


Abstract
We used high- (ACCM) and low- (ACC2) metastasis cell lines of human adenoid cystic carcinoma (ACC) as an experimental model to study metastatic mechanisms and compare their expression levels for angiogenic-related factor vascular endothelial growth factor (VEGF). By using a series of extensive analyses, hypoxia-inducible factor-1 (HIF-1) α-dependent VEGF expression levels were observed to be higher in ACCM cell lines, increasing the possible development of tumor metastasis, compared to ACC2 cell lines. Our findings provide the novel insight that HIF-1α-dependent VEGF overexpression under hypoxic conditions shows to some extent associations with the metastatic tendency of ACC cells and may function as a potential target for ACC therapy.
Adenoid cystic carcinoma (ACC) is a slow-growing and highly malignant carcinoma of the salivary glands (1, 2). Currently there are no standard systemic therapies shown to benefit those afflicted with recurrent and/or metastatic ACC. Some reports have demonstrated that a possible mechanism of this distant metastasis is associated with angiogenesis (3-5), however, relatively little has been reported about reliable quantitative factors that may influence the spread and subsequent distant metastasis of ACC.
Various angiogenic factors such as vascular endothelial growth factor (VEGF), acidic and basic fibroblast growth factors (FGFs), transforming growth factor-β (TGF-β), produced by some types of tumor cells provoke mitogenic activity in surrounding capillary endothelial cells, leading to angiogenesis (6). Out of these factors, VEGF is considered to be one of the most notable and important regulators of both physiological and pathological angiogenesis, including the induction of endothelial cell proliferation, migration and survival, and capillary tube formation (7-9). The vegf gene is highly expressed in certain tumors and promotes angiogenesis and metastasis, with its gene expression being precisely controlled, not only at the transcriptional level, but also at the post-transcriptional level (10, 11). Of particular note is the involvement of hypoxic regulatory machineries via a typical hypoxia-associated transcription factor, hif-1, which is a heterodimer consisting of two basic helix-loop-helix proteins of the PAS family: a hypoxia-regulated α-subunit (hif-1α or hif-2α) and a constitutive β-subunit (hif-1β); hif-1α dimerized with hif-1β binds to specific hypoxia-response elements (HREs) in the vegf promoter for increased promoter activity in response to hypoxia (12, 13). Therefore, it is possible that the activation of HIF-1α is also associated with VEGF-induced angiogenesis and tumor vascularization and can serve as a clinical marker for aggressive ACC.
In the present study, a difference in HIF-1α-dependent VEGF expression levels was observed between the ACC2 (low-metastasis) and ACCM (high-metastasis) cell lines, suggesting that HIF-1α-dependent VEGF expression may be a good diagnostic marker for distant metastasis of ACC, and that the inhibition of HIF-1α may provide a new therapeutic approach to combatting ACC.
Materials and Methods
Materials. Anti-human HIF-1α was purchased from Santa Cruz Biotechnology (Dallas, TX, USA), and anti-human VEGF165 and anti-human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were purchased from R&D Systems (Minneapolis, MN, USA). PX-478 (14) was obtained from MedKoo Biosciences (Chapel Hill, NC, USA), and cycloheximide (CHX) as a protein translation inhibitor was commercially obtained from Wako Chemicals (Osaka, Japan).
Cell culture and hypoxic conditions. Cells from the ACCM and ACC2 cell lines were maintained at 37°C in Dulbecco's modified Eagle's medium (D-MEM) supplemented with 10% fetal bovine serum (FBS) in a 5% CO2 atmosphere. Hypoxia experiments were performed in a humidified triple-gas model IN VIVO2 hypoxic incubator (Ruskinn Technology, Guiseley, UK) calibrated to deliver 5% CO2, 2% O2, and 93% N2 at 37°C.
Bead-based assays. Bio-Plex Pro™ Assays, protein profiles using immunobead-based systems, were performed following the Bio-Rad systems protocol (Bio-Rad Laboratories, Hercules, CA, USA). Bio-Plex Pro™ Human Cytokine Standard Group I 27-Plex Panels of capture antibody-coated beads and labeled detection antibodies were used with the cell lysate samples. The median relative fluorescence units were measured using a Bio-Plex Assay Reader.
