis, gels were transferred electrophoretically onto polyvinylidene difluoride membranes (Immobilon-P; Millipore, Bedford, MA, USA) and blocked overnight at 4°C with Tris-buffered saline (TBS) containing 5% skimmed milk. The primary antibodies used were: rabbit monoclonal antibody against HSF1 (1:1,000, CST, Beverly, MA, USA), rabbit polyclonal antibody against HMGB1 (1:1,000, CST) and goat polyclonal antibody against actin (1:200, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). Membranes were incubated with the primary antibody overnight at 4°C. The membranes were washed three times with TBS containing 0.05% Tween-20 for 3 min and then incubated with horseradish peroxidase-conjugated secondary antibody (dilution 1:10,000; Jackson Immuno Research Laboratories Inc., West Grove, PA, USA) for 1 h at room temperature. After washing, bands of HSF1, HMGB1 and actin were visualized by an enhanced chemiluminescence system (ImmunoStar Long Detection; Wako, Osaka, Japan), and recorded using an Image Reader LAS-1000 Pro (Fujifilm Corporation, Tokyo, Japan). Expression levels of HSF1, HMGB1 and actin in AHCC-treated or untreated KLM1-R cells were quantified by analyzing the intensity of each band using the Multi Gauge ver. 3.0 software (Fujifilm Corporation).
We calculated the ratio of intensities of HSF1 to actin (HSF1/actin) and HMGB1 to actin (HMGB1/actin) in AHCC-treated or untreated KLM1R cells from four independent experiments. The ratio of band intensities in untreated KLM1-R cells was considered 100%. Statistical significance of differences in the ratio of intensities of HSF1/actin and HMGB1/actin in AHCC-treated or untreated KLM1R cells were calculated by the Student's t-test. A value of p<0.05 was considered statistically significant.
Results
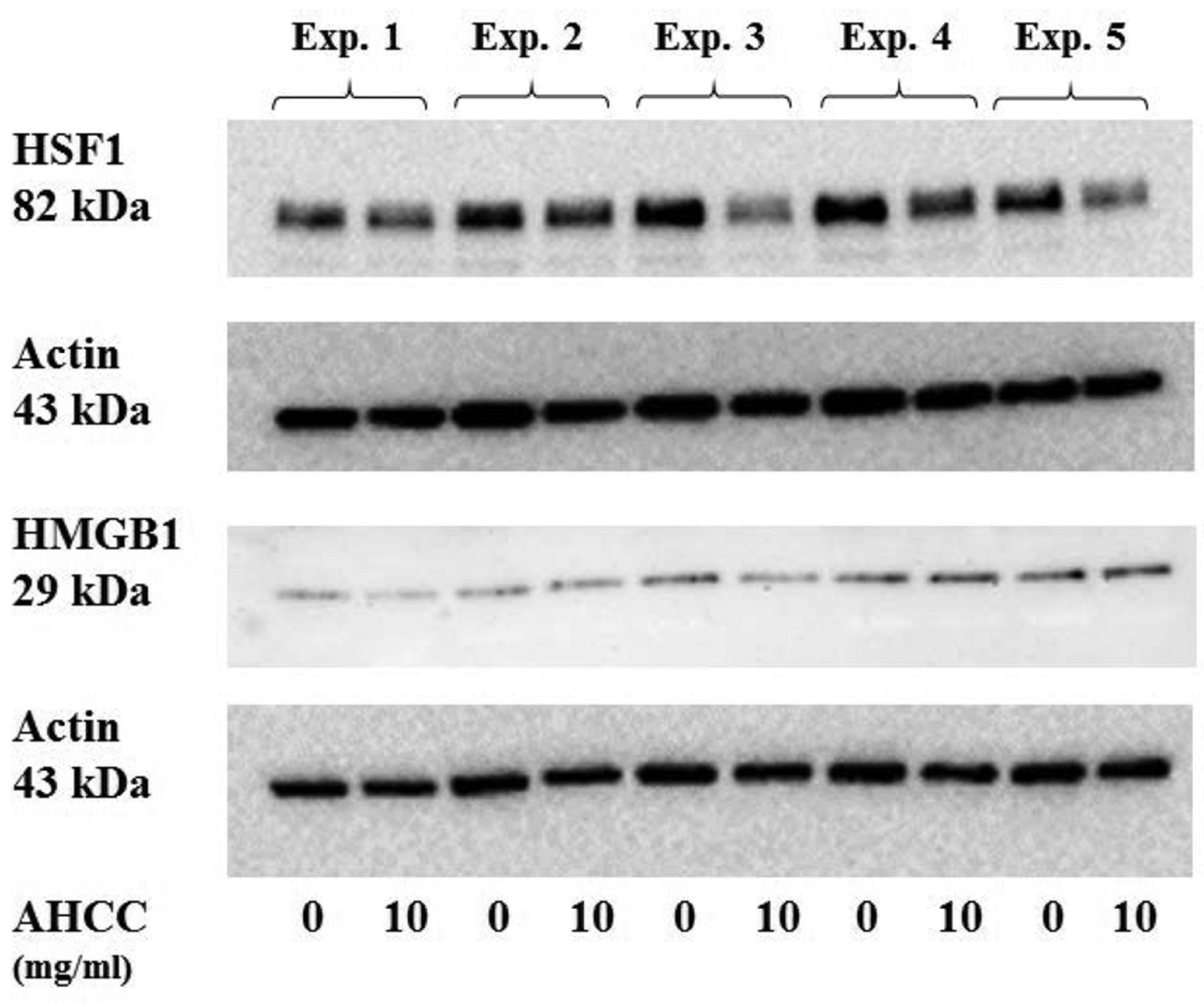
In order to evaluate the effect of AHCC on the HSF1 and HMGB1, we analyzed the intracellular proteins from KLM1-R cells treated with or without AHCC by western blot analysis with a primary antibody against HSF1, HMGB1 and actin (Figure 1). The protein expression of HSF1 was reduced by AHCC treatment in KLM1-R cells, whereas HMGB1 and actin were almost the same in all cells. The ratio of intensities of HSF1/actin and HMGB1/actin in KLM1-R cells were measured (Figure 2). The ratio