


Abstract
Background/Aim: The cancer stem cell (CSC) model postulates the existence of a small proportion of cancer cells capable of sustaining tumor formation, self-renewal and differentiation. Signal Transducer and Activator of Transcription 3 (STAT3) signaling is known to be selectively activated in breast CSC populations. However, it is yet to be determined which molecular mechanisms regulate STAT3 signaling in CSCs and what chemopreventive agents are effective for suppressing CSC growth. The aim of this study was to examine the potential efficacy of curcumin and epigallocatechin gallate (EGCG) against CSC and to uncover the molecular mechanisms of their anticancer effects. Materials and Methods: To suppress the CSC phenotype, two breast cancer cell lines (MDA-MB-231 cells and MCF7 cells transfected with HER2) were treated with curcumin (10 μM) with or without EGCG (10 μM) for 48 h. We used tumor-sphere formation and wound-healing assays to determine CSC phenotype. To quantify CSC populations, Fluorescence-activated cell sorting profiling was monitored. STAT3 phosphorylation and interaction with Nuclear Factor-kB (NFkB) were analyzed by performing western blot and immunoprecipitation assays. Results: Combined curcumin and EGCG treatment reduced the cancer stem-like Cluster of differentiation 44 (CD44)-positive cell population. Western blot and immunoprecipitation analyses revealed that curcumin and EGCG specifically inhibited STAT3 phosphorylation and STAT3-NFkB interaction was retained. Conclusion: This study suggests that curcumin and EGCG function as antitumor agents for suppressing breast CSCs. STAT3 and NFκB signaling pathways could serve as targets for reducing CSCs leading to novel targeted-therapy for treating breast cancer.
Breast cancer is the most frequent malignancy and a leading cause of cancer-related death in women around the world (1). The initiation and progression of breast cancer is a complicated process that results from the loss of the normal regulatory pathways between cell proliferation, differentiation and apoptosis. Previous studies have identified a small subset of tumor-initiating cells within tumors that drive tumor growth and recurrence, termed cancer stem cells (CSCs) (2, 3). CSCs possess self-renewal capabilities, the ability to resist standard chemotherapy, and to generate tumor growth and metastasis. Several signaling pathways have been implicated in the self-renewal behavior of CSCs, including the Wnt (Wingless-Int), Notch and Hedgehog pathways (4-6). Since CSCs possess resistance to conventional chemo- and radiotherapy, new drugs and novel therapies need to be devised to target CSCs in breast cancer.
Signal transducer and activator of transcription 3 (STAT3) is a transcription factor that conveys various signals of cytokines and growth factors from the cell membrane to the nucleus (7). In a variety of human malignancies, including breast cancer, constitutive activation of STAT3 is correlated with the tumor progression and a poor prognosis (8). Recent studies with human lung and breast cancer tissues demonstrated that activated STAT3 was the crucial contributor to tumor invasion (9, 10). However, the molecular mechanisms by which STAT3 promotes CSC traits in breast tumors, as well as the potential contributions of STAT3 to metastasis, have yet to be elucidated.
Several natural products have been suggested as effective agents for cancer prevention, including curcumin and epigallocatechin-3-gallate (EGCG) (11, 12). Curcumin is the principal curcuminoid of the popular Indian spice turmeric, which is a member of the ginger family. Curcumin has been reported to exhibit antioxidant, anti-inflammatory, antimicrobial and anticancer activity in vitro and in animal models of disease (13). Its anticancer activity is exerted through modulation or inhibition of multiple molecular pathways (14-16). Notably, curcumin is a potent inhibitor of NFκB, a transcription factor found to play a role in tumorigenesis of many human malignancies (17). Curcumin has shown dose-dependent chemopreventive and chemotherapeutic effects in numerous carcinogenesis models and in pre-clinical trials (18).