Plasmid and DNA transfection. The vegf promoter-derived firefly luciferase expression plasmid termed −1910/+379-LUC, 1HRE/WT-LUC, 2HRE-LUC, 1HREHM-LUC, and the HIF-1α expression plasmid pcDNA 3.1 hif-1α were obtained from Dr. T. Nishida (15). Sub-confluent ACCs in 96-well plates were transfected with one of the above plasmids (200 ng) and an internal control (20 ng), pRL-TK, containing a Renilla luciferase gene under the control of the herpes simplex virus thymidine kinase (TK) gene promoter (Promega, Madison, WI, USA), using Lipofectamine® 2000 transfection reagent (Invitrogen, Carlsbad, CA, USA). The Dual Luciferase System (Promega) was used for the sequential measurement of firefly and Renilla luciferase activities with specific substrates of beetle luciferin and coelenterazine, respectively.
RNA extraction and quantitative real-time polymerase chain reaction (PCR). Sub-confluent ACCs in 10-cm dishes were replenished with fresh medium containing 10% FBS, and cells were then incubated overnight before hypoxic exposure. Total cellular RNA was harvested at each time (0,1.5, 3, 6 and 12 h) using Isogen (Nippon Gene, Tokyo, Japan) following the manufacturer's protocol. Real-time PCR analysis was performed using a FastStart DNA Master SYBR Green I and Light Cycler apparatus (Roche Diagnostics, Mannheim, Germany). The following primers were used: for vegf165, 5’-tgtttgtacaagatccgcagacgtg-3’ and 5’-tcaccgcctcggcttgtcacatctgcaagtacgtt-3’; for hif-1, 5’-tgctcatcagttgcc acttcc-3’ and 5’-ccaaatcaccagcatccagaagt-3’; and for 18S ribosomal RNA, 5’-tcctgccagtagcatatgctg-3’ and 5’-agaggagcgagcgaccaaagg-3’.
RNA interference. For the present small-interfering RNA (siRNA) gene expression knock-down studies, siRNA for human HIF-1α (MISSION® siRNA Hs-HIF1A-2699-s, Hs-HIF1A-2699-as) and a negative control siRNA (MISSION® siRNA negative control) were purchased from Sigma (St. Louis, MO, USA). Each 21-mer siRNA duplex was transfected into cells using Lipofectamine® 2000 transfection reagent (Invitrogen, San Diego, CA) following the manufacturer's guidelines.
Immunoblotting. Immunoblotting was carried-out as described by T. Nishida (15). Primary antibodies against VEGF165, HIF-1α, and GAPDH were used at a dilution of 1:500. Protein bands were visualized using an enhanced chemiluminescence detection system.
Protein expression profile of hypoxic ACC2 and ACCM cells compared to that of normoxic cells
Statistical analysis. Unless otherwise specified, all experiments were repeated at least twice, with similar results. Statistical analysis was performed by Student's t-test, if necessary. Data were expressed as the mean±SD, and differences were considered significant at *p<0.05, **p<0.01.
Results
Protein expression profile of hypoxic high- and low-metastasis ACC cell lines. The protein expression profile of a variety of cytokines in ACC cells was compared between normoxic and hypoxic conditions using immunobead-based systems. A detailed comparison of the signals from their relative intensities led to the identification of VEGF, which showed a high ratio among a variety of cytokines in both cell lines. Furthermore, the extent of the ratio was higher at ~6.5-fold in ACCM cells than in ACC2 cells (~3.9-fold) (Table I).
