


Abstract
Marine invertebrates called ascidians are prolific producers of bioactive substances. The ascidian Eudistoma viride, distributed along the Southeast coast of India, was investigated for its in vitro cytotoxic activity against human cervical carcinoma (HeLa) cells by the MTT assay. The crude methanolic extract of E. viride, with an IC50 of 53 μg/ml, was dose-dependently cytotoxic. It was more potent at 100 μg/ml than cyclohexamide (1 μg/ml), reducing cell viability to 9.2%. Among nine fractions separated by chromatography, ECF-8 exhibited prominent cytoxic activity at 10 μg/ml. The HPLC fraction EHF-21 of ECF-8 was remarkably dose- and time-dependently cytotoxic, with 39.8% viable cells at 1 μg/ml compared to 51% in cyclohexamide-treated cells at the same concentration; the IC50 was 0.49 μg/ml. Hoechst staining of HeLa cells treated with EHF-21 at 0.5 μg/ml revealed apoptotic events such an cell shrinkage, membrane blebbing, chromatin condensation and formation of apoptotic bodies. Cell size and granularity study showed changes in light scatter, indicating the characteristic feature of cells dying by apoptosis. The cell-cycle analysis of HeLa cells treated with fraction EHF-21 at 1 μg/ml showed the marked arrest of cells in G0/G1, S and G2/M phases and an increase in the sub G0/G1 population indicated an increase in the apoptotic cell population. The statistical analysis of the sub-G1 region showed a dose-dependent induction of apoptosis. DNA fragmentation was also observed in HeLa cells treated with EHF-21. The active EHF-21 fraction, a brominated indole alkaloid Eudistomin H, led to apoptotic death of HeLa cells.
Abbreviations: IC50: Inhibition Concentration 50; HPLC: High-performance liquid chromatography; MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; DMSO: Dimethyl Sulfoxide; FSC/SSC: Forward scatter/Side scatter; MALDI-TOF: Matrix-assisted laser desorption/ionization-time of flight, FTIR: Fourier transform infrared spectroscopy; TMS: Tetramethylsilane; CDCL3: Chloroform-d; ESI-MS: Electrospray Ionisation Mass Spectrometry, US-NCI: United States-National Cancer Institute; ED50: Effective Dose 50, H: Hexane; DCM: Dichloromethane; M or Meth: Methanol; EHF: Eudistoma HPLC fraction; ECF: Eudistoma Column fraction.
Treatments for cancer, such as radiation and chemotherapy, have significant side-effects. Hence, less toxic treatments with significantly fewer side-effects have been the main objective of anticancer research. Significantly, the comprehensive understanding of tumor biology and molecular mechanisms have led to new insights into the mechanisms responsible for the disease, modifying the general approach towards cancer treatment.
Apoptosis, a strongly regulated and structurally-distinct form of programmed cell death, has widespread biological significance, hence apoptosis has been recognized as an important target for therapeutic intervention and rational drug discovery (1) and cancer treatment strategies are directed towards reconstituting the tumor cell's ability to undergo apoptosis (2).
Marine natural products have received much attention due to their chemical novelty and great diversity (3). Sponges, bryozoans and tunicates are among the most promising sources of dynamic principles for drug development (4).
Ascidians are prolific producers of bioactive substances and many ascidian compounds have been shown to possess cytotoxic properties. Metabolites such as halocynthiaxanthin and fucoxanthinol from Halocynthia roretzi (5), meridianins, brominated 3-(2-aminopyrimidine)-indoles from Aplidium meridianum (6), and compounds from Diplosoma virens (7) and Policlinum indicum (8), have novel modes of action of inducing apoptosis and cell-cycle arrest. Among the ascidian families of Didemnidae and Polycitoridae, the most prolific producers of biologically-active compounds (9), the genus Eudistoma, belonging to the most diverse family Polycitoridae and inhabiting mainly tropical regions (10), has been the source of several cytotoxic alkaloids (11), including eudistomins, eilatin, staurosporine derivatives, methyleudistomins and pibocin (12, 13). With this in mind, we investigated the cytotoxic potential of compounds from the ascidian Eudistoma viride, distributed along the Southeast coast of India, and their possible mechanisms of action against HeLa cell lines.
Materials and Methods
Extraction of ascidian. Eudistoma viride Tokioka, 1955 (Chordata, Urochordata, Ascidiacea, Enterogona, Polycitoridae), collected from the Tuticorin coast (Lat 8°45’N and Long 78°13’E) of the Southeast coast of India. The ascidian was identified using standard keys (14) and the identity was confirmed by Dr. V.K. Meenakshi of APC Mahalakshmi College, Tuticorin. The ascidians were brought to the laboratory, washed thoroughly with sterile distilled water, cut into small pieces, rinsed with sterile distilled water and air-dried for 24 h at room temperature before extraction with methanol. Methanol was chosen based on preliminary antibacterial activity screening against human bacterial pathogens with hexane, chloroform, diethyl ether, ethyl acetate, acetone and methanol (data not presented). The crude extract, cold steeped overnight at −18°C and filtered with Whatman No.1 filter paper, was evaporated to dryness in a rotary evaporator (8,15), and used for the preliminary cytotoxic screening against cervical cancer cells (HeLa).
Tumor cell culture. The human cervical carcinoma (HeLa) cell line from the American Type Culture Collection (ATCC, Manassas, VA 20108 USA), obtained from the National Center for Biological Sciences, Bangalore, India, were maintained under a cell density of 1×106/ml (to avoid mutations of the cells) in tissue culture flasks with F-12 Dulbecco's modified Eagle's medium (Sigma, Bangalore, India) (17) supplemented with 500 ml fetal bovine serum (Sigma), penicillin (100 U/ml) and streptomycin (100 μg/ml) in a CO2 incubator. Cells were passaged three times a week and cells cultivated longer than 20 passages (8,18) were not used for the experiments.
In vitro cytotoxicity assay. The MTT assay was followed based on Martinez-Garcia et al (19). HeLa cells (9×105 cells/ml in a volume of 100 μl/well) (20) were incubated with methanol crude extract of E. viride at concentrations of 25, 50, 75 and 100 μg/ml in a 96-well microplates and incubated in an incubator with 5% CO2 at 37°C for 24 h. Then 100 μl of MTT solution [5 mg/ml in phosphate-buffered saline (PBS) (Sigma)] was added to each well and cells were incubated for 2 h at 37°C under the same conditions. The accumulated formazan crystals were dissolved with the addition of 150 μl DMSO to each well. The culture plate was then gently shaken in the dark at room temperature for 1 h and the absorbance was measured at a wavelength of 570 nm with an ELISA plate reader (BIORAD model 680, Life Science, Guragon, India). Cyclohexamide at 1 μg/ml was used as positive control and a negative control without ascidian extract was maintained separately. The experiment was carried out in triplicate. Survival of the cells in treated wells was measured as the percentage of absorbance compared to that of the control wells (non-treated cells, taken as 100% survival). The IC50 value (effective concentration required to inhibit cell growth by 50%) was calculated from dose–response curves (20).