The natural product EGCG is a major catechin in green tea that possesses antioxidant, anti-inflammatory, anticancer, and immunomodulatory properties (19, 20). Studies conducted on cell-culture systems and animal models, as well as human epidemiological studies, show that EGCG in green tea could afford protection against a variety of cancer types (21). Many studies have shown that EGCG produces an anticancer effect by modulating activity of mitogen-activated protein kinases (MAPKs), Insulin-like growth factor 1 (IGF1) receptor, Akt, NFκB and hypoxia-inducible factor 1α (HIF1α) (22-25). However, the investigations on green tea or EGCG in breast cancer using animal models are limited, and the role of EGCG in breast cancer therapy is poorly understood. Herein we studied the curcumin and EGCG anti-CSC effects and the underlying molecular mechanisms in human breast cancer cells.
Materials and Methods
Cancer cell lines and culture. MDA-MB-231 and wild-type MCF7 cell lines were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). They were maintained in a monolayer culture in Dulbecco's modified Eagle's medium (DMEM)/F12 with 10% fetal bovine serum, 2.5% L-glutamine and 0.5% penicillin/streptomycin. The MCF7, transfected with HER2 (human epidermal growth factor 2), (MCF7-HER2) cell line was a generous gift from Dr. C. Kent Osborne (Baylor College of Medicine, Houston, TX, USA). MCF7-HER2 cells were maintained in a monolayer culture in DMEM 1× with 10% fetal bovine serum, 2.5% L-glutamine, 0.5% penicillin/streptomycin, and G418 (400 μg/ml).
Curcumin and EGCG treatment. Both curcumin and EGCG were purchased from Sigma-Aldrich (Saint Louis, MO, USA). Curcumin (10 μM) and EGCG (10 μM) were either treated alone or combined in the tumor-sphere formation, wound healing assay and flow cytometry analyses. The vehicle solvent dimethyl sulfoxide (DMSO) was used as a negative control. Each assay was repeated three times and statistically significant differences were analyzed.
Tumor-sphere formation assay. 200 μl of matrigel (BD Biotech, Cambridge, MA, USA) were spread as a thick layer on a 24-well plate and allowed to polymerize at 37°C for 15 min. Cells (2×104) grown as monolayer were trypsinized to single cells and plated on top of the pre-coated matrigel. Plates were incubated at 37°C to allow cells to fully settle before the media were replaced with appropriate culture media containing 5% matrigel. Cells were grown for 15 days; fresh growth media with matrigel was replenished every two days. Images of representative fields were taken at day 7.
Wound-healing assay. Cells (1×105 cells/well) were plated in growth medium in 24-well plates and incubated for 24 h. After confirming the formation of a complete monolayer, the cells were wounded by scratching lines with a standard 200-μl plastic tip. Migration and cell movement throughout the wound area was observed with a phase-contrast microscope after 24 h.
Western blot analyses. Monolayer cultures of cell lines at an 80-90% confluence were lysed using 100 μl of RIPA buffer (Thomas Scientific, Swedesboro, NJ, USA). Tris-glycine (Bio-Rad, Irvine, CA, USA) gels were loaded with 50-100 μg of lysates. Following electrophoresis, the gel was transferred to a nitrocellulose membrane for 2 h. The membrane was blocked for 1 hour in 5% BSA (bovine serum albumin) or 5% skim milk at 4°C. The membrane was then washed three times with 1×TTBS (tween Tris buffered saline) and incubated overnight with primary antibodies at 4°C. Human TERT (telomerase reverse transcriptase) primary antibody was purchased from Millipore (MABE 14; Billerica, MA, USA). Primary antibodies of STAT3, pSTAT3 and β-actin were purchased from Cell Signaling Technology (Danvers, MA, USA).
Immunoprecipitation. Cells were washed once with PBS buffer and lysed in immunoprecipitation lysis buffer (Thomas Scientific Company; Swedesboro, NJ, USA). STAT3 antibody was added to the cell lysates and incubated for 1 h at 4°C. 70 μl of protein A agarose (Santa Cruz Biotech; Santa Cruz, CA, USA) was added to the cell lysates and incubated overnight at 4°C. After the immunoprecipitation, cell lysates were briefly spun and washed three times with PBS buffer. Immuno-pellets were resuspended in 100 μl of loading dye and resolved by 7.5% polyacrylamide gel electrophoresis.