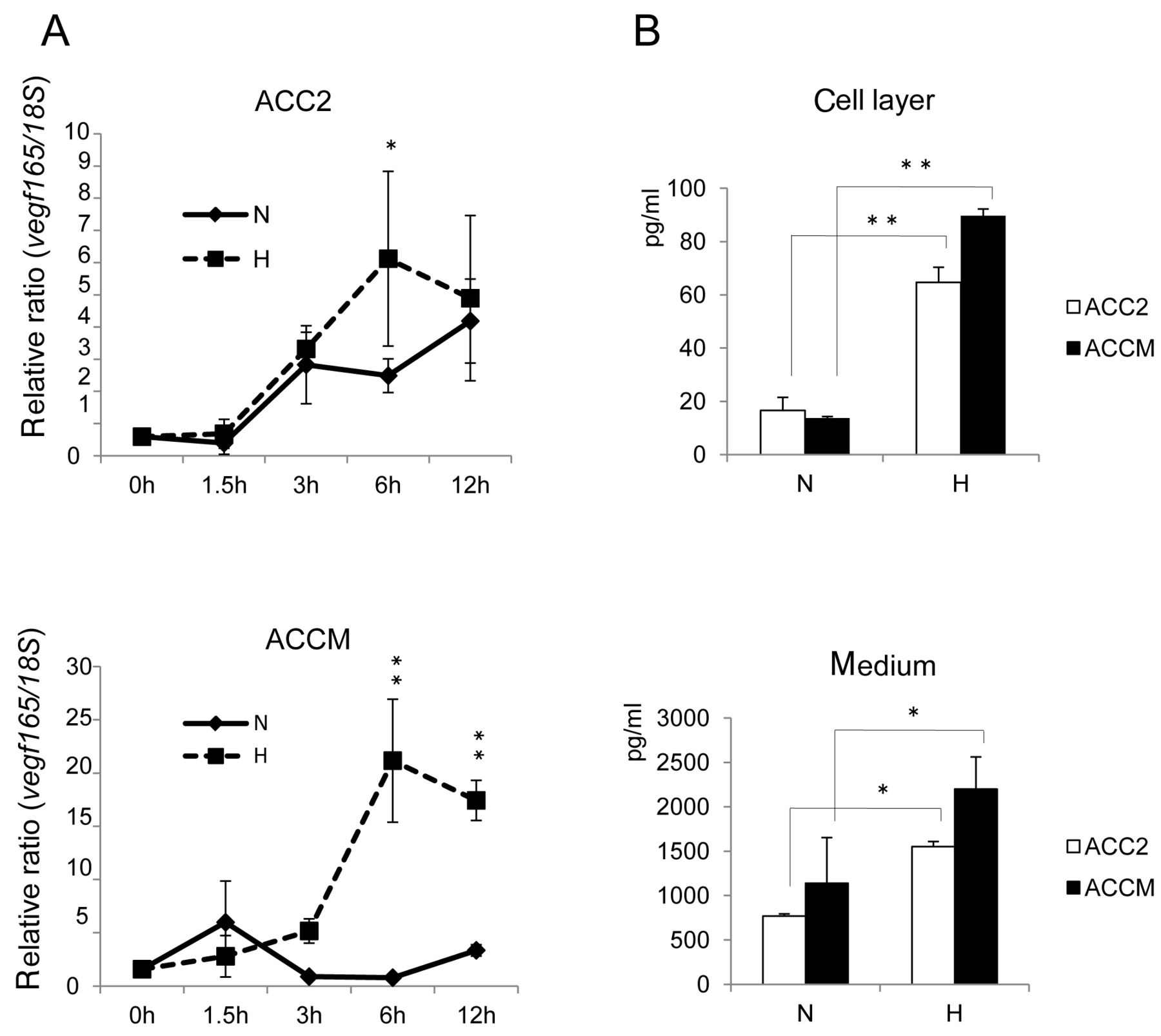
Hypoxia-enchanced vegf expression in ACC cells. (A) ACC cells were exposed to normoxic (N) or hypoxic (H) conditions for the indicated times. Total RNA was extracted and analyzed by real-time PCR for vegf165 mRNA expression. Relative ratio induction by hypoxic versus normoxic conditions at each point is shown. (B) VEGF protein synthesis enhanced by hypoxia. Quantification of growth factor was performed using Bio-Plex panels. Asterisks indicate significant differences from normoxia (*p<0.05, **p<0.01).
Hypoxia increased vegf mRNA and VEGF protein levels in ACC cells. As shown in Figure 1A, vegf mRNA expression was maximally up-regulated by 6 h of hypoxic exposure, with higher ratios than those under normoxia in both cell lines. The extent of the relative ratio was higher in ACCM cells (20-fold; 6 h) than that in ACC2 cells (6-fold; 6 h). Figure 1B shows that the VEGF protein levels in the cell layer and medium fraction in cultures of ACC cells exposed to hypoxia were significantly increased compared to those from normoxic cells, using immunobead-based systems.