Chromatographic isolation of cytotoxic compound. Column fractionation: The crude methanolic extract was fractionated using reverse-phase silica gel flash column chromatography (230-400 mesh; LOBA Chemie, Bangalore, India) (16) with a 43×2 cm size column. Based on gradient partitioning (16) and thin layer chromatography (TLC) [pre-coated TLC plates with silica gel, 0.2 mm thickness, 20×20 cm (Merck, Bangalore, India) silica gel] of crude extract, a step-gradient solvent system of increasing polarity from hexane (Hex) to dichloromethane (DCM) to methanol (Meth) (100% Hex, 75% Hex:25% DCM, 50% Hex:50% DCM, 25% Hex:75% DCM, 100% DCM, 75% DCM:25% Meth, 50% DCM:50% Meth, 25% DCM:75% Meth, 100% Meth) was used to fractionate the crude extract. The purity of the fractions was assessed with TLC and the cytotoxicity of the nine eluted column fractions (ECFs) was determined through MTT assay against HeLa cell lines at a concentration of 10 μg/ml, selected at random based on the crude activity profile.
High performance liquid chromatography (HPLC): Fraction ECF-8 (25% DCM:75% Meth), which showed potent cytotoxicity against HeLa cell line, was collected, pooled, concentrated and further purified using semipreparative HPLC. An isocratic solvent system using methanol/water (0.05% trifluoroacetic acid) (10:90) was employed to separate the mixture of compounds (21). The 23 fractions (EHFs) which exhibited prominent peaks during elution in semipreparative HPLC column fractionation were collected and subjected to MTT assay at 2 μg/ml concentration. The fractions thus obtained were spotted onto TLC plates and observed under UV light. Pancal-D was used as a spray reagent (21.6 g ammonium molybdate, 1.6 g ceric sulphate and 50 ml concentrated H2SO4 were added under ice-cold conditions and then made up to 500 ml with water). The biologically active TLC fraction EHF-21 was further used for structural elucidation.
Dose- and time-dependent cytotoxicity of fraction EHF-21. The dose-dependent (0.25 to 1 μg/ml) activity of fraction EHF-21 over a 24-h period and subsequently, the time-dependent (at 2, 4, 8, 16 and 24 h) activity of 0.50 μg/ml over a 24-h period were assessed. Controls were maintained separately using cycloheximide (1 μg/ml) as positive control. The MTT assay was carried out to determine the viability of treated and untreated cells.
Hoechst staining. The E. viride fraction EHF-21 was added at a concentration of 0.5 μg/ml to a 24-well tissue culture plates with HeLa cells (7×105 cells/ml; 1 ml) (18) and observed for different periods of time (4, 8, 16, 24 h). Untreated HeLa cells were maintained as control. The cells were harvested by centrifugation (110 × g; 10 min; 4°C) at appropriate time periods, 1 ml PBS was used to resuspend the pellets and cells were stained with 10 μl Hoechst solution (dissolved in double distilled water at a stock concentration of 0.1 mg/ml) for 5 min at 37°C in the dark. The dye was removed by centrifugation (110 × g; 10 min; 4°C) and by washing with PBS. The condensed chromatin was visualized at 200-fold magnification with a Nikon EclipseTE 2000-S fluorescence microscope (Nikon Corporation, Kanagawa, Japan) using a filter 02 (λ ex: 365 nm; λ em: 420 nm).
Cell-cycle analysis and quantification of apoptosis. In cell-cycle analysis (22, 23), HeLa cells at 7×105 cells/ml, (18) seeded in a 24-well tissue culture plate (1 ml/well), were treated with E. viride fraction EHF-21 at 0.25, 0.50, 0.75 and 1 μg/ml for 24 h. Negative and positive (1 μg/ml cycloheximide) controls were maintained separately. After incubation, the cells were harvested by centrifugation (600 ×g, 10 min, 4°C), washed with cold PBS and resuspended in 500 μl hypotonic propidium iodide (PI) (Sigma) staining solution. The tubes were stored at 4°C in the dark overnight. The cell fluorescence (FL-2) and scatter characteristics (FSC/SSC) were acquired (23) with a FACS calibur instrument (Becton Dickinson, Heidelberg, Germany), equipped with a 488 nm argon-ion laser and analyzed using LYSYS II software (18). The analysis, depicted in a histogram with the number of cells (counts) plotted against the relative fluorescence intensity of PI (FL-2; λ em: 585 nm; red fluorescence), reflected the cell-cycle distribution of the cell population. Untreated control cells were used for the adjustment of instrument settings. Marked regions of sub-G1 cells were evaluated to quantify the percentage of apoptotic cells (18).
Determination of cell size and granularity. HeLa cells (7×105 cells/ml) (18), seeded in a 24-well tissue culture plate (1 ml/well), were left 24 h after treating with E. viride fraction EHF-21 at a moderately cytoxic concentration of 0.5 μg/ml. Untreated HeLa cells were maintained separately as control. The cells were then harvested by centrifugation (600 ×g, 10 min, 4°C), washed with cold PBS, resuspended in 500 μl of cold PBS and analyzed by flow cytometry. Untreated control cells were used for adjusting cytometer settings (FSC, SSC). The detected signals were illustrated in dot plots (FSC versus SSC). Normal and apoptotic cells were differentiated by assessing the distribution of the dots within the plot (24).