Flow cytometry. Approximately 500,000 MDA-MB-231 cells were washed with 1×PBS, trypsinized and then transferred to a 15 ml tube. Cell suspensions were spun down, re-suspended in 2 ml of 1×PBS and then divided into two 0.5 ml tubes for staining. One tube was used as an unstained control and the other one as stained with 10 μl of antibody to CD44 (FITC; Fluorescein isothiocyanate, Green; BD Biotech, Cambridge, MA, USA). The tubes were shaken briefly and incubated at room temperature for 15 min in the dark. Each tube was then washed with 3.5 ml of 1×PBS and then centrifuged for 6 min. After aspirating the supernatant, the cells were re-suspended in 3 ml of 1×PBS and subjected to FACS (Fluorescence activated cell sorting) profiling at the UCLA FACS Core laboratory (Los Angeles, CA, USA).
Results
In vitro tumorigenesis was inhibited by combined treatment of curcumin and EGCG. First, we examined whether curcumin or EGCG inhibit the tumor-sphere forming capability of breast cancer cells. To test this, we cultured breast cancer cells in a 3-dimensional microenvironment in the absence and presence of curcumin (10 μM), EGCG (10 μM) alone or combined for 14 days and then counted the number of multi-cellular tumor spheres formed under 3-dimensional culture conditions. MCF7-HER2 and MDA-MB-231 cells formed tumor-spheres after five days (Figure 1A). MCF7-HER2 cells formed ~12 