


Abstract
Background: Despite the selectivity of Tumor necrosis factor Related Apoptosis-Inducing Ligand (TRAIL) for cancer cell killing activity, breast cancer cells are resistant to TRAIL-induced apoptosis for various reasons. Materials and Methods: From a functionally-characterized small-molecule dataset, CGP74514A was identified as a TRAIL sensitizer in MCF-7 breast cancer cells. Combination of sub-toxic dose of TRAIL with CGP74514A was evaluated in three TRAIL-resistant breast cancer cells, MCF-7, T47D and SK-BR-3. Results: In all tested cells, CGP74514A enhanced TRAIL sensitivity. Combination treatment triggered apoptotic events faster than single treatment. Regarding its mechanism of action, CGP74514A reduced cytosolic X-linked inhibitor of apoptosis protein (XIAP). Small interfering RNA-mediated knockdown experiments showed that reduction of XIAP is the reason of sensitization. Conclusion: CGP74514A sensitized breast cancer cells to TRAIL via reduction of XIAP expression level.
Breast cancer is the most common cancer in women regardless of race and ethnicity and represents the second leading cause of cancer death (1). Although chemotherapy and radiotherapy are widely used to treat breast cancers, identifying new therapeutic targets and characterizing related signaling pathways still remain the most important challenges for more efficient medical approaches.
Tumor necrosis factor Related Apoptosis-Inducing Ligand (TRAIL), a cytokine member of the tumor necrosis factor superfamily (TNF), triggers cell death signaling by binding to agonistic receptors, death receptor 4 (DR-4) and death receptor 5 (DR-5) harboring the cytosolic death domain (2). Although it is well-characterized that TRAIL kills cancer cells selectively with little toxicity in normal cells (3, 4), certain tumor-derived cell lines and most of patients' primary tumor cells are, regardless of the agonistic TRAIL receptor expression, resistant to this ligand (5). In particular, cancer cell lines derived from breast tumors are generally resistant to TRAIL (6) and the resistance mechanism of breast cancer cells have been identified to overcome the resistance of breast cancer cells to TRAIL. The identified resistance mechanisms include loss of agonistic receptors expression on plasma membrane (6), inhibition of expression/activity of pro-apoptotic players (7) and aberrant control of the cell death machinery by overexpression of anti-apoptotic proteins (8).
Cyclin-dependent kinases (CDKs) are a family of enzymes discovered for their role in cell-cycle regulation (9, 10). However, other roles in transcriptional regulation (11), nucleic acid processing (11) and neuron cell differentiation (12, 13) have been additionally characterized. Recently, inhibition of CDK activity is considered an attractive way to treat cancer (14). Although some CDK inhibitors are entering clinical trials, previous results using single-agent therapy are disappointing, suggesting a need for combination with other therapeutic reagents (15, 16).
In this report, we identified CGP74514A, a CDK inhibitor with selectivity for CDK1, as a sensitizer of TRAIL in MCF-7 breast cancer cells. Combination of TRAIL with CGP74514A induced reduction of XIAP in breast cancer cells with resulting apoptotic cell death.
Materials and Methods
Chemicals and reagents. CGP74514A was purchased from Sigma Aldrich (St. Louis, MO, USA). Purified human TRAIL (PHC1634) and Lipofectamine RNAiMAX reagents were from Life Technologies Inc. (Carlsbad, CA, USA). Caspase inhibitors Z-Val-Ala-Asp-fluoromethylketone (Z-VAD-FMK), Z-Ile-Glu-Thr-Asp-fluoromethylketone (Z-IETD-FMK) and Z-Asp-Glu-Val-Asp-fluoromethylketone (Z-DEVD-FMK) were from Tocris (Bristol, UK). CellTiterGlo cell viability assay kit was from Promega (Madison, WI, USA).
Compound screening. The compound screen was performed as previously described using functionally characterized compound collection (LOPAC-1280; Sigma) (17).
Cell culture. The MCF-7, T47D and SK-BR-3 human breast carcinoma cells were maintained in Roswell Park Memorial Institute (RPMI)-1640 supplemented with 10 % fetal bovine serum (Gibco, Rockville, NY, USA), L-Glutamine (2 mM) and penicillin–streptomycin (100 Units per ml).