DNA fragmentation assay. E. viride fraction EHF-21 at concentrations of 0.25, 0.50 and 1 μg/ ml were added to 24-well tissue culture plates with HeLa cells and incubated for 24 h to assess the internucleosomal cleavage of DNA (25). Then the cells were washed twice with ice-cold PBS, resuspended in lysis buffer containing 10 mM Tris–HCl (pH 8.0), 20 mM EDTA and 0.5% Triton X-100) (Sigma) and incubated on ice for 30 minutes. Then the suspension was centrifuged at 8400rcf xg for 10 min at 4°C and DNA was extracted with phenol–chloroform, precipitated with 0.1 vol 3 M sodium acetate and 2.5 vol ethanol and stored at −20°C overnight. DNA was pelleted by centrifugation at 8400rcf for 5 min at 4°C, rinsed with 70% ethanol and then resuspended in TRIS-EDTA buffer (pH 8.0) (Sigma) containing, 30 μg/ml of RNase (Sigma) and incubated for 6 h at 37°C. DNA was run on a 2% agarose gel with ethidium bromide staining (26). After electrophoresis, DNA was visualized under UV and documented.
Characterization of the active compound. The mass spectrum for the cytotoxic fraction EHF-21 was obtained in ultraflex MALDI-TOF (Bruker Daltonics, Germany). Infrared spectrum for functional group analysis of Eudistomin H was recorded by using a Vector 22 Brucker FTIR instrument. Solid samples were recorded as KBr pellets and liquid samples were taken as solution spectra in CHCl3 (27). 1H and 13C nuclear magnetic resonance (NMR) spectra were recorded on a BRUKER DRX 500 solid state NMR spectrometer with proton frequency of 300.013 MHz and probe of 5 mm QNP (1H, 13C, 19F, 31P) instrument using TMS as the internal standard (28). CDCl3 was used as the solvent. By using the generated data, the molecular formula and structural characteristics of the active compound were elucidated.

Dose-dependent cytotoxicity of crude methanolic extract of Eudistoma viride against HeLa cells (mean±SD).
Statistical analysis. The results were expressed as the mean±SD of three replicates. Student's t-test indicated the significance of the experimental results, compared to the control and a value of p<0.05 was considered as significant.
Results
The crude methanolic extract of E. viride exerted dose-dependent cytotoxicity against HeLa cells (Figure 1) and at 100 μg/ml, it was more potent than cyclohexamide (1 μg/ml), causing low cell viability of 9.2%. The IC50 was 53 μg/ml. Among the nine column fractions, the cytotoxic activity was low in fractions ECF-1 (100% Hex) and ECF-2 (75% Hex:25%DCM) with higher hexane content (Figure 2). ECF-4 (25% Hex:75% DCM) also exhibited low activity. Comparatively, fractions ECF-3 (50% Hex:50% DCM) and ECF-5 (100% DCM) had higher cytotoxicity. The fractions with higher methanolic content had higher cytotoxicity, with fraction ECF-8 (25% D CM: 75% Meth) showing prominence. However, the TLC of the active fraction ECF-8 revealed many compounds with different retention time Rf values (results not presented).
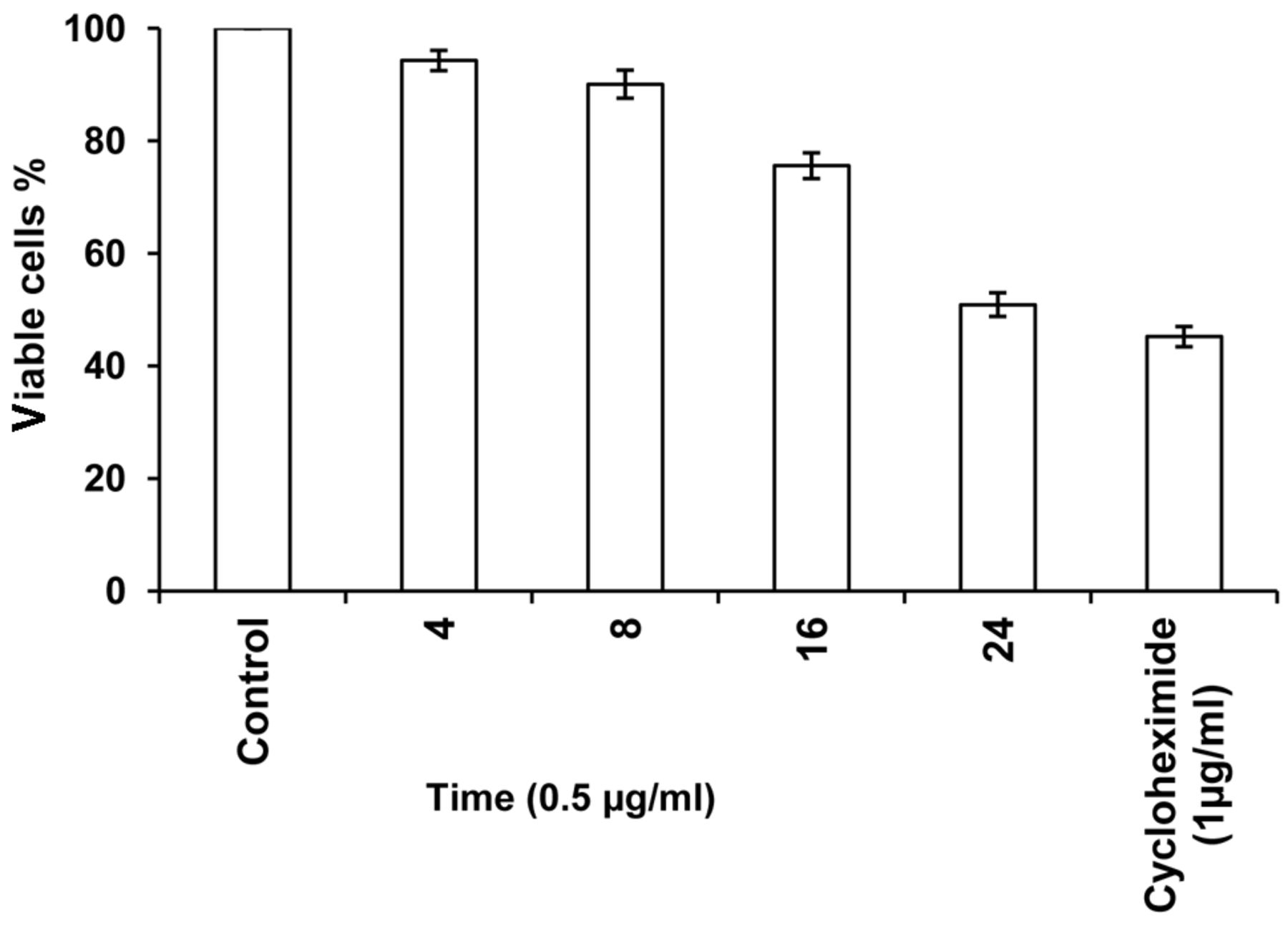
In the subsequent semipreparative HPLC of ECF-8, 23 fractions (EHF-1 to EHF-23) with prominent peaks with good retention time were collected and among them, 21 fractions showed more than 50% inhibition of cell proliferation at 2 μg/ml concentration (Figure 3). HPLC fractions EHF-12 to 21 and 22 had prominent cytotoxicity, with fractions EHF-20 and EHF-21 showing potent activity. EHF-21 showed remarkable dose- and time-dependent cytotoxicity against HeLa cells (Figures 4 and 5). At 1 μg/ml concentration, the percentage of viable cyclohexamide-treated cells was 51% and that for cells treated with EHF-21 was 39.8%; 0.5 and 0.75 μg/ml EHF-21 led to 48.6% and 44.2% viable cells, respectively. lower than that for cells treated with cyclohexamide at 1 μg/ml. The activity of EHF-21 at 0.5 μg/ml increased with increasing incubation time. The IC50 was 0.49 μg/ml.

