


Abstract
Background/Aim: The role of oncogenic or high-risk human papillomavirus (HPV) in cervical carcinogenesis is inevitable, yet not fully understood. Detailed analysis of microRNA (miRNA) alterations occurring during high-risk HPV transformation will increase our current understanding over cervical carcinogenesis. The two main aims of the study were: (i) finding association between HPV infection characteristics and socio-demographic variables, (ii) finding an predictors of clinical outcome. Materials and Methods: The expression levels of different microRNAs (miR-21, miR-27a, miR-34a, miR-155, miR-196a, miR-203) were determined in formalin-fixed paraffin-embedded (FFPE) human HPV-positive cervical dysplastic and tumorous tissue samples using quantitative real-time PCR (qPCR). Sociodemographic and life-style factors were also analyzed. Results: The expression of miR-27a was significantly higher in cervical intraepithelial neoplasia (CIN)2-3 compared to CIN1 (p=0.023) and in squamous cell carcinoma (SCC) compared to CIN2-3 (p=0.033). Moreover, significantly lower levels of miR-34a were detected in CIN2-3 than in CIN1 (p=0.041) and in SCC than in CIN2-3 (p=0.021). Furthermore, we found significant differences in subjects with multiple HPV in miR-27a (p=0.015) and miR-203 (p=0.025) in CIN2-3 compared to CIN1 and miR-21 (p=0.002), mir-27a (p=0.001) and miR-34a (p=0.001) in SCC/CIN2-3. Expression of miR-27a, showing up-regulation in CIN2-3 compared to CIN1 (p=0.028) and miR-34a (down-regulated), correlated with HPV 16 positivity (CIN2-3/CIN1: p=0.027 and SCC/CIN2-3: p=0.036). MiR-34a expression was also significantly altered in connection to smoking status and presence of HPV 16. Conclusion: The demand for additional, alternative molecular biomarkers with prognostic potential is strong. Evaluation of miRNA expression might be helpful to distinguish different cervical lesions and might be able to help in the prediction of HPV infection outcome.
- Cervical intraepithelial neoplasia
- cervical cancer
- microRNA expression
- single and multiple HPV infection
Based on the latest worldwide cancer statistics, cervical cancer is the third most commonly diagnosed cancer in females with approximately 530,000 new cases annually and with 275,100 deaths in 2008 (1). The initial hypothesis concerning the link of human papilloma virus (HPV) infection with the development of cervical cancer was proposed by zur Hausen nearly 40 years ago (2). HPV 16 and 18 are known to be highly associated with cervical cancer (3). HPV 16 is invariably the most common HPV type, regardless of study design and geographical area. HPV 18 is also common but its prevalence and incidence is more variable across populations (4). Other common high-risk (HR) HPV types include: 31, 33, 35, 45, 52 and 58. Together with 16 and 18, these HR HPVs show a combined worldwide relative contribution of above 90% (5). Through the course of this virus-mediated carcinogenesis, the initial, persistent oncogenic or high-risk HPV infection can progress to well-defined precursor lesions of different grade, known as cervical intraepithelial neoplasia (CIN). More than 80% of cervical cancer (CC) cases are histologically squamous cell carcinomas (SCC) preceded by high-grade dysplasia (CIN2-3).
The Pap smear (cytology) was a revolutionary technique with a major public health impact - that has changed dramatically the incidence of cervical cancer since its widespread introduction in screening programs in developed countries. One of its limitations is the lack of sufficient information regarding infection and disease outcome (6) besides the frequently mentioned inter-observer variation (7). On the other hand, with extensive HPV-testing a high rate of insignificant HPV infections are revealed. Even though co-testing (HPV testing combined with cytology) is a reasonable alternative, the need for alternative molecular biomarkers with prognostic value, especially regarding HPV infection outcome, is overwhelming.
MicroRNAs (miRNAs) are small, non-protein-coding RNAs representing a new class of regulatory mechanisms with important functions in a wide range of biological processes. Aberrant or altered miRNA expression has been reported in a variety of human cancers with a wide range of clinical potential. miRNAs are not only tissue-, organ- or cell-specific. Their de-regulation, caused by genetic and/or epigenetic mechanisms, plays a role in malignant transformation (8, 9) as both mechanisms were shown to contribute to the alterations in miRNA expression in cervical cancer. A more detailed analysis of miRNA alterations occurring during high-risk HPV transformation will increase our current understanding over cervical carcinogenesis. In addition, miRNAs are considered to be a better choice for expression studies since they are more accurate for distinguishing disease states than mRNA expression analysis (9). Archival collection of formalin-fixed, paraffin-embedded (FFPE) tissues is widely used for biomarker research due to the link to rich clinical databases and utility. Previous studies have already demonstrated that miRNAs are minimally affected and display reliable expression levels regardless of fixation time or age of tissue blocks (10). Their small size and a possible protein protection by the RNA-induced silencing (RISC) complex are thought to contribute to their stability during fixation and processing (11). Furthermore, a high correlation has been shown by Goswani et al. between miRNA expression profiles in FFPE and fresh-frozen samples (12).