Cell viability assay. Cell viability was evaluated using a conventional assay to read cytosolic adenosine triphosphate (ATP) content (CellTiterGlo assay kit; Promega). After 0.5 h of incubation with CGP74514A, the cells were treated with purified recombinant TRAIL for additional 24 h. Each well received 10 μl of assay solution, and raw luminescence values were acquired by Victor 3 plate reader (Perkin Elmer, Waltham, MA, USA). The raw values were transferred to a GraphPad Prism software (GraphPad, La Jolla, CA, USA) to evaluate relative cell survival. Mean of raw values from untreated wells were adjusted to 100 % survival after subtracting background luminescence values.
Immunoblotting. Crude cell extracts were made by scraping cells with cell lysis buffer (25 mM Tris–Cl with pH 7.0-7.4, 1% Nonidet-40, 400 mM NaCl, 1 mM MgCl2, 2 mM Na3VO4, 5 mM β-Glycerophosphate, 250 μM NaF, and 1X protease inhibitor mixture from Roche). Following protein concentration determination, 50 μg of protein was dissolved in sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). After transfer of protein to Immobilon-P polyvinylidene difluoride membranes (Millipore, Billerica, MA, USA), the membranes were incubated with 1:1,000 dilutions of primary antibodies against caspase-3 (CS-9665), BAD (CS-9292), XIAP (2045), cIAP (7065), Bid (CS-2002), Bcl-2 (CS-2872), Bcl-XL (CS-2764), Mcl-1 (CS-5453), death receptor-5 (CS-8074), poly-ADP ribosyl polymerase (PARP) (CS-9532), (all from Cell Signaling, Danvers, MA, USA). Antibody–antigen complexes were detected with horseradish peroxidase-conjugated immunoglobulin G (Bio-Rad, Hercules, CA, USA), and a chemiluminescent substrate (SuperSignal West Pico; Pierce, Rockford, IL, USA). The membranes were then stripped and probed with α-tubulin antibody (T9026, Sigma) for loading control.
siRNA experiment and cell viability evaluation. XIAP siRNAs (HSS100566: 5’-CCAGAAUGGUCAGUACAAAGUUGAA-3’, HSS100564: 5’-ACACUGGCACGAGCAGGGUUUCUUU-3’, HSS100565: 5’-GAAGGAGAUACCGU GCGGUGCUUUA-3’) and non-silencing siRNA (5’-ACGUGACACGUUCGGAGAAUU-3’) molecules were purchased from Dharmacon (Lafayette, CO, USA). Lipofectamine RNAiMAX reagent was used for transfection. Briefly, 5×105 cells were seeded in a well of 6-well dishes in 1.5 ml of culture media and incubated with 0.5 ml transfection mixture containing 25-40 pmol siRNA and 5 μl transfection reagent. After 48 hours of incubation, the cells were treated with recombinant TRAIL for additional 24 h and the cell viability was assessed using trypan blue exclusion. Knockdown efficiency was evaluated by western blotting using anti-XIAP antibody.
Results
CGP74514A enhanced TRAIL-induced cell death in breast cancer cells. We recently reported that breast cancer cells are resistant to TRAIL (17). To understand the resistance of breast cancer cells to TRAIL, we performed a chemical screen in order to identify new sensitizers of TRAIL in MCF-7 breast cancer cells (17). Sub-toxic dose of TRAIL (25 ng/ml) induced ATP contents reduction down to 0.75-fold of control, with no apparent cell death activation. We defined ‘hit sensitizer’ as a compound which reduced cell proliferation lower than 0.4-fold compared to control. From a collection of 1,280 bioactive compounds, we identified CGP74514A, a CDK inhibitor with selectivity for CDK1, as a strong sensitizer of TRAIL (Figure 1A). For cell line selectivity evaluation, we compared sensitization effect of CGP74514A in other breast cancer cells. When TRAIL-resistant breast cancer cells, MCF-7, T47D and SK-BR-3, were pre exposed to CGP74514A (5 μM), cells were sensitized to TRAIL (25 ng/ml)-induced cell death with minimal self toxicity of CGP74514A or TRAIL.