tumor spheres per well, whereas curcumin- and EGCG-treated cells formed 8 and 9 tumor spheres per well, respectively. However, combined treatment of curcumin and EGCG clearly reduced tumor-sphere formation, to 3 spheres per well (Figure 1B). Similarly, MDA-MB-231 cells formed 6 tumor spheres per well, while combined treatment of curcumin and EGCG reduced this to an average of 2 per well (Figure 1C). Our data suggest the combined treatment of curcumin and EGCG effectively inhibits tumor-sphere formation in the two breast cancer cell lines tested.
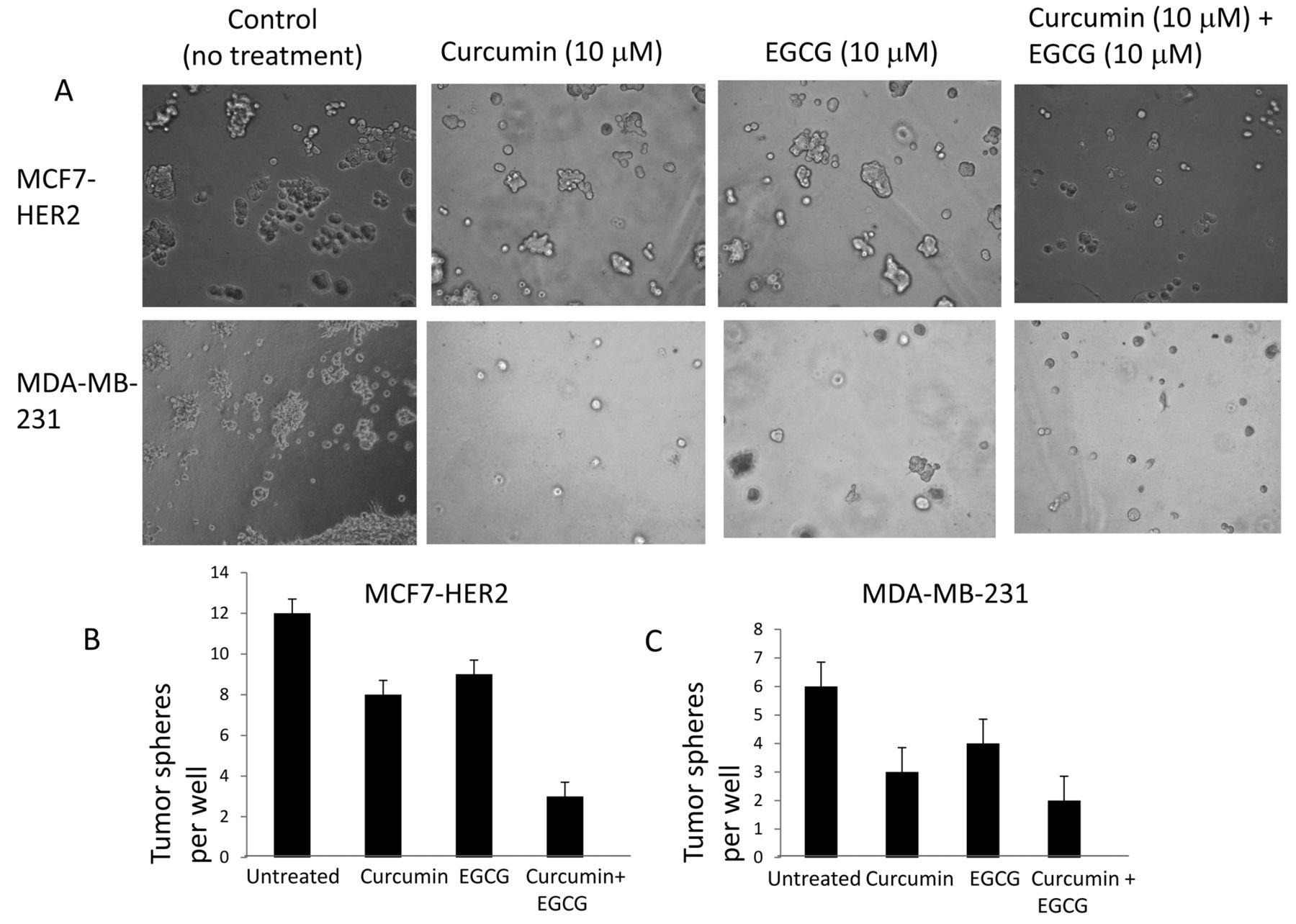
Tumor-sphere formation assays of MDA-MB-231 and MCF7-HER2 cell lines. A: MDA-MB-231 and MCF7-HER2 cell lines were seeded onto 24-well plates. Each well contained 2,000 cells. After the cells settled, they were treated with curcumin (10 μM) alone, EGCG (10 μM) alone and curcumin and EGCG in combination. Tumor-sphere formation was observed and counted for each well. B: Quantitative tumor-sphere formation by MDA-MB-231 and MCF7-HER2 cells under curcumin and EGCG treatments. Each experimentation was triplicated and error bars show the statistically significant differences between each group.
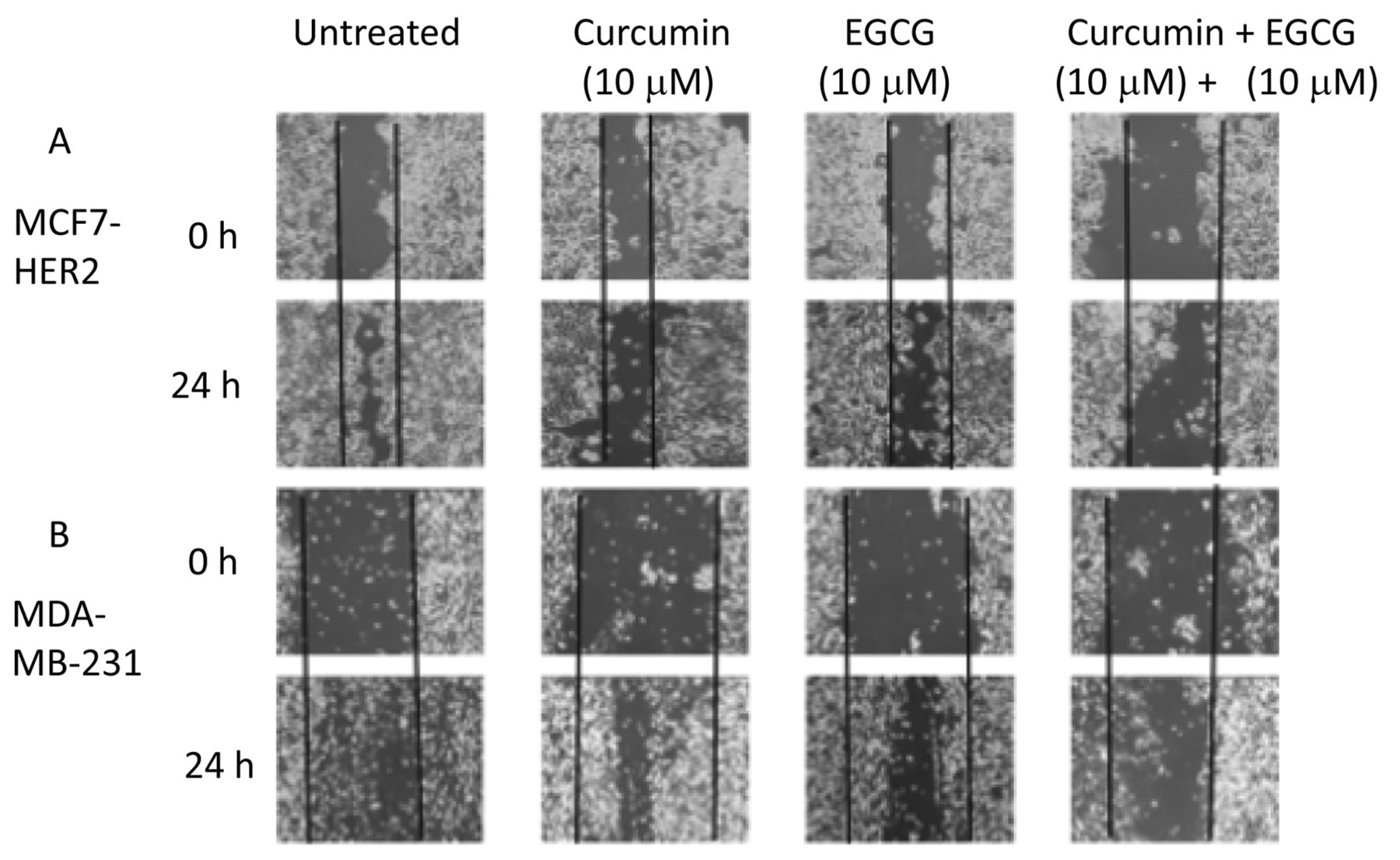