Cytotoxicity of fractions eluted by column chromatography of methanolic extract of Eudistoma viride against HeLa cells (mean±SD). H: DCM: M.
Hoechst staining of cells treated with fraction EHF-21 and untreated cells displayed differential staining due to the highly condensed chromatin in treated cells and intact normal chromosomes in untreated cells. The apoptotic process in the cells treated with EHF-21 at 0.50 μg/ml showed sequential events from 4 to 24 h period, portraying the apoptotic events in drug-induced apoptosis. This was clearly indicated the characteristic morphological and biochemical features of apoptosis such as cell shrinkage, membrane blebbing, chromatin condensation and formation of apoptotic bodies in HeLa cells. These processes started from 4 h and blebbing appeared from 8 h, leading to formation of apoptotic bodies (Figure 6). The present study showed changes in light scatter i.e. a decrease in FSC as a sign of lower cell size and an increase in side scatter as a sign of granularity compared to control cells, indicating the characteristic feature of cells dying of apoptosis (Figure 7). The occurrence of a second population of apoptotic cells in the range of lower FSC and higher SSC compared to control cells was noticed.
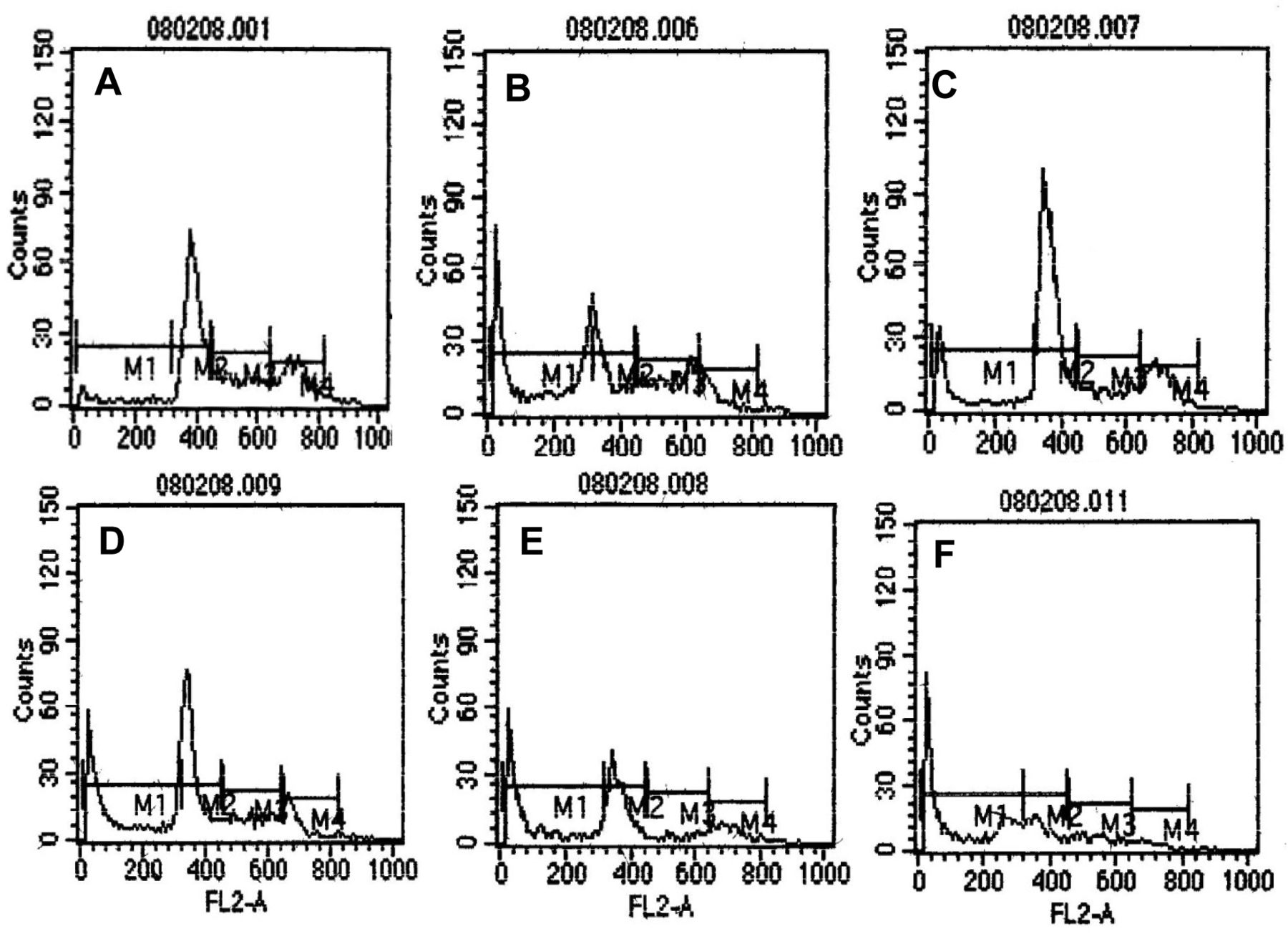
In the cell-cycle profiles of HeLa cells, determined by flow cytometry after staining of the DNA with PI, the positive control of cycloheximide-treated HeLa cells showed an increase in the sub G0/G1 population, targeting cells in G0/G1 and G2/M phases (Figure 8). Treatment of HeLa cells with EHF-21 at 0.25 μg/ml led to a substantial elevation in the sub G0/G1 population, indicating the induction of apoptosis. The G0/G1 population continued to increase with increasing concentration of fraction EHF-21. The arrest was marked in the G2/M phase on treatment with 0.50 μg/ml EHF-21, which also Ied to marked arrest of cells in G0/G1 and a further increase in the sub G0/G1 population. EHF-21 at 0.75 μg/ml led to the marked arrest of cells in S and G2/M phases and substantial arrest of the G0/G1 population. At 1 μg/ml of EHF-21, marked arrest of cells in G0/G1, S and G2/M phases and an elevated sub-G0/G1 population indicated an increase in the apoptotic cell population. The statistical analysis of the sub-G1 region showed dose-dependent apoptosis induction by fraction EHF-21 (Figure 9). At the highest concentration tested (1 μg/ml), the effect of EHF-21 was almost equally potent as the positive control cycloheximide at inducing apoptosis in the HeLa cells. After 24 h, the percentage of apoptotic cells at all concentrations increased, with an increase of 65.21% for 1 μg/ml.