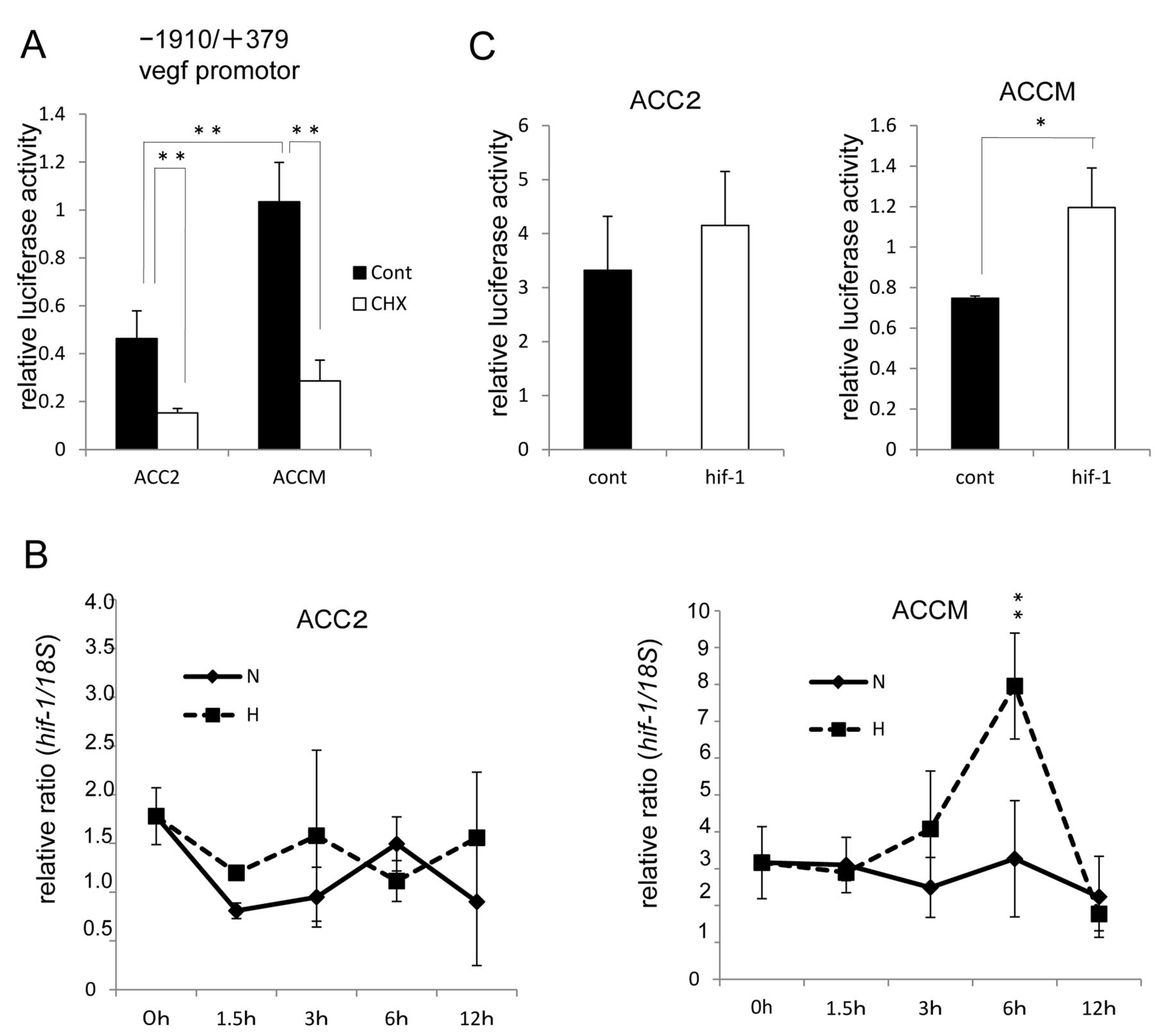
Elevated vegf promoter activities are HIF-1α-dependent in ACCM, but not ACC2, cell lines. To investigate the reason for the higher extent of VEGF expression levels in ACCM cell lines than in ACC2 cell lines, we next examined the association of vegf promoter via HIF-1α. As shown in Figure 2A, the activity of −1910/+379 vegf promoter with a corresponding reporter plasmid predominantly increased in ACCM, but not in ACC2. Notably, CHX inhibited the activation of the VEGF promoter in both cell lines. These data suggest that de novo synthesis of cellular proteins such as the transcription factor HIF-1α are required to mediate the activation of vegf promoter. Indeed, the time course of hif-1α gene expression by quantitative real-time PCR shows maximal up-regulation by 6 h of hypoxic exposure at levels higher than those under normoxia in ACCM cell lines (Figure 2B). Furthermore, to determine whether HIF-1α expression increases vegf promoter levels, we tested the exogenous overexpression of HIF-1α in both ACC cell lines using a pcDNA 3.1 hif-1α plasmid. As shown in Figure 2C, overexpression of HIF-1α increases the activation of the vegf promoter in the ACCM cell line, while in contrast, this activation was not observed in the ACC2 cell line.