Materials and Methods
Samples. A total of 98 female patients' (admitted between 2009-2014 to the Department of Obstetrics and Gynaecology, University of Pécs) cervical FFPE tissue samples (obtained either by conisation or hysterectomy), known to be high-risk HPV-positive (pre-selection criteria based on previous HPV genotyping) with definitive histological diagnosis of cervical lesions (CIN1, CIN2-3 and invasive cervical cancer), were used in this study. Histological diagnoses were produced by the same gynaecological pathologist. Tissue blocks were retrieved from the Department of Pathology, University of Pécs, according to approved protocols (Ethics Committee of Pécs University: 3440.316-8331/KK41/2009) and were microdissected restraining to selected morphologically distinctive areas. A total of 4 pieces of 10 μm thick FFPE sections were collected and used for further analysis.
Clinical specimens were grouped according to histological diagnosis: 1, mildly dysplastic lesions (CIN1: n=30), which typically spontaneously regress; 2, precancerous lesions (CIN2: n=10 and CIN3: n=20), about 20% would progress to invasive cervical cancer; and 3, squamous cell cervical carcinoma (SCC: n=38). Single (n=65) and multiple (n=33) HR HPV-positive samples were found in every category. Low-risk HPV types (6 and 11) were identified in one case simultaneously with HR HPV positivity.
RNA extraction and microRNA real-time polymerase chain reaction (qPCR) expression analysis. Steps include deparaffinization (xylol, absolute alcohol) prior to total RNA isolation using the Trizol protocol. Reverse transcription and qPCR reactions, along with detection settings, were carried out as described previously (13). The miRNAs investigated in this study (miR-21, miR-27a, miR-34a, miR-155, miR-196a, miR-203) were systematically chosen based on our previous results and published literature (cell lines, clinical specimen) (13-18). Overall, the selected miRNAs were reported to be involved in oncogenesis and progression in cervical and other neoplasms (over- or under-expression). All runs, including no template controls (nuclease-free water), were carried out in triplicate. Inter-run calibrators were used to calculate correction factors to remove the run-to-run differences. Data normalization was performed using 5SrRNA and U6snRNA as endogenous references. Relative miRNA expression levels were compared via the 2−ΔΔCT method.
Statistical methods. Statistical analyses were performed using the IBM SPSS Version 21 (Armonk, New York, USA). Mann-Whitney U-test, unpaired t-test and one-way analysis of variance (ANOVA) were used to compare continuous and categorical variables based on data distribution. Logistic regression analyses were conducted to identify variables (miRNA expressions, sociodemographic and life-style factors) associated with worse pathological outcomes. The level of significance was set at p<0.05.
Results
As far as for the distribution of HR HPV, HPV 16 was the most prevalent (57.53%) considering both single and multiple infections; types 18 and 31 were the second most frequent (10.96%) followed by 56 (9.59%) and types 51, 52 and 58 (8.22%). These findings correspond to the results of the first Hungarian HPV-Center in Budapest from 2007-2011 (19). Galamb et al. detected a relatively low representation (compared to data from developed countries) of HPV 18 along with high prevalence of other, not so common, HPV types. At the same time, the prevalence of types 51 and 31 among patients with cytological abnormality exceeded the expectations based on international data reflecting dominance of types 16, 18 and 45 (3-5). The prevalence of multiple HPV infection was 33.67%, much lower than expected based on previous data (19).
As described previously, the total of 98 samples were grouped according to histological diagnosis (CIN1: 30.61%, CIN2-3: 30.61% and SCC: 38.78%) and the presence of single or multiple HPV positivity. The mean age of the studied population was 36.17 (range: 18-65). Mean age was significantly lower among multi-HPV-positive patients than among mono-HPV-positive individuals (33.30±7.994 compared to 37.98±10.516, p=0.039). The mean age for patients infected with multiple HPV types was lower in every disease stage than the mean age for patients with single HPV infection, with the only exception being the CIN1 group where the mean age and the age range were the same or quite similar for both subcategories (single: 33.2±8.804 and multiple: 33.2±10.207).