of band intensities in untreated KLM1-R were considered 100%. The ratio of intensities of HSF1/actin in AHCC-treated or untreated KLM1-R cells were 100%±20.1 and 73.5%±4.6, respectively. On the other hand, the ratio of intensities of HMGB1/actin in AHCC-treated or untreated KLM1-R cells were 100%±26.5 and 102%±50.7, respectively. The ratio of intensity of HSF/actin was significantly different (p<0.05) between AHCC-treated and untreated KLM1-R cells. These results suggested that HSF1, but not HMGB1, was down-regulated by AHCC treatment in vitro.
Discussion
HSPs are induced by various stress factors and are implicated in cell survival, thus controlling the apoptotic signaling pathway. In cancer cells, HSPs are involved in anti-apoptotic function and resistance to chemotherapy. Thus, HSPs have been considered anticancer therapeutic targets in many cancers (24, 25).
In pancreatic cancer cells, we previously showed that interferon-γ and N-formy1-3,4-methylenediosy-benzylidene-γ-butyrolactam (KNK437) reduced the expression of HSP27 to enhance the gemcitabine sensitivity in the gemcitabine-resistant pancreatic cancer cell line KLM1-R (9, 10). HSP90 is also a molecular target of pancreatic cancer therapy (26). HSP90 inhibitors, such as geldanamycin, inhibit many oncogenic client proteins, whereas this agent induces heat shock response and increases HSP27 expression through HSF1 activation (27). HSF1 normally exists as the inactive monomer in unstressed cells. Under stress conditions, HSF1 forms a trimer and translocates to the nucleus to activate the expression of HSPs. In the nucleus, HSF1 is phosphorylated and transcriptionally activated (28). HSF1 is a master transcription factor for heat shock responses and HSPs, thus the inhibition of HSF1 is an anticancer therapy strategy. Recent studies reported that knockdown of HSF1 by siRNA induced apoptosis in pancreatic cancer cell lines (29). The triazole nucleoside analog reduced HSF1 expression and HSPs expression and inhibited tumor formation in pancreatic cancer cell lines (30). Furthermore, high levels of HSF1 are implicated in poor prognosis of breast cancer (31). Herein, we showed that AHCC-treatment significantly down-regulated the expression of HSF1 in a gemcitabine resistant pancreatic cancer cell line in vitro.