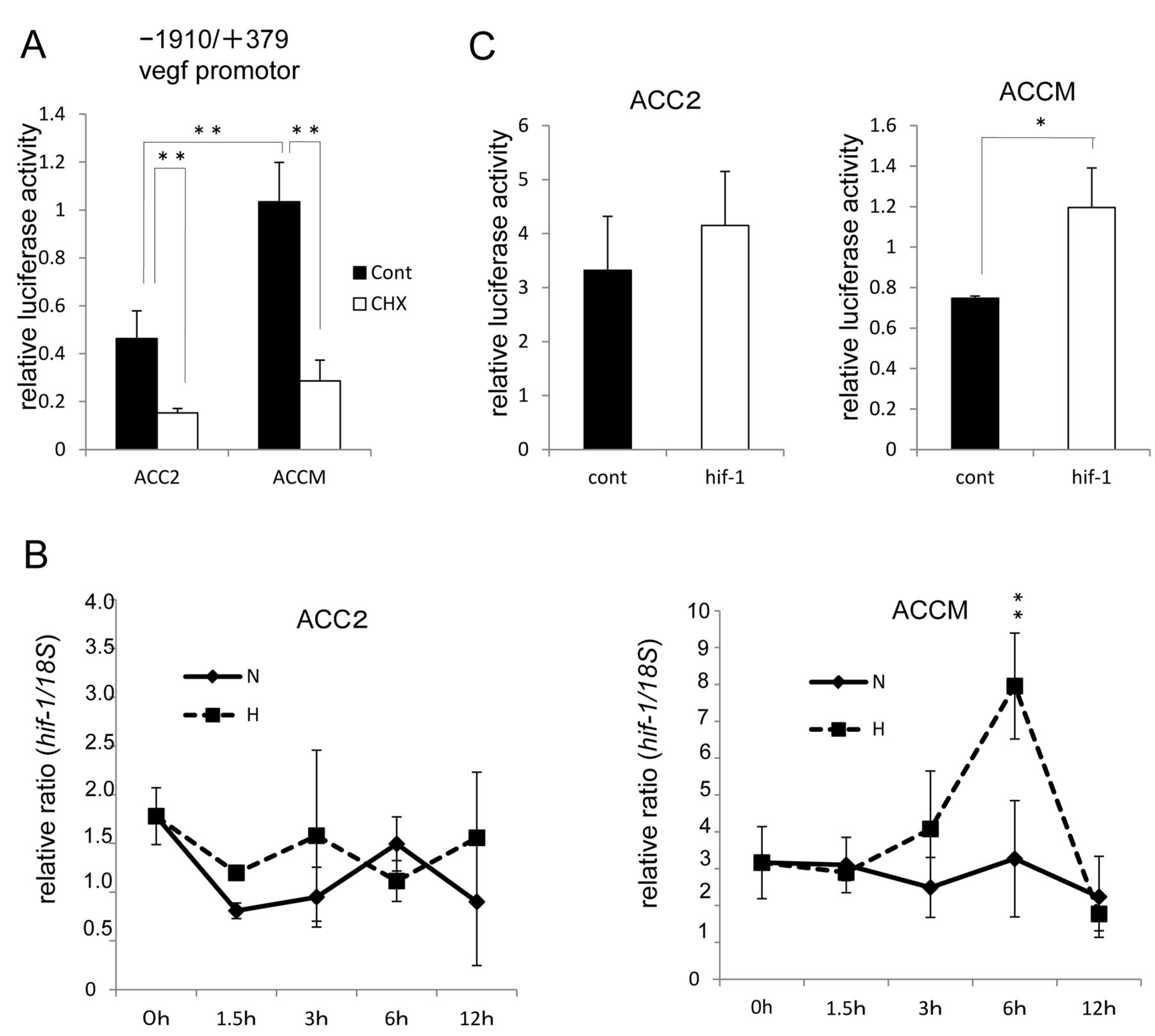
HIF-1α predominantly mediates the activation of vegf promoter in ACCM, but not in ACC2 cells. (A) Comparison of vegf promoter activity between ACC2 and ACCM cells. Addition of the protein synthesis inhibitor CHX inhibits the increased synthesis of a nascent transcription factor, for example, HIF-1α (**p<0.01). Columns are mean of duplicates, bars are SD. (B) Hypoxia-enhanced hif-1α expression in ACC cells. Total RNA was extracted and analyzed by real-time PCR for hif-1α mRNA expression. Relative ratio induction by hypoxic versus normoxic conditions at each point is shown (**p<0.01). Columns are mean of duplicates, bars are SD. (C) Effect of HIF-1α on vegf promoter activity in ACC cells. ACC cells were co-transfected with 200 ng of the vegf promoter (−1910/+379)-driven firefly luciferase reporter plasmid, 20 ng of TK promoter-driven Renilla luciferase reporter plasmid (pRL-TK, internal control), and 200 ng of HIF-1α expression plasmid (hif-1) or pcDNA3.1 GFP vector (cont) and then assayed for luciferase activities. Relative luciferase activities are presented as relative values of the measured luminescence of firefly luciferase versus Renilla luciferase (*p<0.05). Columns are mean of duplicates, bars are SD.
The fragment -1006/-954 containing a duplex HRE mediates activation of the vegf promoter by hypoxia or HIF-1α expression in ACCM, but not ACC2, cell lines. We attempted to assess the potential association of exposure to hypoxia and HIF-1α expression on vegf promoter in ACC cell lines by using fragment −1006/−954, which contains the single major HRE motif (1HRE/WT-LUC), its duplex (2HRE-LUC), and mutated HRE (1HRE/HM-LUC). As shown in Figure 3A and B, activation of the vegf promoter at the 1HRE/WT-LUC and 1HRE/HM-LUC plasmids tested in the ACCM cell lines is higher than that in the ACC2 cell lines, even though the extent of difference in promoter activity for each cell line between normoxia and hypoxia or expression and non-expression of HIF-1α, was almost identical. On the other hand, normoxia/hypoxia exposure or expression/non-expression of HIF-1α acts on ACCM cell lines, but not on ACC2 cell lines, to provoke the activation of the vegf promoter at 2HRE-LUC, as shown on the extreme right panel of Figure 3A and B.

The difference in the accessibility of HIF-1α binding to HREs on vegf in ACC cells. (A) Effect of hypoxia on the activation of vegf promoter due to the fragment -1006/-954. The firefly luciferase reporter construct, 1HRE/WT-LUC, or its mutant, 1HRE/HM-LUC, and 2HRE-LUC and TK promoter-Renilla luciferase reporter plasmid (pRL-TK, internal control) were each co-transfected and the cells were placed under the normoxic or hypoxic condition, then assayed for luciferase activity. Open bar, normoxia; closed bar, hypoxia (**p<0.01). (B) Effect of HIF-1α expression on the activation of vegf promoter by the fragment -1006/-954. pcDNA3.1-hif-1 or pcDNA3.1 GFP were additionally co-transfected in assays of vegf promotor by the fragment. Open bar, non-overexpressed HIF-1 (pcDNA3.1 GFP); closed bar, exogenously overexpressed HIF-1 (pcDNA3.1 hif-1) (**p<0.01).