CGP74514A-sensitized breast cancer cells to TRAIL with enhanced apoptosis. To examine the characteristics of cell death sensitization, MCF-7 cells were subjected to qualititative dose-dependent analysis and apoptosis assay. First, trypan blue exculsion assay revealed that CGP74514A sensitized MCF-7 cells to a subtoxic dose of TRAIL (25 ng/ml) in a dose-dependent manner (Figure 2A). At 5 μM concentration, CGP74514A showed minimal toxicity while combination with TRAIL reduced cell viability to 35%. Secondly, incubation of cells with various caspase inhibitors including Z-IETD-FMK (25 μM, caspase-8 inhibitor), Z-DEVD-FMK (25 μM, caspase-3/7 inhibitor) and Z-VAD-FMK (20 μM, pan caspase inhibitor), inhibited cell death induced by combination treatment suggesting that CGP74514A sensitized MCF-7 cells by caspase-dependent cell death machinery. Lastly, we performed a DNA fragmentation assay using the same experimental settings. As Figure 2C shows, that combination treatment of CGP74514A with TRAIL induced DNA fragmentation, a marker of apoptotic cell death, while single-treatment of cells with TRAIL or CGP74514A did not show DNA fragmentation. Taken together, our results suggest that CGP74514A sensitized MCF-7 cells to TRAIL by triggering the apoptotic machinery.

CGP74514A sensitizes breast cancer cells to TRAIL. A: Assay plate where CGP74514A was identified as a TRAIL sensitizer. Normalized survival rate to control (y axis) was plotted against well position (x axis). Control wells received DMSO while the wells in far right received only a subtoxic dose of TRAIL (25 ng/ml). After acquiring raw luminescence values from each well, relative survival rates were evaluated by adjusting the mean value of control well to 1. The chemical structure of CGP74514A is provided. B: Sensitization effect of CGP74514A in breast cancer cells. The raw ATP content was determined in cells treated with CGP74514A (5 μM), TRAIL (25 ng/ml) and their combination for 24 h. Bars represent mean±SD (n=3).
