


Abstract
Background/Aim: It is well-established that HPV E7 proteins, encoded by human papillomavirus (HPV) genes, frequently associated with cervical cancers bind avidly to the retinoblastoma (RB) family of pocket proteins and disrupt their association with members of the E2F transcription factor family. Our previous study showed that the repressive p130-dimerization partner, RB-like, E2F and multi-vulval class (DREAM) complex was disrupted by HPV16 E7 proteins in order to maintain the viral replication in CaSki cells. However, we would like to address whether the activator B-myb-DREAM complex is critical in regulating the replication and mitosis phase since our previous study showed increased B-myb-DREAM expression in HPV-transformed cell lines when compared to control cells. Results: The association of B-myb with both LIN-54 and LIN-9 was equally decreased by depleting LIN-54 in CaSki cells. Flow cytometry analysis showed that LIN-54 depletion caused an increased proportion of G2/M cells in T98G, SiHa and CaSki cells. The mRNA levels of certain S/G2 genes such as cyclin B, aurora kinase A and Polo-like kinase 1 have demonstrated a marginal increased in CaSki-Lin-54-depleted cells when compared to SiHa- and T98G-Lin-54-depleted cells. We further confirmed this experiment by depleting the B-myb itself in CaSki cells and the results showed the same pattern of cell cycle and mRNA levels for S/G2 genes when compared to LIN-54- and LIN-9-depleted cells. Conclusion: The B-myb-DREAM complex might not be vital for progression through mitosis in cells lacking a G1/S checkpoint and not as crucial as the p130-DREAM complex for the survival of the HPV virus.
Cervical cancer accounts for almost 12% of all cancers in women and represents the second most common gynaecological malignancy in the world. Human papillomaviruses (HPVs) are associated with almost all cervical cancers. Infection with the high-risk HPV types 16 and 18 results initially in unobtrusive squamous intraepithelial lesions (SIL) in women. Most of these lesions are cleared 6-12 months after appearance, possibly due to the host's immunological defense.
High-risk HPV E6 and E7 oncoproteins expression are important for malignant transformation of cervical cancer cells. In particular, the E7 of high risk HPV binds to pRB family members (pRB, p107 and p130) for degradation. It has recently been discovered that the p107 and p130 pocket proteins are members of the mammalian dimerization partners, RB-like, E2F and multi-vulval class B (DREAM) complexes. The composition of mammalian DREAM is cell cycle regulated; being associating with E2F4 and p130 in G0/G1 and with the B-myb transcription factor in S/G2.
As with many cell cycle-associated transcription factors, B-Myb expression and function is dynamically regulated. The MYBL2 gene, which encodes B-Myb, is regulated directly by E2F transcription factors and is maximally induced at the G1/S boundary of the cell cycle (1). The trans-activation and gene regulatory potential of B-MYB is regulated by cyclin A/cdk2-mediated phosphorylation (2), while B-MYB is degraded through a ubiquitin-mediated process late in S phase (3).
A multiprotein unit has been identified in Drosophila melanogaster, which is involved in transcriptional repression. The complex is known as dREAM (Drosophila, Rb, E2F and Myb-interacting proteins) or Myb-MuvB (4). The homologs of all subunits of the invertebrate complexes have also been identified in human complexes, named DREAM or (LINC), whose composition is regulated at distinct phases of the cell cycle (4). The core DREAM complex contains Lin9, Lin37, Lin54, Lin52 and RbAp48 (the human homologues of Drosophila Mip130, Mip40, Mip120, dLin52 and Caf1p55, respectively). The complex dynamically interacts with pocket proteins/E2F-4 or B-myb in a cell cycle-dependent manner. In G0-G1, DREAM binds to E2F4 and either p130 or p107 represses transcription of E2F target genes regulating the G1/S transition (5). In S-G2, DREAM switches to B-MYB to activate genes required for G2/M transition and mitosis (6).