Differential expression of miRNAs in dysplastic tissues of different grade. The expression values were calculated according to the 2nd derivative maximum method and normalized to 5S rRNA. The Mann-Whitney U-test was used to evaluate statistical significance.
Differential expression of miRNAs in dysplastic and tumour tissues. The expression values were calculated according to the 2nd derivative maximum method and normalized to 5S rRNA. The Mann-Whitney U-test was used to evaluate statistical significance.

MiRNA-27a levels by patient group (low-grade dysplasia: cervical intraepithelial neoplasia (CIN)1, high-grade dysplasia: CIN2-3, squamous cellular cancer: SCC). Boxes represent the inter-quartile range and the line indicates the median value. Whiskers indicate 95th and 5th percentiles. 5s rRNA was used for normalization. Logistic regression was used to evaluate statistical significance (p' refers to multiple HPV infection).
Significant associations were found as the expression of miR-27a increased with progression: CIN2-3 related to CIN1 (p=0.023) and SCC to CIN2-3 (p=0.033). MiR-155 also showed an increase in the transition from CIN1 to SCC with statistically significant correlation in CIN2-3 over CIN1 (p=0.023). MiR-196a expression showed an increase from CIN1 to CIN2-3 but was significantly lower in SCC than in CIN2-3 (p=0.016). These associations were unaffected by differences in the type or number of HPV. Even though we registered elevations in miR-21 expression through every disease stage from the progression of CIN1 to CC, the differences were not significant. On the other hand, significantly lower levels of miR-34a were detected in CIN2-3 than in CIN1 (p=0.041) and in SCC than in CIN2-3 (p=0.021) (Tables I and II).
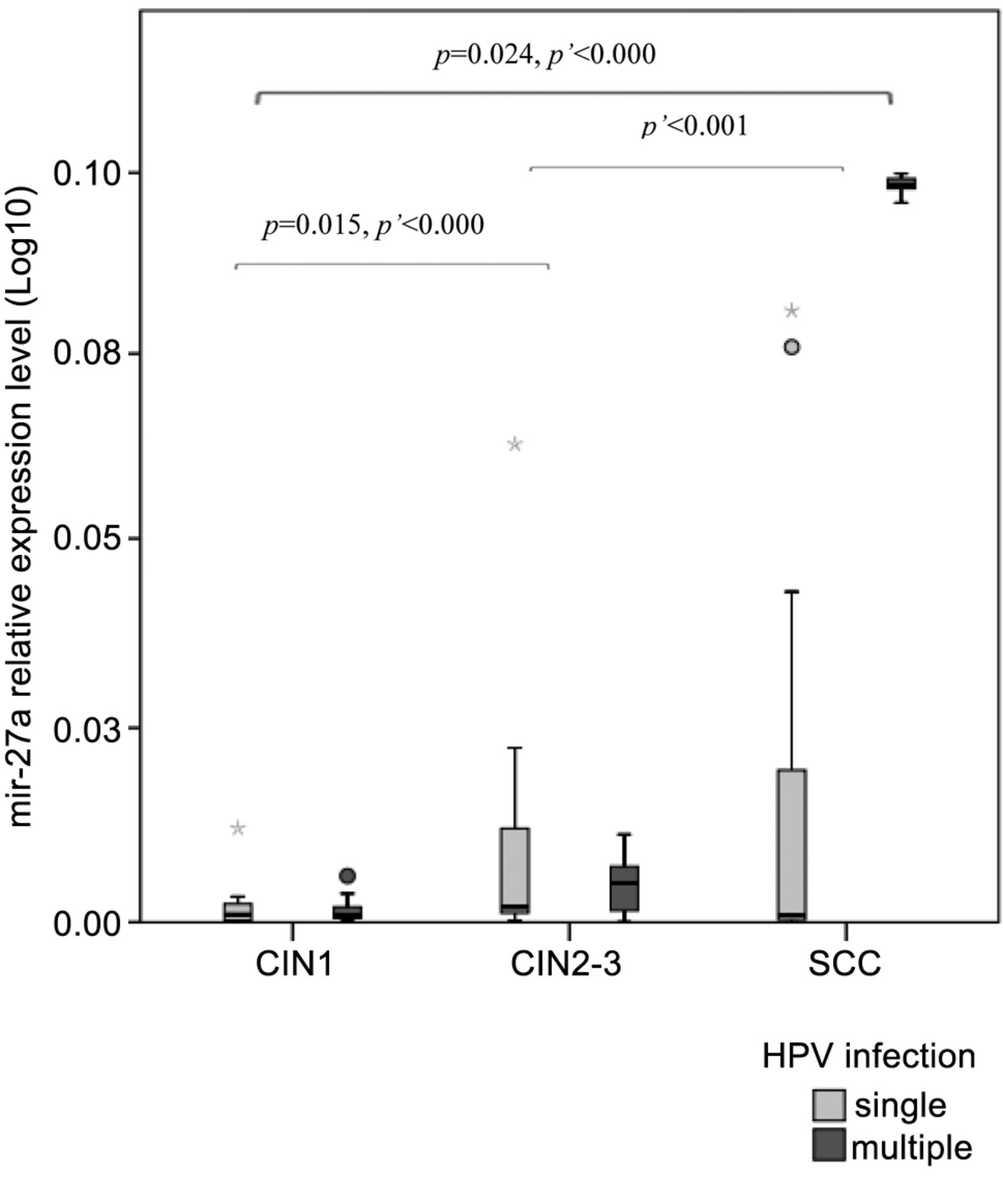
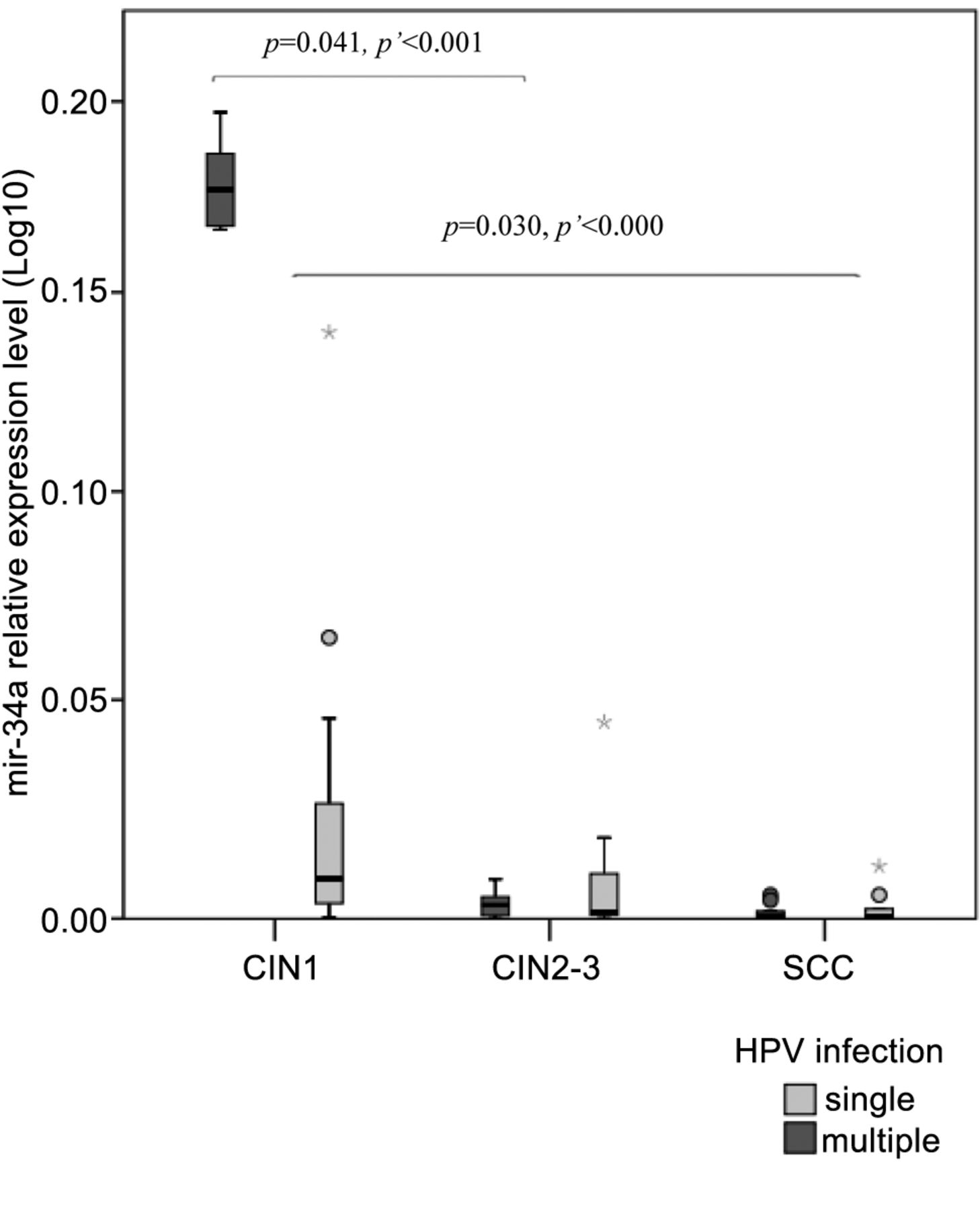
Furthermore, we found significant differences in subjects with multiple HPV for miR-27a (p<0.000), miR-34a (p<0.001) and miR-203 (p=0.025) in CIN2-3 compared to CIN1 and for miR-21 (p=0.002) and mir-27a (p<0.001) in SCC/CIN2-3 with miR-21 (p<0.000), miR-27a (p<0.000) and miR-34a (p<0.000) in CIN1/SCC. The analysis regarding the correlation between miRNA expression and the role of single or multiple HPV showed significant association with disease outcome in the case of miR-27a (p=0.029) and miR-34a (p=0.045) (Figures 1 and 2, respectively).