CGP74514A reduced XIAP in breast cancer cells. To analyze the molecular mechanism of cell death modified by CGP74514A, apoptotic markers were examined for changes of the expression and activity in T47D cells (Figure 3). By combining incubation of TRAIL with CGP74514A, there was no remarkable expression changes of TRAIL receptor-1 and - 2 (DR-4 and DR-5). For the activation of apoptotic markers, caspase-3 was activated by combination treatment of CGP74514A with TRAIL, while poly ADP-ribose polymerase accordingly cleaved the p85 fragment. Bid, the shuttle protein which inter-connects intrinsic and extrinsic cell death signaling, was cleaved by combination treatment implying that caspase-8 was activated. We could not detect any substantial changes in the expression level of apoptosis-associated Bcl-2 family proteins, Mcl-1, Bcl-2, Bcl-XL and Bad. However, we could detect the reduction of X-linked inhibitor of apoptosis protein (XIAP), while expression of CIAP, another member of the IAP family protein, was not altered. The reduction of XIAP expression was mild by only CGP74514A, but amplified by combination treatment with TRAIL.
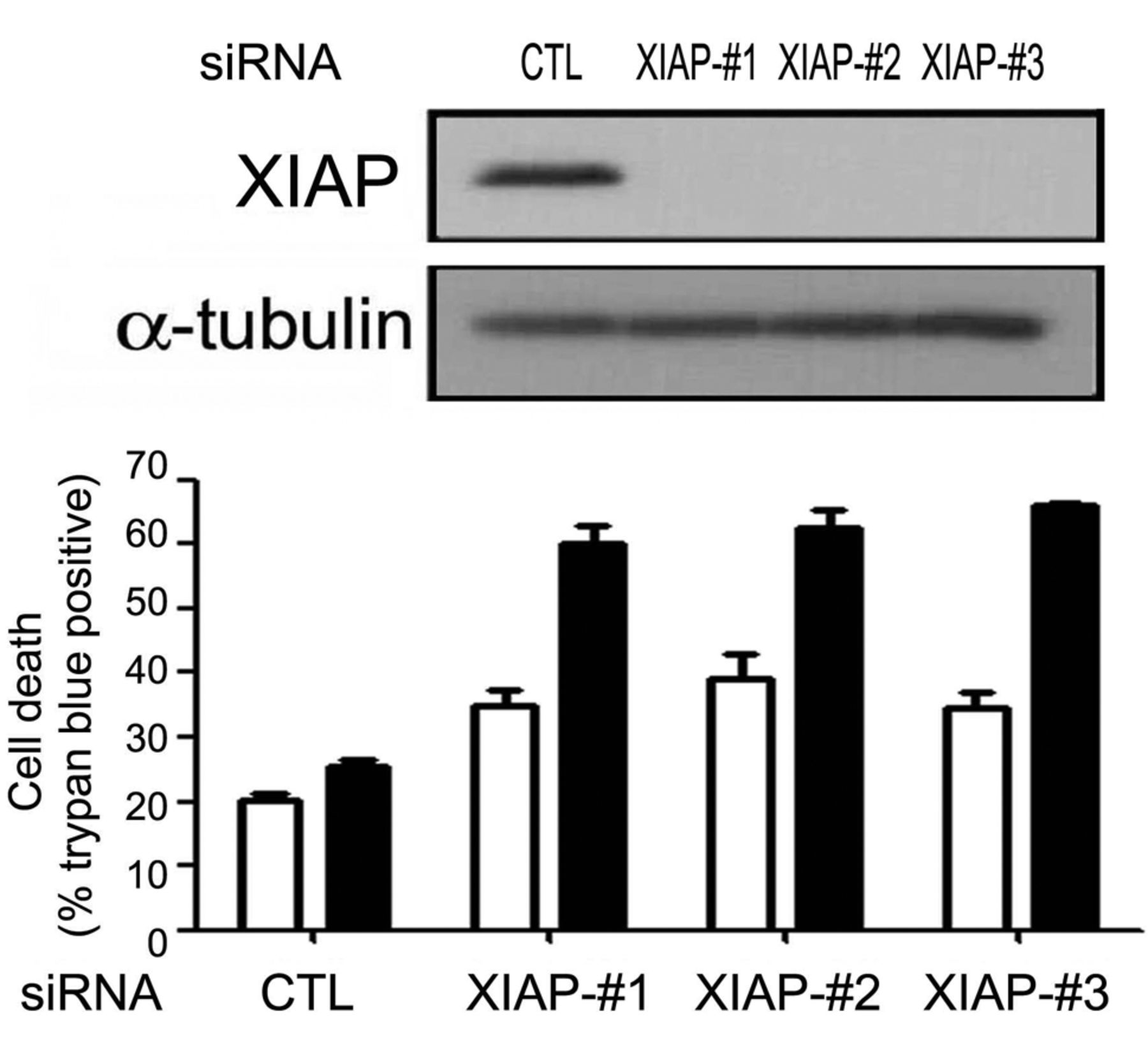
Down-regulation of XIAP caused sensitization of MCF-7 cells to TRAIL. To evaluate the effect of XIAP reduction by CGP74514A on sensitization, we artificially down-regulated the XIAP protein by small interference RNA and examined the sensitivity to TRAIL. We used three different siRNAs against XIAP-knockdown. As Figure 4 shows, all of three siRNAs were successful to down-regulate XIAP. When cells were exposed to sub-toxic concentration of TRAIL, XIAP-down-regulated cells showed 32%, 44% and 37% of cell death as evaluated by trypan blue exclusion, while control siRNA only marginally reduced cell survival rates. This data suggests that XIAP reduction by CGP74514A is the reason of sensitization to TRAIL in breast cancer cells.

CGP74514A triggers an apoptotic program for TRAIL. A: T47D cells were plated in 6-well plate, treated by CGP74514A with indicated dose with or without TRAIL (25 ng/ml) for 24 h. Trypan blue exclusion assay was performed to analyze late phase apoptotic cell death. The graph contains mean±SD values (n=3). B: Cell death sensitization of CGP74514A was inhibited by caspase inhibitors. Before exposure to CGP74514A, T47D cells were pre incubated with indicated caspase inhibitors for 1 h. The raw ATP content was determined and percent relative survival rate was calculated by adjusting the mean of control sample as 100%. Bars show mean±SD (n=3). C: DNA fragmentation was analyzed after incubation with indicated reagents.