In agarose gel electrophoresis of the chromosomal DNA isolated from HeLa cells treated with fraction EHF-21 (Figure 10), the study clearly established the DNA damage caused by ascidian E. viride fraction EHF-21.
The characterization of fraction EHF-21 resulted in the isolation of Eudistomin H as a yellow powder, 6-bromo-1-(3,4-dihydro-2H-pyrrol-5-yl)-9H-β-carboline, with a molecular formula of C15H12BrN3, established on the basis of interpretation of the ESI-MS, FT-IR spectroscopy and NMR data. The molecular weight was 314 Da. A strong peak at 1017.27 cm−1 in the infra-red spectrum clearly illustrated the presence of halogen (bromine). The structure of Eudistomin H is presented in Figure 11.
Discussion
The cytotoxic activity of the extract of E. viride was similar to that of E. vannamei and Eudistoma sp. from Northeast Brazil (29). However, the IC50 of 53 μg/ml for the crude methanolic extract was comparatively higher than that reported for E. vannamei (<2 μg/ml for CEM, 11.2 μg/ml for HL-60, 23.8 μg/ml for B16 and 14.3 μg/ml for HCT-8 cell lines) (29); Ecteinascidin-743 from Caribbean tunicate Ecteinascidia turbinata (0.5 ng/ml against L1210 leukemia cells) (30, 31) and Lamellarin b from Didemnum sp. (4.8 μg/ml against HL-60) (32). However, the activity was much greater than that of pyrrolopyrimidine alkaloid, rigidin E, rigidin, and 1-methylherbipoline, isolated from a Papua New Guinea tunicate Eudistoma sp., which led only to minimal growth inhibition at 100 μg/ml against HCT 116 human colon carcinoma cell lines (33). Nevertheless, the IC50 of the crude extract was higher than the limit prescribed by the US-NCI by which an extract is recognized as active by an IC50 of <20 μg/ml (34).
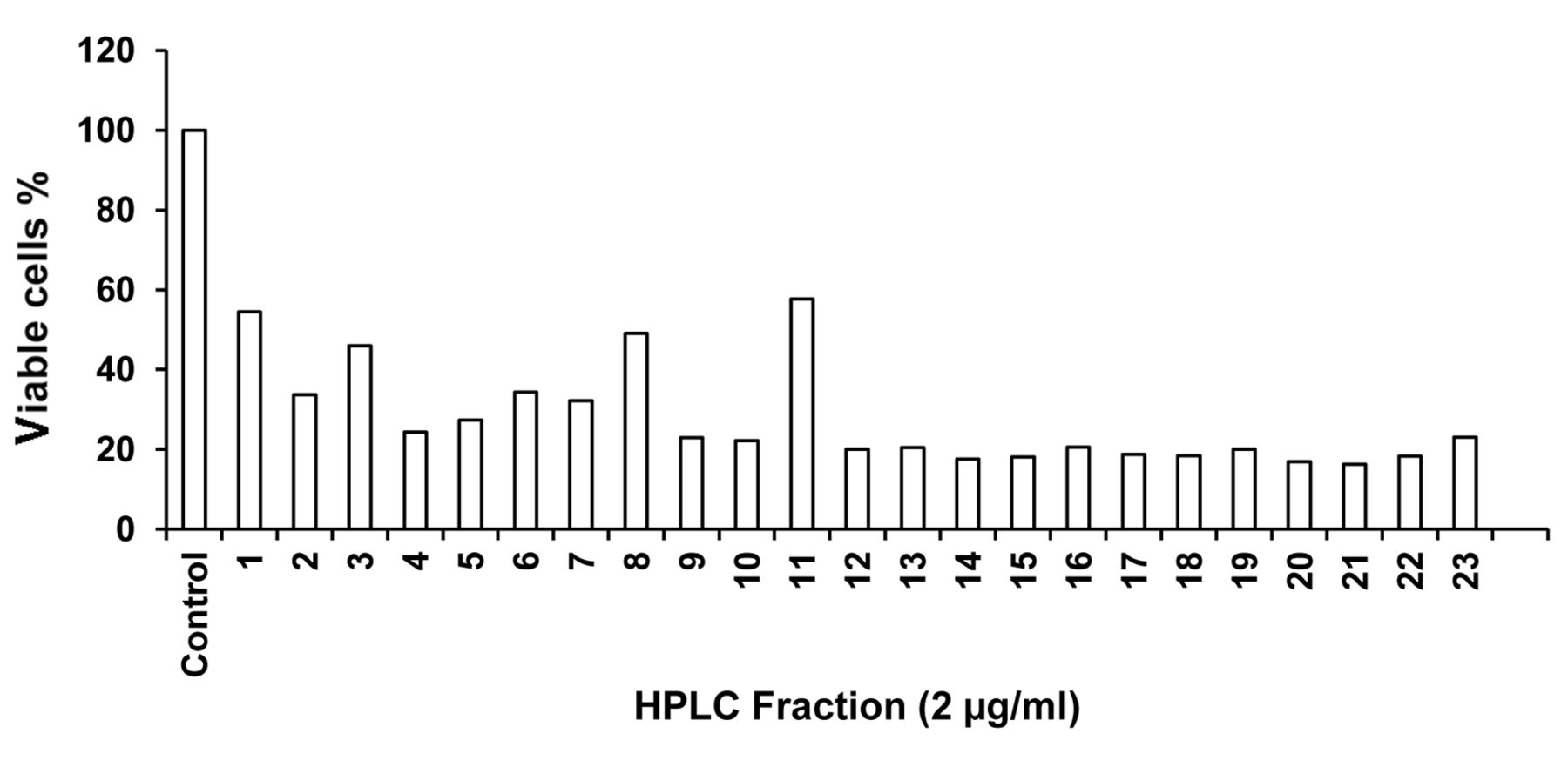
Cytotoxicity of HPLC fraction of methanolic extract of Eudistoma viride against HeLa cells.