MiRNA-34a levels by patient group (low-grade dysplasia: cervical intraepithelial neoplasia (CIN)1, high-grade dysplasia: CIN2-3, squamous cellular cancer: SCC). Boxes represent the inter-quartile range and the line indicates the median value. Whiskers indicate 95th and 5th percentiles. 5S rRNA was used for normalization. Logistic regression was used to evaluate statistical significance (p refers to multiple HPV infection).
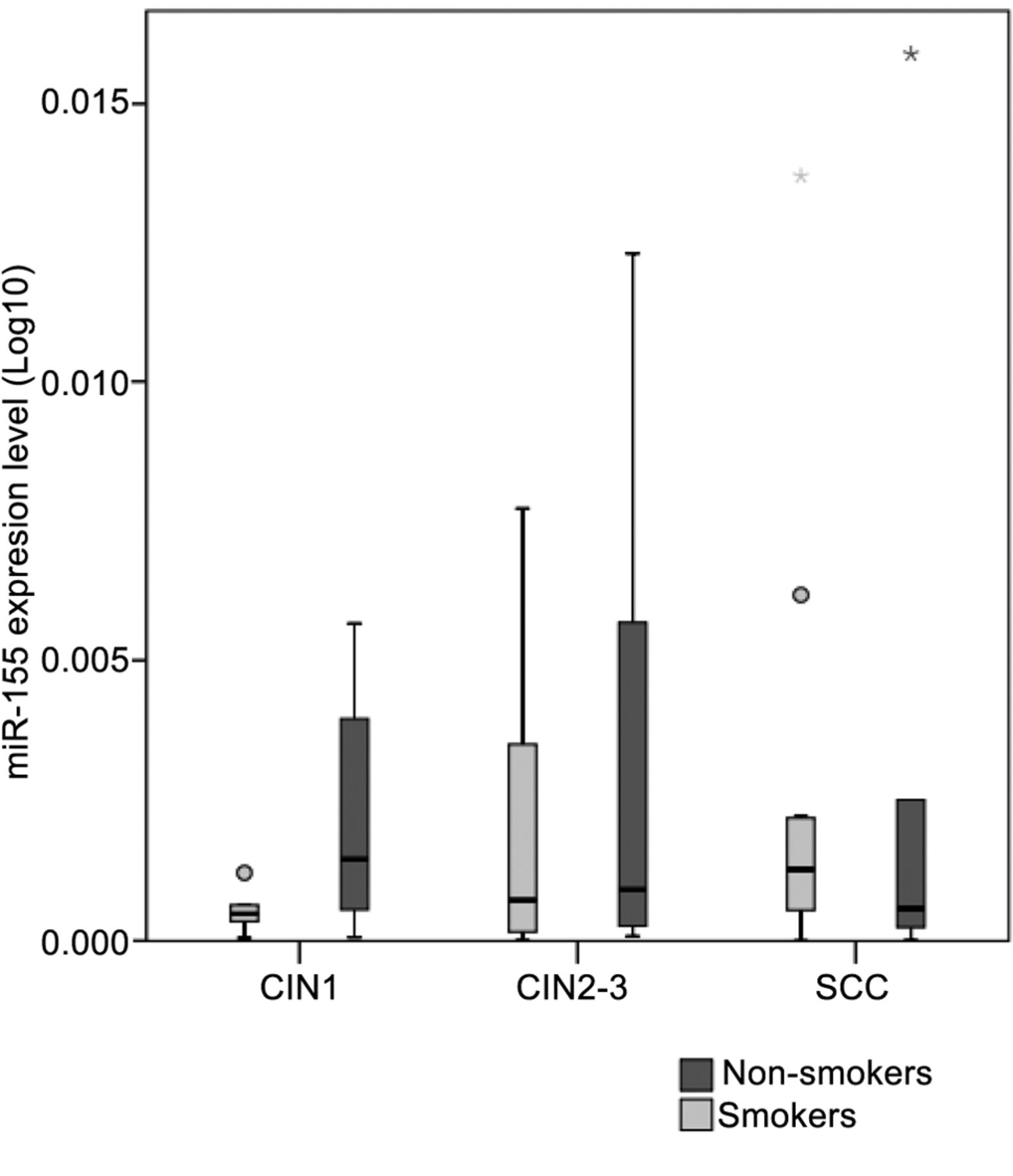
Regarding other factors, we focused on cigarette smoking and HR HPV type. The presence of HPV 16 has a significant effect on disease outcome (p=0.004) (CIN1, 40%; CIN2-3, 63.33%; SCC, 65.22%). During the transition from CIN1 to CIN2-3, miR-27a and miR-34a correlated with HPV 16 positivity (p=0.028 and p=0.027, respectively). As for the progression of CIN2-3 to SCC, in correlation with HPV 16, miR-34a reached significance (p=0.036) (Figure 3). Our study population had a rate of cigarette smoking at 65% among women with CIN1, 46.67% among women with CIN2-3 and 53.33% among CC subjects. Alteration in miRNA expressions between smokers and non-smokers appeared statistically significant in the case of miR-27a and miR-34a among CIN1 subjects (p=0.036 and p=0.029, respectively) (Figure 4). When, based on smoking history, the expressions of miR-155 were compared, opposite trends with relative increase among non-smokers and a decrease among smokers in the transition from pre-malignant dysplasia to cancer were shown (Figure 5).

MiRNA-34a levels by patient group (low-grade dysplasia: cervical intraepithelial neoplasia (CIN)1, high-grade dysplasia: CIN2-3, squamous cellular cancer: SCC). Boxes represent the inter-quartile range and the line indicates the median value. Whiskers indicate 95th and 5th percentiles. 5S rRNA was used for normalization. Logistic regression was used to evaluate statistical significance. Transition from CIN1 to CIN2-3: miR-34a correlated with HPV 16 positivity (p=0.027), CIN2-3 to SCC: p=0.036.
Discussion