Expression levels of HSF1 and HMGB1 in active hexose-correlated compound (AHCC)-treated and -untreated KLM1-R cells. Western blot analysis of HSF1, HMGB1 and actin in AHCC-treated (10 mg/ml) or -untreated (0 mg/ml) gemcitabine-resistant pancreatic cancer KLM1-R cells. The protein expression of HSF1 (bands of 82 kDa) was reduced by AHCC treatment in KLM1-R cells compared to untreated KLM1-R cells. On the other hand, protein expression of HMGB1 (bands of 29 kDa) and actin (bands of 43 kDa) did not change significantly in the cells.
AHCC has been used as an immunotherapeutic agent for cancer patients and healthy volunteers (32, 33). AHCC has attenuated the side-effects of antitumor agents (34, 35). AHCC is thought to enhance the chemotherapeutic efficacy. AHCC plus UFT reduced the metastasis of rat mammary adenocarcinoma cells (36). The present study showed that AHCC down-regulated the expression of HSF1. As described above, HSF1 is activated by phosphorylation at Ser326 (37). Human epidermal growth factor receptor-2 (HER2; ErbB2/Neu) signals induce HSF1 phosphorylation via the PI3k-AKT-mTOR pathway. Recently, it was reported that HER2 induces HSF1 trimerization and phosphorylation and facilitates HSF1 protein synthesis (38, 39). In our previous study, AHCC down-regulated HSP27 expression and enhanced the cytotoxic effect of gemcitabine in vitro (12). We also reported that AHCC down-regulated Sex-determining region Y-box2 (SOX2) of KLM1-R cells (40). SOX2 is part of the sox gene family encoding transcription factors. It has been reported that SOX2 