Concentration-dependent cytotoxicity of fraction EHF-21 against HeLa cells (mean±SD).
The cytotoxicity of HPLC fractions against HeLa cell lines was comparatively higher than that reported for Hydroperoxysterol 1, from Eudistoma sp. against Hep3B (human hepatoma cells) and A549 (human lung adenocarcinoma) and that of hydroperoxysterol 2 towards Hep3B, HepG2 (human hepatocellular carcinoma), MECF7 (human breast carcinoma) and MDA-MB-231 (human breast carcinoma) cells (35). The observation of more than 50% inhibition of cell proliferation by almost 21 HPLC fractions at 2 μg/ml highlights the potential of E. viride as a source of effective cytotoxic agents. The IC50 value was higher than that of a range of 0.022-4.78 μM for the chlorinated didemnin classes of compounds from tunicate Trididemnum against SK-MEL (melanoma), KB (epidermal carcinoma), BT549 (breast carcinoma) and SK-OV-3 (ovarian carcinoma) cancer cell lines (36) and Tanjungide A (0.19 to 0.33 μM) and Tanjungide B (1.00 to 2.50 μM) from the tunicate Diazona cf formosa against lung carcinoma (A549), colorectal carcinoma (HT29) and breast adenocarcinoma (MDA-MB-231) cell lines (37). The IC50 value of 0.49 μg/ml for the EHF-21 fraction was very well below the 4 μg/ml prescribed by the US-NCI for purified compounds to be considered as active (34).

Time-dependent cytotoxicity of fraction EHF-21 (0.5 μg/ml) against HeLa cells (mean±SD).
It has been suggested that susceptibility to apoptosis-inducing effects of chemotherapeutic drugs may depend on the intrinsic ability of tumor cells to respond by apoptosis (38, 39). In the present study, the EHF-21 fraction was shown to induce apoptosis of HeLa cells in contrast to the induction of both necrosis and apoptosis by substances from Didemnum psammathodes (40). Apoptosis induction by compounds isolated from E. viride was found to be due to Ecteinascidin-743 (ET-743) from Ecteinascidia turbinata, which induced a massive apoptosis of Ewing's sarcoma cells (41). Aplidine, from the Mediterranean tunicate Aplidium albicans, at nanomolar concentrations (10-100 nM) induced apoptosis of human leukemic cell lines and primary leukemia cell cultures from patients with leukemia (42) and two compounds from the ascidian Diplosoma virens were cytotoxic against HCT116 cells (human colorectal cancer cells) by triggering apoptotic cell death (7).

Apotopsis induced in HeLa cells by Eudistoma viride fraction EHF-21 at 0.50 μg. A: Control; B: at 4 h, clear chromatin condensation (arrow); C: 8 h, cell shrinkage apparent (arrow); D: 16 h, visible membrane blebbing (arrow); E: 24 h, membrane blebbing and formation of apoptotic bodies (arrows).

HeLa cells treated with Eudistoma viride fraction EHF-21 also exhibited change in morphological features in flow cytometric analysis. A: Untreated HeLa cells exhibited uniform cell size and granularity. B: HeLa cells treated with EHF-21 exhibited a second population of cells which denoted the presence of apoptotic cells in the range of lower FSC and higher SSC when compared to untreated cells.
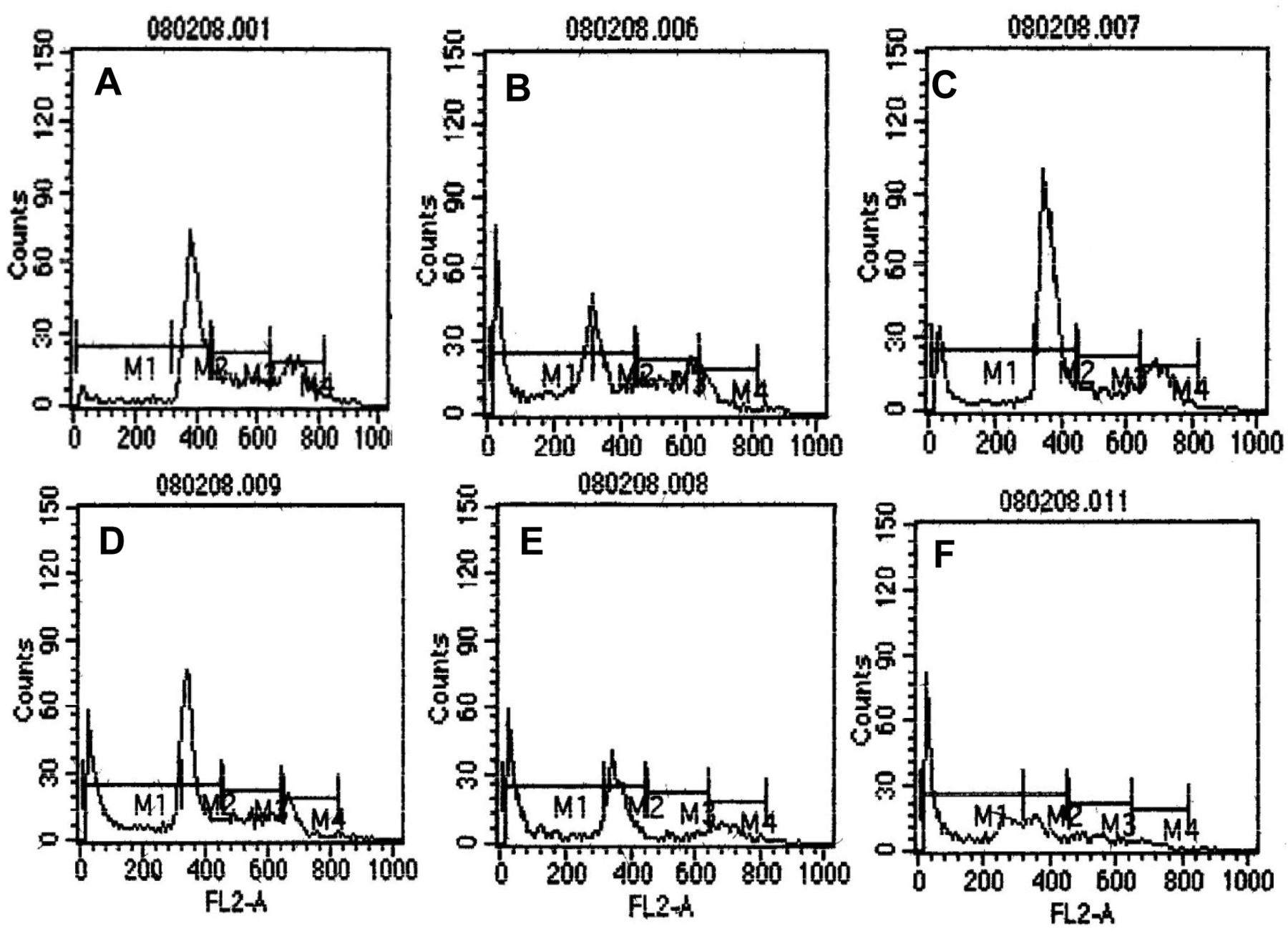
Cell-cycle analysis indicating the different phases of the cell cycle. A: Untreated control; B: positive control of HeLa cells treated with 1 μg/ml cycloheximide; C-F: HeLa cells treated with fraction EHF-21 at 0.25 μg/ml, 0.50 μg/m, 0.75 μg/ml and 1 μg/ml, respectively. The peak in sub-G0/G1 phase of the cell cycle indicates the presence of apoptotic cells. M1: Sub G0/G1, M2: G0/G1, M3: S, M4: G2/M.