The increased transcription factor HIF-1α contributes to the activation of VEGF expression in ACCM, but not ACC2, cell lines. To assess the functional loss of HIF-1α in VEGF165 protein expression in ACC cell lines, we first carried-out immunoblotting analysis using lysates from cells co-transfected with either HIF-1α siRNA or a negative control siRNA duplex. As shown in Figure 4A, a knockdown of HIF-1α protein expression by HIF-1α siRNA occurred at a dose of 400 pmol HIF-1α siRNA in ACCM cells. Therefore, this concentration was chosen for our next experiments. Although a knockdown of HIF-1α protein expression by HIF-1α siRNA does not fully affect the expression of VEGF levels under normoxia in both cell lines, it did inhibit the levels of hypoxia-induced VEGF expression. The extent of this inhibition in ACCM cell lines was higher than that in ACC2 cell lines (Figure 4B). Next, we confirmed the role of HIF-1α on VEGF protein expression using a chemical inhibitor of HIF-1 (PX478) in ACCM cells. Similar results were observed in HIF-1α knockdown cells: that is, the inhibition of HIF-1α protein expression by the addition of PX478 was found to prevent increased VEGF165 levels under the hypoxic condition in ACCM cells (Figure 4C).
Discussion
In the present study, we found that VEGF gene and protein levels are overexpressed in ACCM cell lines, increasing the possible development of tumor metastasis compared to ACC2 cell line. The result of VEGF-alone dominant expression among other cytokines and angiogenic factors is observed in the highly-metastatic ACCM cell lines compared to the less metastatic ACC2 cell lines (Table I, Figure 1). Thus, the most commonly identified protein, VEGF, can be associated with ACC tumor metastasis. However, it has been widely recognized that a single angiogenic factor may be secreted in an early tumor, while a more advanced tumor can produce many angiogenic factors (6). Although tumor-induced VEGF is a major factor in angiogenesis (7-9), such a discrepancy observed in highly metastatic ACCM cells is poorly understood at the molecular level. One possibility is that VEGF might regulate the expression of other angiogenic factors and/or signaling upstream and/or downstream in the cascade, leading to a change in metastatic phenotype depending on cell type (16).