De-regulation of microRNAs, in connection with malignant transformation, is well known. The etiological relevance of high-risk HPV types in the development of cervical cancer has been demonstrated by a series of epidemiological and biological studies during the past few decades. Nevertheless, a high-risk HPV infection does not inevitably progress to invasive cervical cancer. Multiple HPV genotypes have been found to be a risk factor for persistent infection in healthy young women and co-infections involving HPV 16 and 58 seemed particularly prone to increased risk (4). The phenomenon that we observed in the case of miR-27a and miR-34a, in correlation with HPV 16 positivity during the transition from low-grade to high-grade dysplasia, seems to reinforce this previous observation. Alterations in the expressions of both oncogenic and tumor suppressor miRNAs (miR-27a (Figure 1) and miR-34a (Figure 2)) also verify the opinion that multiple HPV types appear to act synergistically in cervical carcinogenesis (20, 21).

MiRNA-34a levels by patient group (low-grade dysplasia: cervical intraepithelial neoplasia (CIN)1, high-grade dysplasia: CIN2-3, squamous cellular cancer: SCC). Boxes represent the inter-quartile range and the line indicates the median value. Whiskers indicate 95th and 5th percentiles. 5S rRNA was used for normalization. The Mann-Whitney U-test was used to evaluate statistical significance. MiR-34a expression was significantly lower in smokers than non-smokers, p=0.029.
Previous studies have used different types of samples (cultured cells, cancer-derived cell-lines, human tissue samples - fresh-frozen or FFPE, serum) for the analysis of miRNA expression profiles related to cervical cancer (13-18) and, despite the overlapping results obtained, there is a considerable amount of variability (16, 17).
In our study, we wanted to distinguish characteristic alterations in miR-21, miR-27a, miR-34a, miR-155, miR-196a, miR-203 expression in different stages of cervical dysplasia and SCC. MiR-27a is an oncogenic miRNA modulated by p53, E2F and c-Myc. Both HR HPV E6 and E7 interact with c-Myc and augment c-Myc transactivation activities (18). We found that in HR HPV positive tissue samples miR-27a expression correlated with the progression of high grade cervical lesions to cancer making it a credible biomarker for HR HPV-associated cervical carcinogenesis.A similar phenomenon was observed with higher level of miR-27a in CC compared with CIN3, CIN1+2 or normal cervical tissues in oncogenic HPV-infected raft tissues by Wang et al. (22). Our results are unique regarding associations with single and multiple HPV positivity.