Wound-healing assay revealed that curcumin and EGCG suppress cell invasiveness. In agreement with the tumor sphere data, combined treatment with curcumin and EGCG suppressed the wound-healing ability of both MCF7-HER and MDA-MB-231 cells (Figure 2). Single treatment with either curcumin or EGCG also exerted inhibitory effects, however, combined treatment showed a synergistic effect (Figure 2). These data suggest that curcumin and EGCG possess cell invasion-suppressing capabilities. This is consistent with the tumor sphere formation data.
Combined treatment of curcumin and EGCG decreased the CD44-expressing subpopulation. CD44 has been recognized as one of the key cell surface biomarkers for tumor-initiating cells in breast cancer. Hence, we tested whether curcumin and EGCG reduce the expression of the CSC marker CD44 in breast cancer cells. MDA-MB-231 cells, known to express CD44, were treated with curcumin and EGCG either as single agents or in combination for 24 h and the CD44-positive sub-population was monitored by FACS profiling. As shown in Figure 3, following curcumin treatment, the CD44-positive cell population decreased from 99.05% to 98.31% (Figure 3A and B; R2 areas). Likewise, when MDA-MB-231 cells were treated with EGCG alone, the CD44-positive sub-population decreased to 98.89% (Figure 3C; R2 area). Finally, when curcumin and EGCG were used in combination, the CD44 (+) sub-population was decreased to 97.91% (Figure 3D; R2 area). These results may reflect an early stage of conversion of CSCs to non-stem cells.