Vegf is a known HIF-1α target gene. Our findings reveal that HIF-1α-elevated vegf promoter activities are higher in ACCM than in ACC2 cell lines (Figure 2). In addition, we found that under hypoxic conditions and/or in the presence of HIF-1α expression vector, duplex existence of HRE DNA sequence permits accessibility of its binding to HRE on the vegf promoter in ACCM cell lines (Figures 3A and B). The ability of HIF-1α to induce VEGF suggests that HIF-1α might affect metastatic ability through the induction of angiogenesis in these highly-metastatic ACCM cells. Following this speculation, we demonstrated that the suppression of HIF-1α activity through siRNA and PX478 significantly reduces VEGF expression levels in the highly-metastasic ACCM cell lines (Figure 4). Although it has been reported that inhibition of the VEGF pathway may result in lower anti-tumor effects in ACC (5), the suppression of HIF-1α expression through siRNA and PX478 may indirectly inhibit tumor cell proliferation and survival as well as directly inhibit VEGF expression in ACC cells.
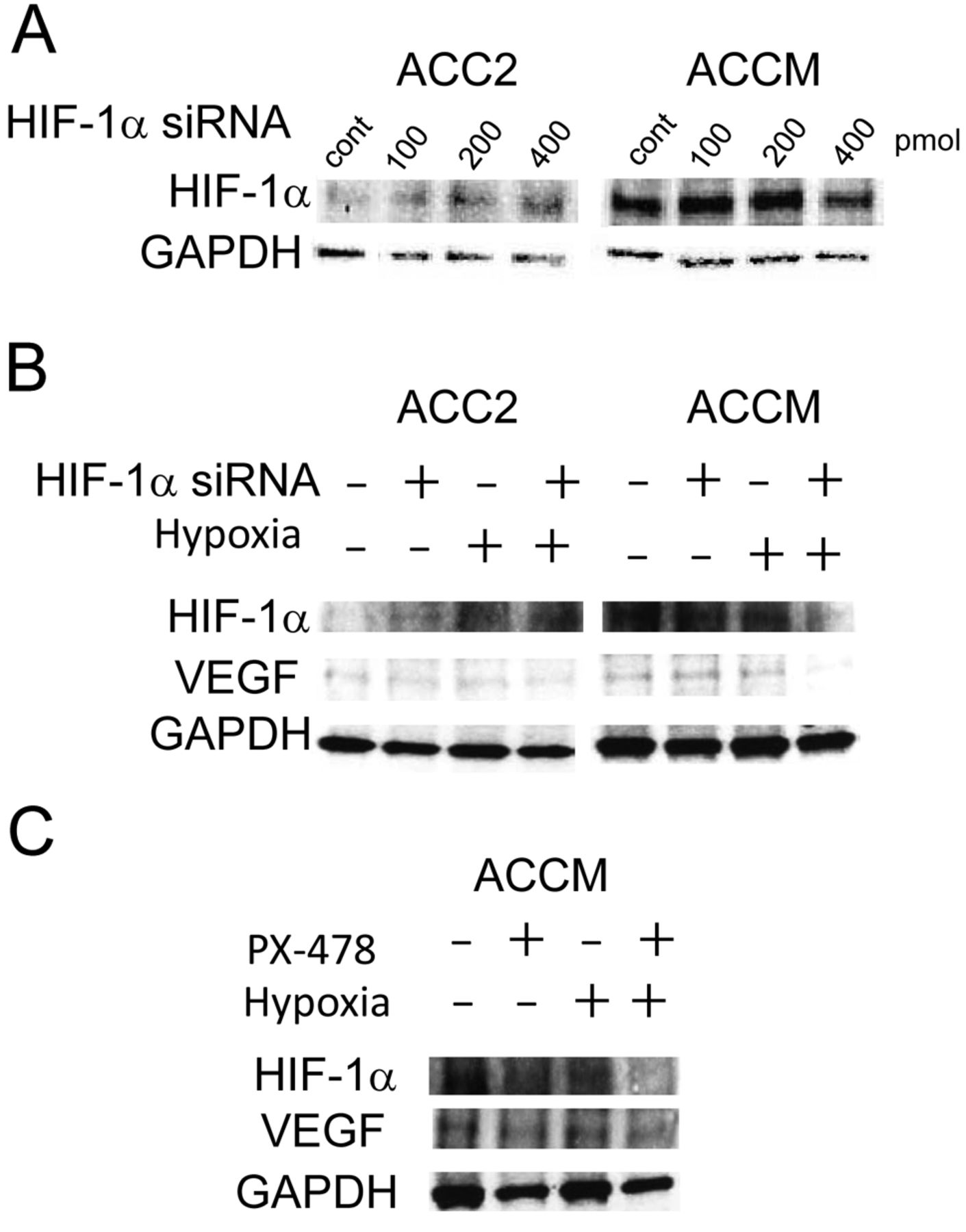
VEGF levels increased by hypoxia in ACCM cells are HIF-1α-dependent. (A) Cells were incubated under normoxic or hypoxic conditions for 18 h after transfection with either HIF-1α siRNA or a negative control siRNA duplex (cont). Immunoblotting showed the expression knockdown effects of HIF-1α siRNA on HIF-1α in ACC2 and ACCM cells. (B) Effects of HIF-1α siRNA on expressed VEGF165 protein levels under normoxia or hypoxia. Cells were incubated under normoxic or hypoxic conditions for 18 h after being transfected with either HIF-1α siRNA (400 pmol) or a negative control siRNA duplex. Immunoblotting results demonstrated that increased VEGF165 levels under hypoxic conditions are blocked in the presence of HIF-1α siRNA in ACCM cells. In contrast, HIF-1α siRNA did not block increased VEGF165 levels under hypoxic conditions in ACC2 cells. (C) Effects of a chemical inhibitor of HIF-1α (PX478) on expressed VEGF165 protein levels under normoxia or hypoxia. Cells were incubated under normoxic or hypoxic conditions for 18 h after the administration of PX478 (25 μM). Immunoblotting demonstrated that increased VEGF165 levels under hypoxic conditions are blocked in the presence of PX478 in ACCM cells.
The results of the present study indicate that HIF-1α-dependent VEGF expression is a major contributor to the high metastasic ability of ACC cell lines. These findings provide the novel insight that HIF-1α-dependent VEGF may be a potential target for the therapeutic treatment of ACC. Further investigation is required to fully examine this hypothesis.
Acknowledgements
This work was supported by Grants-in-Aid for Scientific Research (C) to S.K. from the Japan Society for the Promotion of Science.
Footnotes
-
Conflicts of Interest
The Authors declare that they have no conflicts of interest.
- Received January 4, 2014.
- Revision received January 15, 2014.
- Accepted January 16, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}