Discussion
Anticancer therapeutics with no side effect are the ideal aims of oncology research. However, targeted cancer therapy with small molecule inhibitors cannot guarantee the safety of side effects because of the off target effect based on structural similarity. To get over this safety issue, biologic drugs with less off target effects and combination treatment schemes of pre-approved drugs are under investigation. At this point, human recombinant TRAIL or TRAIL receptor sensitizing monoclonal antibody drugs have been intensively studied, since the TRAIL-induced cell death system works in preferentially malignant or transformed cells. However, all clinical trials using TRAIL with single agent protocol failed because of weak activity for primary tumors (5). This disappointing outcome led investigators to research about the resistance of cancer cells to TRAIL and sensitizer discovery. Since breast cancer cells are usually resistant to TRAIL, they serve as a good model for sensitizer discovery and characterization of resistance mechanism(s).
Until now, we and other groups have identified several TRAIL sensitizers in breast cancer cells including a DNA damaging agent (daunorubicin) (17), a natural herbal product (curcumin) (18), an epigenetic protein inhibitor (MS-275) (19) and CGP74514A, a CDK inhibitor, as a TRAIL sensitizer in breast cancer cells which is presented in this study.
Inhibitors of CDKs are reported as promising agents for future clinical cancer therapeutics. More than 10 CDK inhibitors have been tested in clinical trials but previous clinical trials produced disappointing results (20) as the case with TRAIL was (5). These findings are probably due to the targeting of very conservative ATP-binding sites with low selectivity. However, they are still considered useful options for cancer treatment in combination with other anticancer therapeutic agents. For example, flavopiridol, a multiple CDK inhibitor, is very useful to treat p53 defective tumors in combination with vorinostat, a FDA-approved histone deacetylase inhibitor suggesting thus a good strategy to improve the use of CDK inhibitors (21). In this work, we report that CGP74514A, a CDK1 selective inhibitor, also represents another option. CGP74514A could sensitize breast cancer cells to TRAIL by inducing an apoptotic cell death program. The mechanism of action for CGP74514A includes, at least, the down-regulation of XIAP protein. A subtoxic dose of CGP74514A down-regulated XIAP in collaboration with TRAIL (Figure 3) while siRNA mediated knockdown of XIAP resulted in sensitization to TRAIL. These results suggest that the target of CGP74514A is related with the expression of XIAP. However, it is not clear whether reduction of XIAP is mediated by inhibition of CDK1, the original target of CGP74514A. Considering the structural similarity between CDKs, it is possible that other cyclin-dependent kinases, related with the XIAP expression, can be inhibited by CGP74514A.