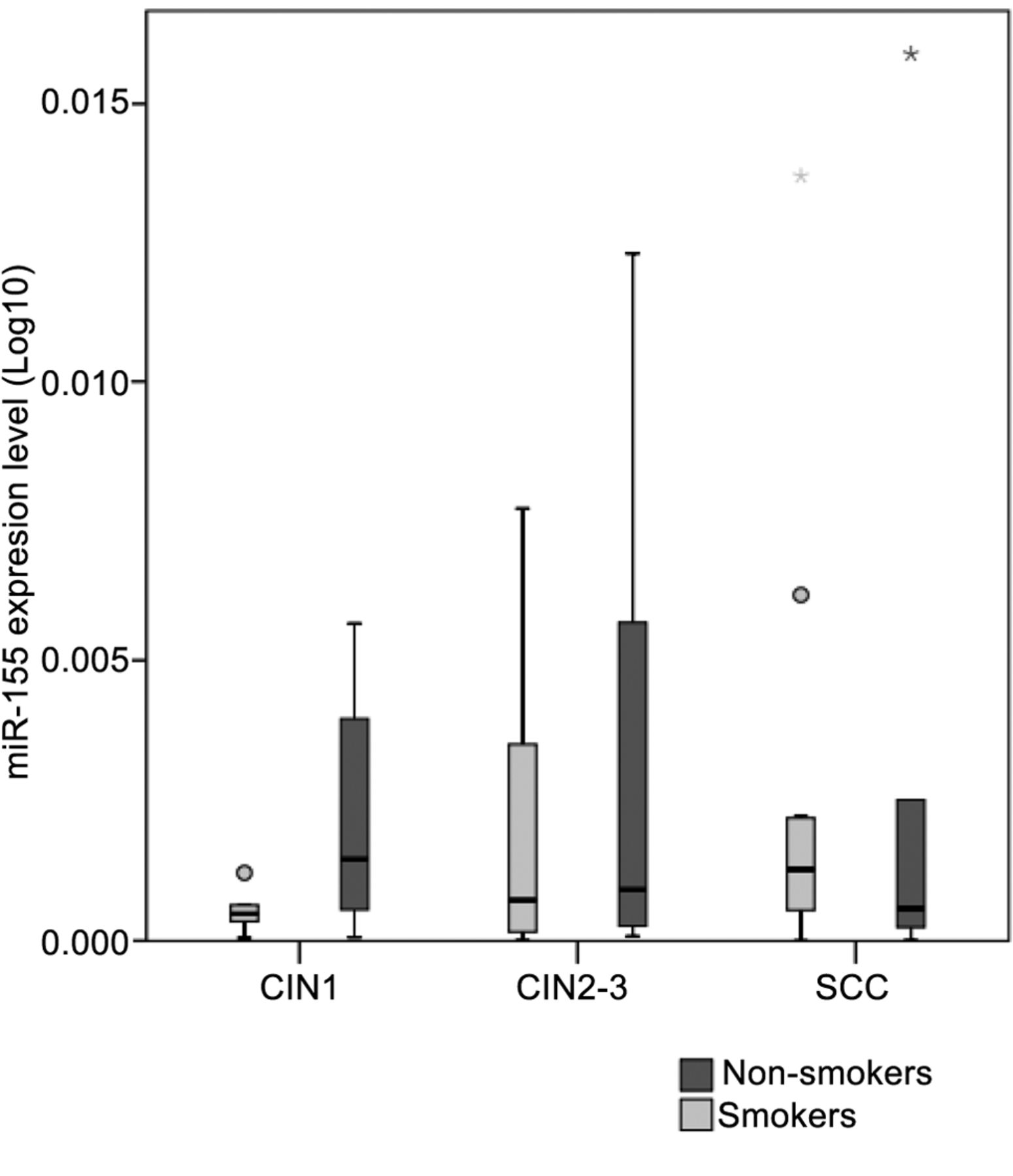
MiRNA-155 levels by patient group (low-grade dysplasia: cervical intraepithelial neoplasia (CIN)1, high-grade dysplasia: CIN2-3, squamous cellular cancer: SCC). Boxes represent the inter-quartile range and the line indicates the median value. Whiskers indicate 95th and 5th percentiles. 5s rRNA was used for normalization.