associated with HSP27 and HMGB1 in the regulation of HSP27 expression (17, 41). We thought that AHCC down-regulated the expression of HSP27 through down-regulation of HSF1, although further studies are required to elucidate the mechanism by which it reduces the expression of HSF1. AHCC can be considered a valid candidate for combinatorial therapy in anticancer drug regimens for drug-resistant cancer cells.
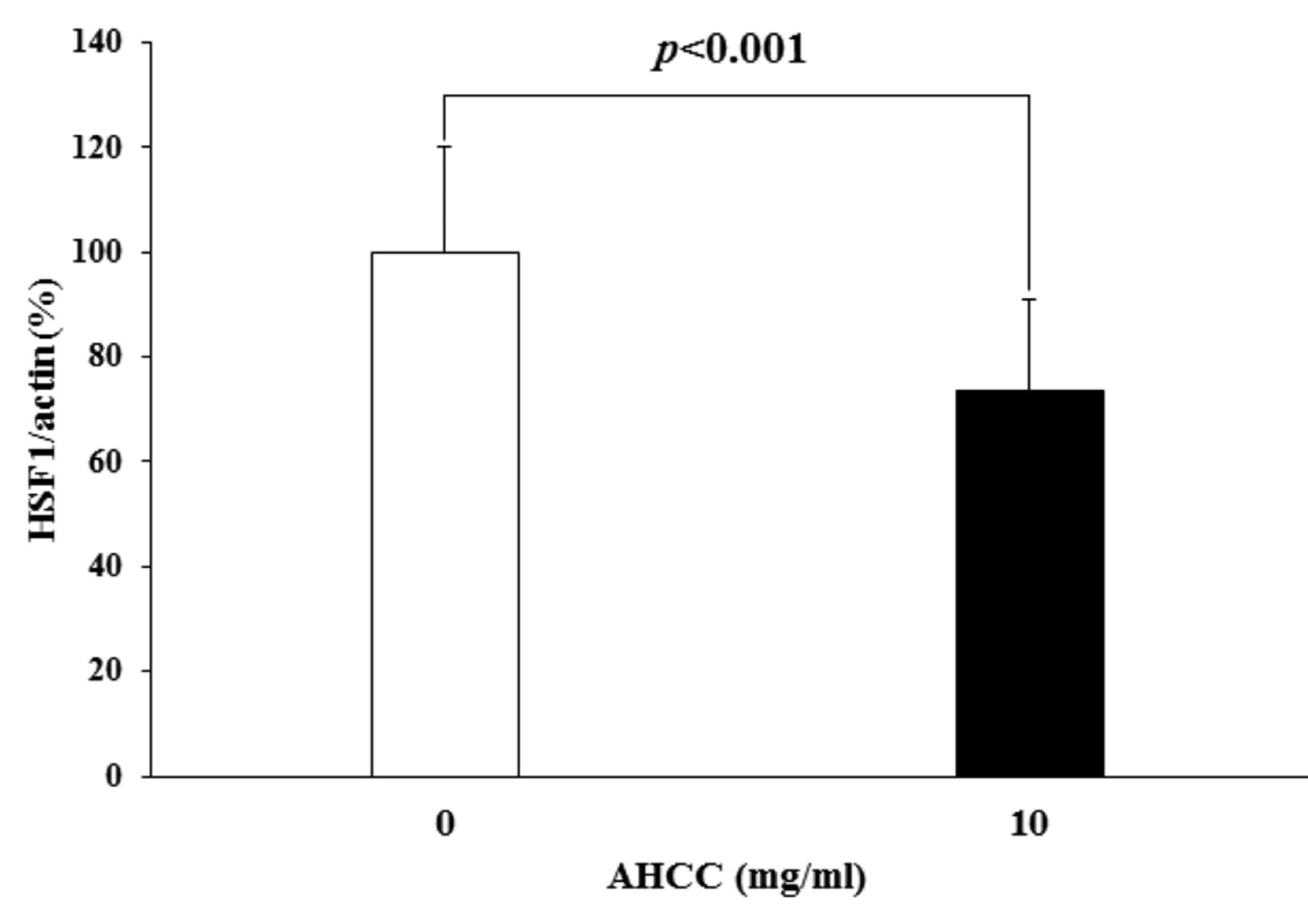
The intensity of the HSF1/actin bands in KLM1-R cells. This graph shows the ratio of the intensities of HSF1 protein to actin protein bands in AHCC-treated or untreated KLM1-R cells. The intensity of HSF1/actin in KLM1-R cells was significantly reduced by AHCC treatment in vitro (p<0.001 by Student's t-test). A value of p<0.05 was considered statistically significant (n=4).
Acknowledgements
The Authors thank Dr. Shin-ichiro Maehara and Prof. Yoshihiko Maehara at Kyushu University to give us KLM1-R cells. Immunoblot detection by LAS-1000 was performed at Gene Research Center of Yamaguchi University.
- Received July 17, 2015.
- Revision received August 27, 2015.
- Accepted August 31, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- A Standardized Extract of Cultured Lentinula edodes mycelia Up-regulates COX-2 in Inflammation-related Malignant Progressive Fibrosarcoma Cell Clone QRsP-11
- CUB Domain-containing Protein 1 (CDCP1) Is Down-regulated by Active Hexose-correlated Compound in Human Pancreatic Cancer Cells
- Enzyme-treated Asparagus Extract Down-regulates Heat Shock Protein 27 of Pancreatic Cancer Cells