Wound-healing assays of MDA-MB-231 and MCF7-HER2 cell lines. MDA-MB-231 (A) and MCF7-HER2 (B) cells were treated with either curcumin (10 μM) or EGCG (10 μM) or both. Wound-healing abilities were observed for 24 h.
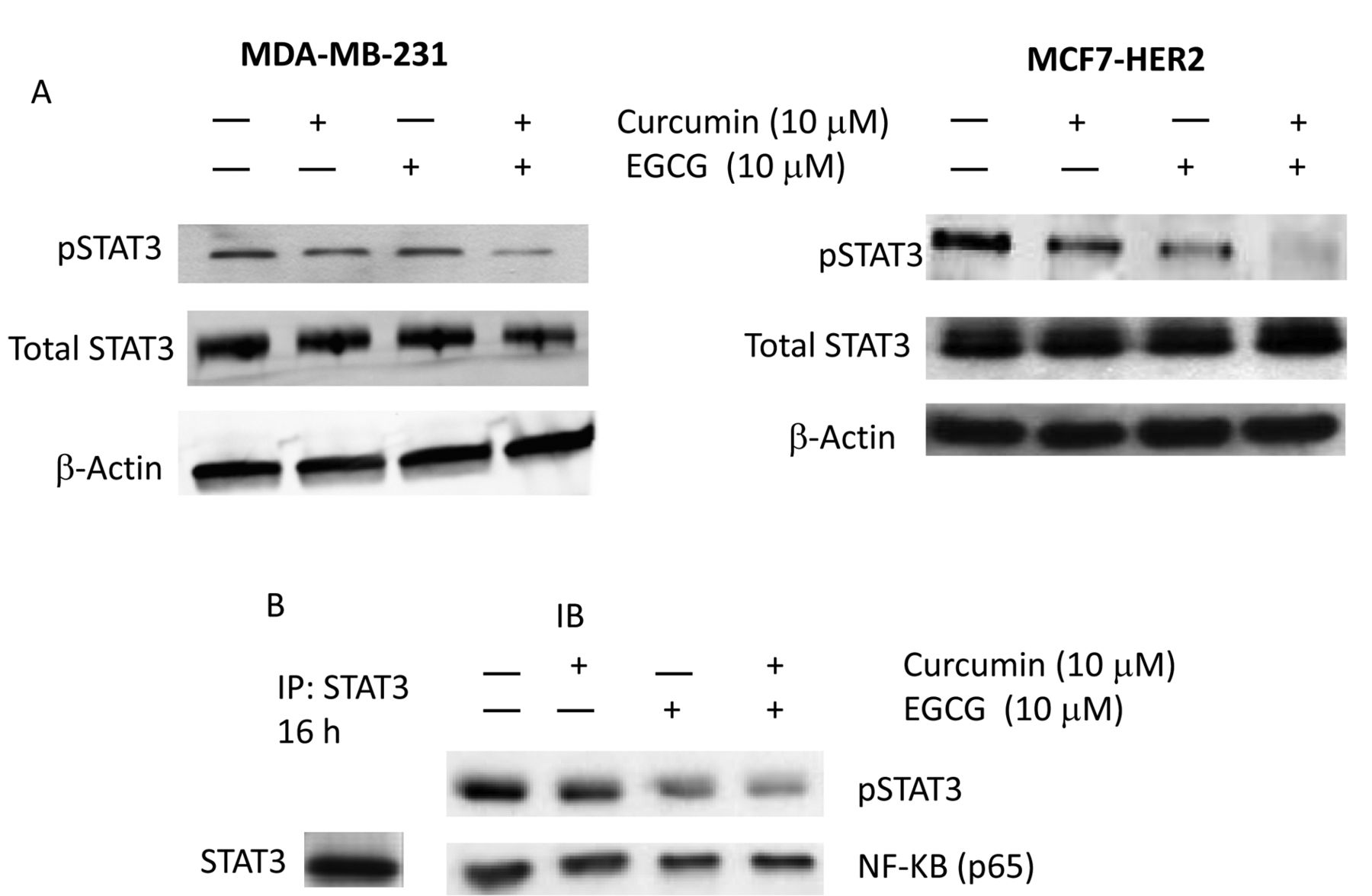
Curcumin and EGCG treatment blocked STAT3 phosphorylation step while STAT3-NFkB interaction was retained. Since we observed reduced tumorigenecity and cell invasiveness upon treatment with curcumin and EGCG, we wanted to study the molecular mechanisms that drive the CSC phenotype inhibition process in human breast cancer cells. Recently, it was demonstrated that CD44-positive breast cancer stem cells had preferential activation of STAT3, suggesting STAT3 as a potential therapeutic target in human breast tumors (26). Based on previous studies and our data, we decided to monitor the STAT3 activation status in breast cancer cells. We performed western blot analysis for pSTAT3 and total STAT3 in MDA-MB-231 and MCF7-HER2 cells treated with curcumin and EGCG. As shown in Figure 4A, when cells were treated with curcumin or EGCG, the pSTAT3 expression level clearly decreased in both MDA-MB-231 and MCF7-HER2 cells. The inhibition was further enhanced with the combined treatment of curcumin and EGCG.
NFκB transcription factor is constitutively activated in many types of human cancers (27). NFκB blocks apoptosis by stimulating anti-apoptotic genes and by suppressing apoptosis-inducing genes (28). Therefore we wanted to determine whether the treatment reduced STAT3 interaction with NFκB. STAT3 immuno-precipitation has revealed the protein-protein interaction between STAT3 and NFκB in MDA-MB-231 cells (Figure 4B). The interactions between STAT3 and NFkB proteins were mostly retained upon curcumin and EGCG treatments, however, the STAT3-interacting NFkB protein amount was modestly reduced. STAT3 protein still interacted with NFkB while phosphorylation was clearly inhibited with the curcumin and EGCG treatment. Our data suggest that curcumin and EGCG treatments block the STAT3 phosphorylation step, down-regulate the target genes, hence inhibit the cancer stem cell phenotype in human breast cancer cells.
Discussion