MiR-34a is directly regulated by p53 and acts as a tumour suppressor, while HR HPV E6 induces the inhibition of miR-34a through the p53-pathway. It is a downstream target of the p53-network with key regulatory functions in cellular apoptosis, G1-arrest, DNA repair and senescence, which are essential for the maintenance of cellular stability and stress response. Li et al. have demonstrated that the expression of miR-34a is significantly lower in HR HPV-infected tissues (23). Our observations clearly confirm that miR-34a acts as a tumour suppressive miRNA in HPV-induced cervical transformation. We also note that alterations in miR-34a expression are associated with the presence of single or multiple HR HPV types supporting the theory of synergism on multiple HPV infection. Cigarette smoke-induced dysregulation of miRNA expression has been demonstrated in both rats (24) and humans but mainly focusing on lung cells or the bronchial airway epithelium (25). One of the miRNAs significantly dysregulated by cigarette smoke (CS) and implicated in lung carcinogenesis is miR-34a, causing disruption of transcription. Smoke seems to leave a unique “molecular signature” in the p53 gene in smoke-exposed lung tumours (26). In the light of these observations, we can rightly hypothesize that CS might influence miRNA regulatory mechanisms in other organs, such as the cervix as well. Based on these findings, we can conclude that miR-34a could help to predict HPV infection outcome and assist in distinguishing different cervical lesions.
The fact that the expression of miR-155 correlated with the progression of low grade to high grade cervical lesions makes miR-155 eligible as a possible biomarker for HR HPV-associated cervical carcinogenesis. Even though we did not find a significant correlation, the difference in miR-155 expression linked to smoking history must be mentioned. Functionally miR-155 mediates immune response by shaping the transcriptome of lymphoid cells that control diverse biological functions vital in adaptive immunity (27). Its altered expression has also been implicated in viral infections and persistence (Epstein-Barr Virus) (28). Lao et al. registered up-regulated expression of miR-155 in cervical cancer tissues compared to adjacent non-cancer tissues. They found that over-expression of miR-155 promoted the proliferation of Hela and SiHa cells and the down-regulation of miR-155 inhibited the growth of cervical cancer cells and promoted apoptosis and induced cell cycle arrest. They also suggested that miRNA-155 promoted the proliferation of cervical cancer cells by regulating tumour suppressor liver kinase B1 (LKB1) expression (29). Functional studies revealed that miR-155 directly targets the tumour protein 53-induced nuclear protein 1 (TP53INP1) and suppresses apoptosis in tumour cells (30, 31). MiR-155 over-expression has been observed in other types of tumours, including oral SCC (OSCC) (32) and head and neck squamous cell carcinomas (HNSCC) (33), also known to be associated with HPV infection.
With respect to variations of miR-196a expression during HPV-mediated cervical carcinogenesis, we confirmed previously reported alterations in tissue samples (16). We found that these alterations were uninfluenced by the dimension of HPV infection (single-multiple, HPV 16 or other). Villegas-Ruiz et al. have validated HOXC8, involved in embryonic development and cellular remodelling, as a target of miR-196a in CC (34). Hou et al. suggested that up-regulation of miR-196a enhanced G1/S-phase transition and the proliferative ability of cervical cancer cells through the regulation of the phosphoinositide 3-kinase (PI3K)/Akt signalling pathway (35). It is important to note that miR-196a also plays a distinct role in the regulation of cell proliferation in HNSCC (36). Wu et al. have demonstrated that miR-196a might be involved in tumour invasion and in the interactions of tumour microenvironment in patients with squamous cell lung carcinoma (37). In the case of OSCC, high expression of miR-196a was associated with tumour recurrence, nodal metastasis, mortality and poor survival (38).
We have proven, in conjunction with Pereira et al., that miR-203 shows a relative reduction in expression during the transition from normal tissue to atypical dysplasia and cancer (16). This observation is due to the assumption that high levels of miR-203 (targeting, amongst others, the p63 family of transcription factors) are inhibitory to HPV amplification and that HPV proteins (E7 in particular) act to suppress expression of this microRNA to allow productive replication in differentiating cells (39). It has also been indicated that miR-203 down-regulation in cervical cancer is caused by promoter hypermethylation (40). Investigations regarding invasion and metastasis showed that low expression of miR-203 correlated with lymph node metastasis (41).
Acknowledgements
We thank Zsuzsanna Brunner and Monika Herczeg for their excellent (precise, coherent) technical assistance. This research was supported by the European Union and the State of Hungary, co-financed by the European Social Fund in the framework of TÁMOP 4.2.4.A/2-11-1-2012-0001 ‘National Excellence Program’.
Footnotes
-
Conflicts of Interest
The Authors have declared that no competing interests exist.
- Received September 19, 2014.
- Revision received October 15, 2014.
- Accepted October 21, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.