Dose-dependent (0.25-1 μg) induction of apoptosis of HeLa cells by fraction EHF-21.
The appearance of two different cell populations in cells treated with EHF-21 when compared to untreated control cells in flow cytometric analysis indicated that the cells treated with fraction EHF-21 had undergone some cellular changes, either in surface or inner components, coinciding with the observation of production of two different cell populations in ascididemin-treated human leukemia Jurkat T-cells measured by flow cytometry (18). A marked arrest of HeLa cells in G2/M phase on treatment with 0.50 μg/ml of fraction EHF-21 and significant appearance of cells in the sub-G0/G1 phase indicated induction of apoptosis. The arrest of HeLa cells in the G2/M phase corroborates the observation that some anticancer drugs, when applied in vivo, are able to intercalate with DNA in cancer cells and do arrest cancer cells in the cell cycle via G2/M arrest (41, 43, 44). The present study was also in line with the observation that ascidian metabolites didemnin B and aplidine interfered with the synthesis of DNA and proteins and induced cell-cycle arrest (45).
The percentage of apoptotic cells increased gradually with increasing concentration of fraction EHF-21. The interference of fraction EHF-21 with the cell cycle at the G2/M phase and induction of apoptosis in HeLa cells in a dose-dependent manner correlated with the reported interference of paclitaxel with the cell cycle at the G2/M phase and induction of apoptosis of HL-60 cells in a dose- and time-dependant manner (46). The marked arrest of HeLa cells in the G2/M phase by EHF-21 parallels the observation that meridianin B induced proliferation arrest in exponentially growing cells, clearly accompanied by an accumulation in G0/G1 phase, whereas meridianin E treatment led to an arrest in the G2/M phase (6). The mode of marked arrest of cells in the G2/M phase concurs with findings with halichondrins, potent tubulin inhibitors, which noncompetitively bind to the Vinca binding site and cause a characteristic G2-M cell-cycle arrest with concomitant disruption of the mitotic spindle (47, 48). Similarly, Dolastatin 10, isolated from a sea hare, cause cell-cycle arrest in metaphase as it binds to the rhizoxin/maytansine binding site and to the exchangeable guanosine triphosphate site on tubulin (49). The time-dependent increase in activity of EHF-21 fraction in the present study was in line with the cytotoxicity of lissoclinolide, isolated from the marine ascidian Lissoclinum patella, which at 2.4 μM resulted in a strong arrest in the G2/M phase of the cell cycle of HCT 116 human colon tumor cells after 24 h exposure (50).
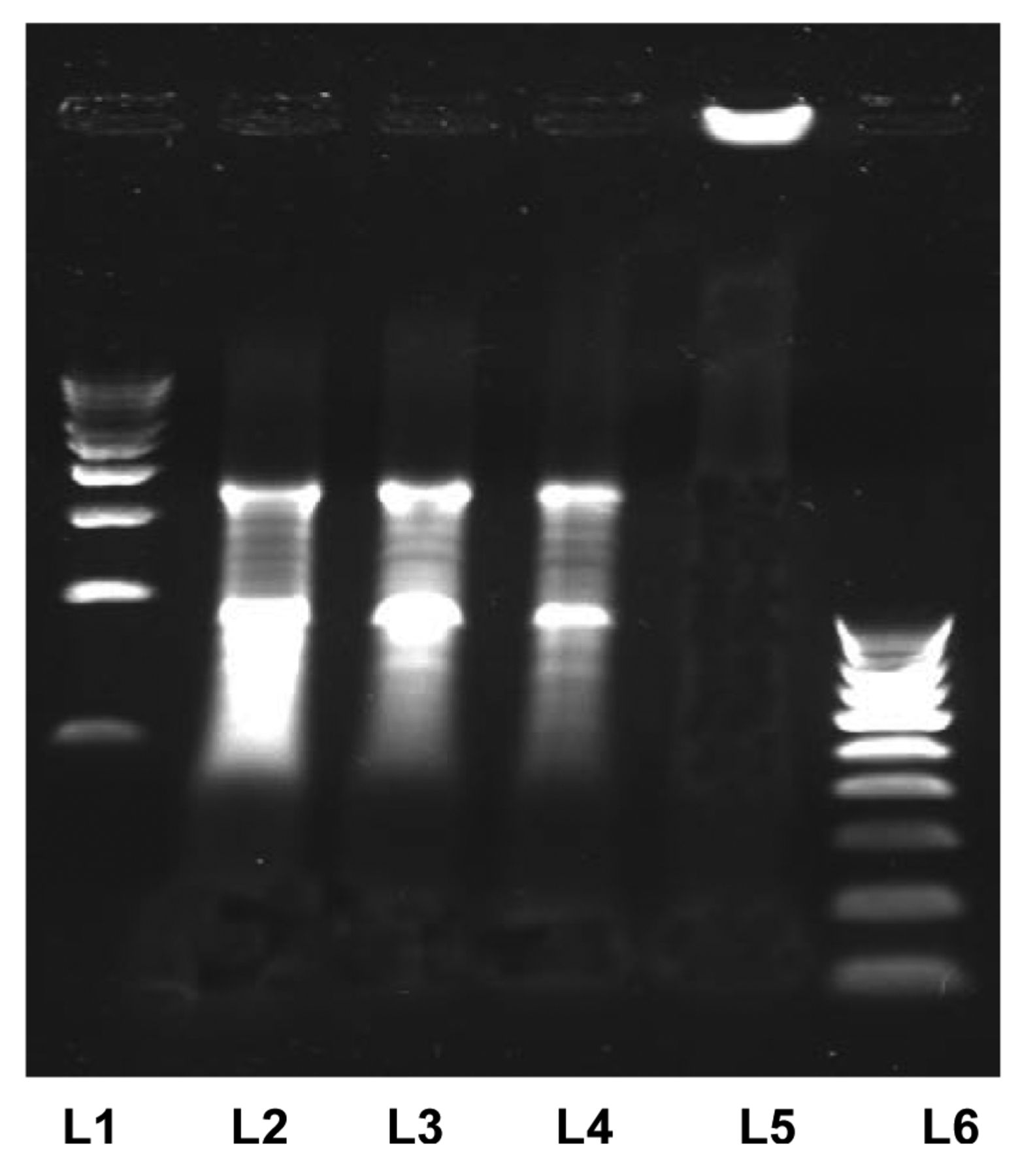
DNA fragmentation analysis. Lane 1 and 6: 500 bp, 100 bp Molecular weight markers; lanes 2-4: HeLa cells treated with fraction EHF-21 at 0.25, 0.50 and 1 μg/ml; lane 5: untreated HeLa cells. Fragmented DNA of the HeLa cells treated with fraction EHF-21 can be seen in lanes 2-4.