In the present this study, we selected two breast cancer cell lines, MDA-MB-231 and MCF7-HER2, to test the potential effects of curcumin and EGCG on CSCs. MDA-MB-231 is a triple-negative, i.e. ER−/PR−/HER2−, aggressive breast cancer cell line with a high proportion of CD44+ population. MCF7-HER2 is an ER+/HER2+ subtype with a high CD44+ cell population. As both cell lines possess a high CD44+ cell population, they serve as a good model for breast CSCs. Herein, we investigated the tumor-suppressive effects of curcumin and EGCG against breast CSCs. We employed the tumor-sphere formation culture and wound-healing assay to characterize the cancer stem cells. In tumor sphere culture, cells were cultured without attachment to the plates and examined for in vitro tumorigenecity. Wound-healing assay was employed to monitor cell invasiveness. When the cell lines were treated with curcumin (10 μM) and EGCG (10 μM) for 48 h, there was a significant reduction in tumorigenecity and cell invasiveness.

Flow cytometry profiling for the CD44+ sub-population. CD44+-expressing MDA-MB-231 cells were untreated (A), treated with curcumin (10 μM) (B), treated with EGCG (10 μM) (C) and treated with both curcumin and EGCG (each at 10 μM) for 48 h, then subjected to FACS profiling for the CD44+ cellular sub-population (R2 areas of each profile).
Increasing evidence suggests that STAT3 is constitutively activated in a variety of human malignancies, including breast tumors. The STAT3 signal pathway has been linked to cancer, and it activates critical target genes regulating cell proliferation and survival. Aberrant STAT3 activation promotes uncontrolled tumor cell growth and survival through multiple mechanisms, including increased expression of oncogenes, such as c-MYC (Myelocytomatosis oncogene), S-phase kinase-associated protein 2 (SKP2), and cyclin D1, as well as anti-apoptotic proteins, including BCL2, BCL-xL, myeloid leukemia cell differentiation protein (MCL1), and SURVIVIN (29-32). Notably, STAT3 has been shown to regulate growth and self-renewal of glioblastoma stem cells (33). Therefore, interfering with STAT3 oncogenic pathways may provide a novel targeted-therapy based on the capacity of STAT3 to maintain the CSC population. Curcumin and EGCG treatment clearly inhibited STAT3 activation in our model system, suggesting these two phytochemicals may be good candidates for STAT3 targeting.