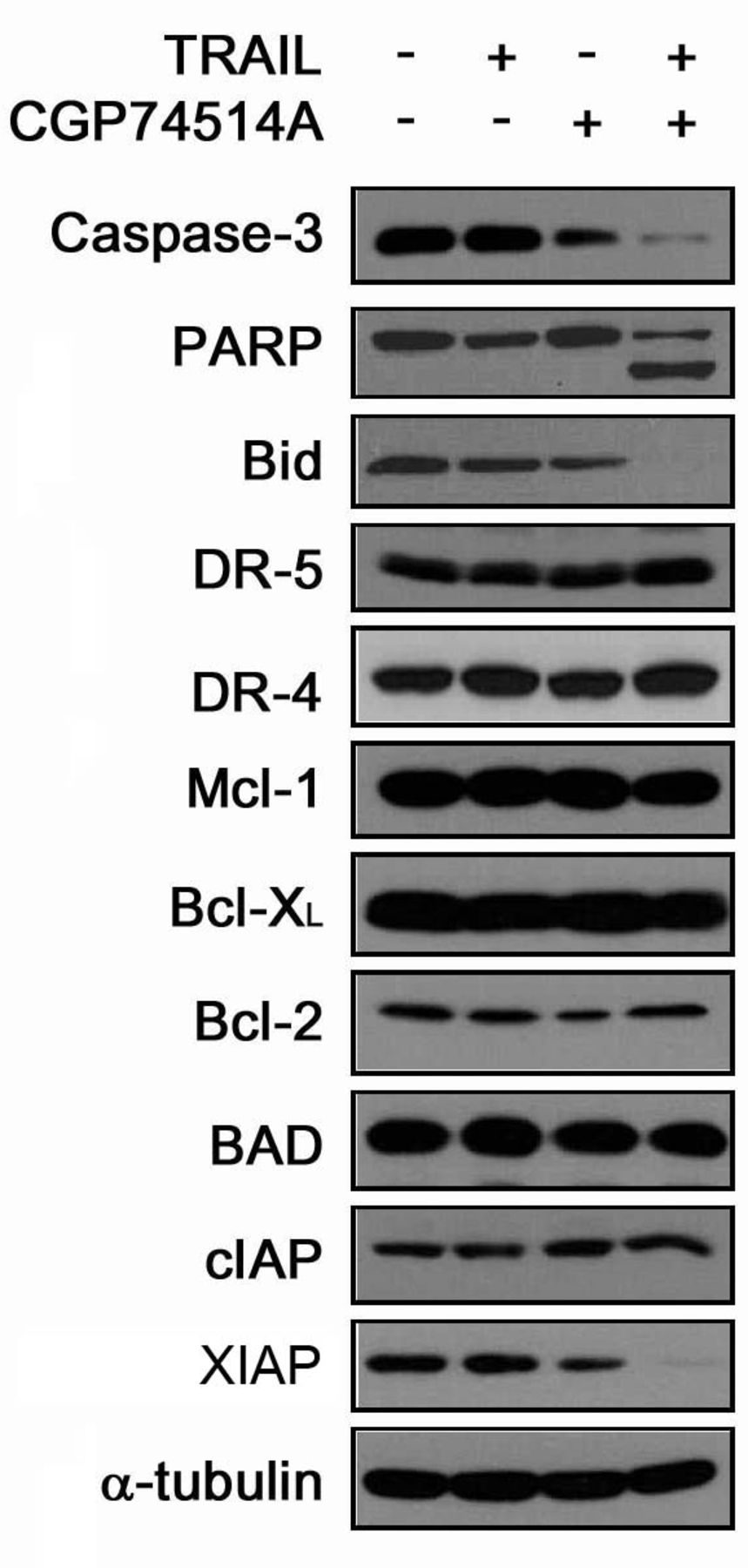
Combination of TRAIL with CGP74514A induces changes in apoptotic programs. T47D cells were treated with TRAIL (25 ng/ml), CGP74514A (5 μM) and the combination of both agents for 6 hours. After harvesting cells, the changes of apoptotic proteins were examined by immunoblotting.
Inhibitors of apoptosis proteins (IAPs) are members of a family of proteins serving for anti-apoptotic roles in cells (22). All members of this family have at least one baculovirus IAP repeat (BIR) domain by which protein-protein interactions are mediated for cell death control (23). From the large numbers of IAPs, XIAP (BIRC4) was identified as the direct inhibitor of caspase activity (24). It binds to effector caspases directly by BIR2 and BIR3 domains and inhibits activity by BIR2 and linker domain. Besides of caspase activity inhibition, C-terminal ‘Really Interesting New Gene (RING) domain’ serves a core domain for E3 ubiquitin ligase activity, and can directly control ubiquitination of caspase (25, 26). XIAP is known to be overexpressed in various cancers including breast carcinomas suggesting the possibility of this protein as an attractive cancer targets (27, 28, 29). Thus, a practical method to down-regulate IAPs, including XIAP, will be considered as an effective way to treat cancers. As our data suggest, CGP74514A appears to be a good example for down-regulating XIAP in breast cancer cells but future studies concerning the kinase selectivity of CGP74514A are required to shed light on the molecular signaling network controlling the expression of XIAP in cancer cells.

Reduction of XIAP-sensitized cells to TRAIL. T47D cells were transfected with three siRNAs of XIAP as described in Materials and Methods. After 48 hours of transfection, cells were treated with 25 ng/ml of TRAIL (black bars) and incubated for 24 h. Cell death was evaluated by trypan blue exclusion assay. The graph contains mean±SD values (n=2).
In conclusion, we propose CGP74514A as a strong sensitizer of TRAIL-induced apoptosis in breast cancer cells. Although none of TRAIL or CDK inhibitors have been approved for clinical treatment, we recommend that a low dosage combination of agents may be promising for clinical trial after animal model testing.
Acknowledgements
This work was supported by the International Research & Development Program of the National Research Foundation of Korea (Grant No. NRF-2012K1A3A1A07045330) and MRC grant of the National Research Foundation of Korea (Grant No. 2008-0062286) funded by the Ministry of Science, ICT and Future Planning (MSIP) of the Korean government.
Footnotes
-
↵* These authors contributed equally.
- Received April 3, 2014.
- Revision received May 26, 2014.
- Accepted May 27, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.