During apoptosis, a specific nuclease cuts genomic DNA between nucleosomes to generate DNA fragments and the presence of this ladder is a hallmark of apoptotic cell death (51). The ascidian E. viride fraction EHF-21 may be a DNA-damaging agent due to its intercalating properties, predominantly targeting cells in the G2 /M and S phases, which correlated with that of Ascididemin, a DNA-damaging agent due to its intercalating properties, which predominantly target cells in the S phase (18). Similarly, the present observation concurs with findings for Aplidine, which interfered with the synthesis of DNA and proteins and induced cell-cycle arrest (44). The inhibition of topoisomerase II and inducement of topoisomerase II-mediated DNA cleavage as noted in the potent cytotoxicity of Ascididemin, a pyridoacridine alkaloid of Didemnum sp. against tumor cells in vitro and in vivo (52) is also valid for the possible mechanism of activity of E. viride fraction. The discovery of a new cell-killing mechanism mediated by DNA nucleotide excision repair (NER) and interaction of ET-743 with the transcription-coupled NER machinery to induce lethal DNA strand breaks (53) corroborate the DNA fragmentation found in HeLa cells treated with E. viride fraction EHF-21.

The structure of Eudistomin H isolated from fraction EHF-21 of methanolic extract of Eudistoma viride.
Many novel and bioactive secondary metabolites have been isolated from the ascidian genus Eudistoma. Among the metabolites isolated from this genus, the predominant structure class was found to be β-carboline alkaloids (13). Macrolides (54), indoles (12, 55) and aromatic polysulfides (56) have also been reported from the same genus. Among them, the best known metabolites are the eudistomins, which are a series of β-carboline derivatives that display variety of pharmacological activities (57).
The present apoptogenic compound, a brominated indole derivative, isolated from the green ascidian E. viride is similar to that of β-carboline skeleton. Similarly, the genus Eudistoma from Korea yielded seven new beta-carboline-based metabolites with the presence of a benzoyl group attached to the beta-carboline nucleus at C-1, designated as eudistomins Y1-Y7 (58). The isolated compound from E. viride was considered potent when compared to that of Pibocin B, the first representative of marine alkaloids with a unique structural feature [(8 beta)-2-bromo-N-O-methyl-6,8-dimethylergoline], an N-O-methylindole group, isolated from the Far-Eastern ascidian Eudistoma sp., which showed moderate cytotoxic activity against mouse Ehrlich carcinoma cells (12). This simple aromatic nitrogenous compound could be a highly potent metabolite as evidenced from the strong cytotoxic activity. Similar bromine carrying beta-carbolines eudistomin E and eudistalbin A from the marine tunicate E. album were shown to possess cytotoxic activity (ED50 less than 5.0 ng and 3.2 μg/ml, respectively) in vitro against the growth of KB human buccal carcinoma cells (59). The beta-carboline alkaloid 14-methyleudistomidin C isolated from E. gilboverde exhibited potent cytotoxic activity with IC50s of <1.0 μg/ml against four different human tumor cell lines (60).
Although the mechanism of action of the E. viride fraction EHF-21 (Eudistomin H) was not ascertained, the induction of apoptosis and DNA fragmentation indicate the initiation of the pathway through cleavage of caspase-3, as the complex apoptosis mechanism involving extrinsic or intrinsic or perforin/granzyme B pathways ultimately converge on the execution pathway i.e. activation of caspase-3 (61) which leads to apoptosis, including chromosomal DNA degradation and formation of apoptotic bodies. But the observation of no cleavage of caspase after treatment with Ritterazine B from ascidian Ritterella tokioka and its induction of apoptosis via a novel anti-mitotic mechanism (43) poses an interesting question as activation of caspase-independent parallel cell death pathway by granzyme A pathway via single-stranded DNA damage, has been reported (62). Hence, the potent apotogenic compound Eudistomin H isolated here, with an IC50 of 0.49 μg/ml, could play an important role in overcoming chemoresistance in cancer cells through novel modes of action (63).
Acknowledgements
The Authors are thankful to Suganthi Devadason Marine Research Institute, Indian Institute of Science and National Center for Biological Sciences for the laboratory facilities and help. The Authors are also thankful to Dr. V. K. Meenakshi, APC Mahalakshmi College, Tuticorin for his help in identification of ascidian. R.P. Rajesh gratefully acknowledges the Ministry of Environment and Forests, Government of India for a fellowship desired from the project (grant no. 14/30/2003-ERS/RE). The first author (RRP) also acknowledges funding from DST-Young Scientist Scheme for works being carried out to complete this manuscript successfully with research grant DST No: SERB/F/405/2013.
- Received September 5, 2014.
- Revision received October 5, 2014.
- Accepted October 5, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.