STAT3 phosphorylation was blocked by curcumin and EGCG treatment in a synergistic manner. A: Western blot analyses of pSTAT3 and total STAT3 when MDA-MB-231 and MCF7-HER2 cells were treated with curcumin (10 μM) and EGCG (10 μM). STAT3 phosphorylation was clearly inhibited in both cell lines. B: Pull-down assay with STAT3 monoclonal antibody. Whole cell lysates were subjected to immunoprecipitation with STAT3 antibody. The immuno-pellets were collected, washed and run on immune-blotting gels. STAT3 protein was bound to NFκB in the MDA-MB-231 cell line. When cells were treated with curcumin and EGCG, the STAT3-NFkB interaction was retained.
This report also examined p65, a subunit of the NFκB transcription factor, in these breast cancer cell lines. The immunoprecipitation study revealed the protein-protein interactions between STAT3 and NFkB in human breast cancer cells. When STAT3 phosphorylation was inhibited, the STAT3-NFkB interaction was still retained, yet the STAT3-interacting NFkB protein amount was slightly decreased. This is possibly due to the decreased interaction between STAT3-NFkB in the nucleus. STAT3 can translocate into the nucleus only when it is phosphorylated. Since curcumin and EGCG blocked the phosphorylation step, STAT3 could not translocate into the nucleus, hence could not interact with NFkB there. Most STAT3-NFkB interactions take place in the cytoplasm in this case. It remains to be seen whether there is a differential interaction mode between cytoplasmic and nucleus STAT3-NFkB proteins upon curcumin and EGCG treatment.
The CD44+ cell population was also decreased following curcumin and EGCG treatments. Our results suggest that the final destination of STAT3 and NFκB signaling may be the CD44 expression and accompanied by a cancer stem cell phenotype. Multiple studies have demonstrated that invasion and metastasis are mediated by a cellular component that displays high CD44 activity, suggesting that CD44 may be a significant target for cancer therapy (34, 35). CD44, a transmembrane glycoprotein, is a major receptor for extracellular proteins and has been recognized as one of the key cell surface biomarkers for tumor-initiating cells in breast cancer. The present work suggests that STAT3 signaling-mediated CD44 can be a good cellular target for human breast cancer.
The specific mechanism causing the reduction in CSC population by curcumin and EGCG was explored in this study. Curcumin and EGCG selectively block STAT3 phosphorylation, hence inhibiting STAT3 translocation into the nucleus. Interaction between STAT3 and NFkB was weakened in the nucleus due to the lack of pSTAT3. As a result, the expression levels of STAT3-NFkB target genes were decreased. The decreased expression levels of these target genes down-regulated the CD44 expression. Taken together from in vitro studies so far, that curcumin and EGCG seem to reduce reduced the stem cell properties of the breast cancer cells by modulating STAT3–NFκB signaling pathways. We are now using in vivo models to study the role of STAT3, NFκB and CD44 in tumorigenesis.
Acknowledgements
We thank the members of the Division of Cancer Research and Training at Charles R. Drew University for helpful suggestions. This work was supported by grants from NIH/National Cancer Institute 1U54 CA14393 and NIMHD U54 MD007598 to J.V. Vadgama and NIH/NIMHD U54 MD007598 AXIS pilot grant awarded to S.S. Chung. S. Steven Chung is a scholar supported by the Clinical Research Education and Career Development by the NIMHD under award numbers 5MD007610.
Footnotes
-
This article is freely accessible online.
- Received September 26, 2014.
- Revision received October 22, 2014.
- Accepted October 27, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.