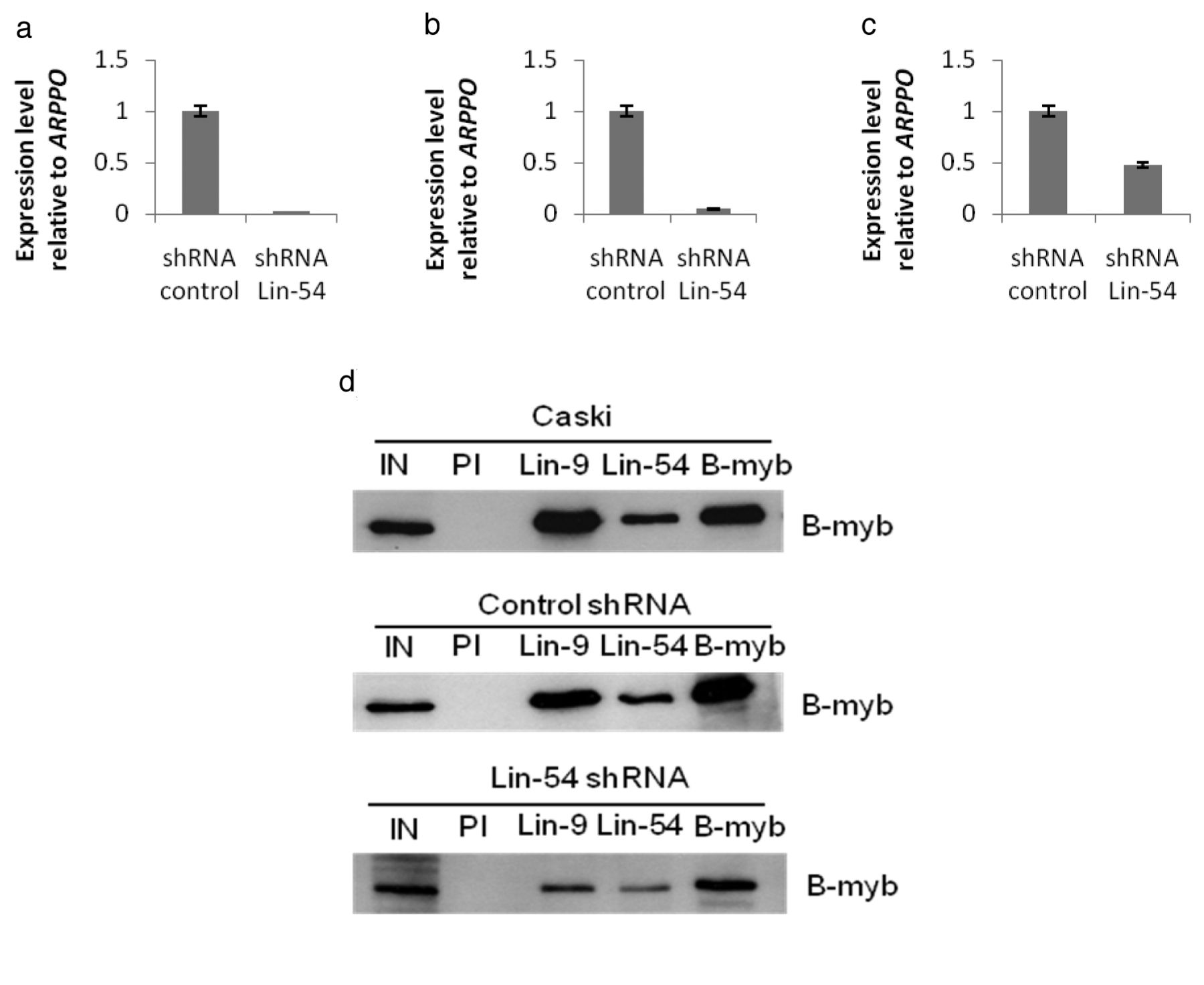
Depletion of Lin-54 in T98G, SiHa and Caski cells. Quantitative RT-PCR analyses of Lin-54 expression in Lin-54-depleted cells (a) T98G (b) SiHa (c) Caski relative to control cells. Expression was normalized to ribosomal ARPPO. (d) Nuclear lysates from Caski, Caski transduced with scrambled shRNA (control) and Caski transduced with Lin-54 shRNA cells were immunoprecipitated with preimmune Lin-9 rabbit serum (PI), Lin-9, Lin-54 and B-myb antibodies and Western blotted for B-myb. The input control (IN) comprised 10% of the lysates used in immunoprecipitation.
Knight et al., have shown that the B-myb-DREAM complex is required for transition through mitosis in embryonal stem cells, in which the retinoblastoma protein family is inactive (2). If the B-myb/DREAM complex is similarly required for mitosis in cervical carcinoma cells, particularly in CaSki cells, which also lack pocket protein function, targeting the complex may arrest cells in mitosis. Depleting LIN-54 in embryonal stem cells had a particularly pronounced effect in cell-cycle regulation. Therefore, under this understanding, we wished to investigate whether B-myb/DREAM has a similar function in HPV16-transformed cells, which also have deficient pocket protein/DREAM complexes. This question is significant, because previous experiments have shown increased B-myb/DREAM expression in CaSki and SiHa cells, at least when compared to control T98G cells, suggesting that disruption of the repressive p130/DREAM complex by 16E7 promotes the formation of the transcriptionally-active B-myb/DREAM complex, which could then have a positive impact on cell cycling.
Herein we attempted to address whether the B-myb/DREAM is critical for transit through mitosis in HPV16-transformed cells. We recognized that the LIN-54 is necessary for the stability of the DREAM complexes or its association with B-MYB. Based on propidium iodide-stained cells, the G2/M phase cells were not arrested when compared to control cells where the regulation of mitotic genes is similar to the control cells. Therefore, we suggest that E7-B-myb/DREAM complex is not really important for cell-cycle progression in HPV16–transformed cells.
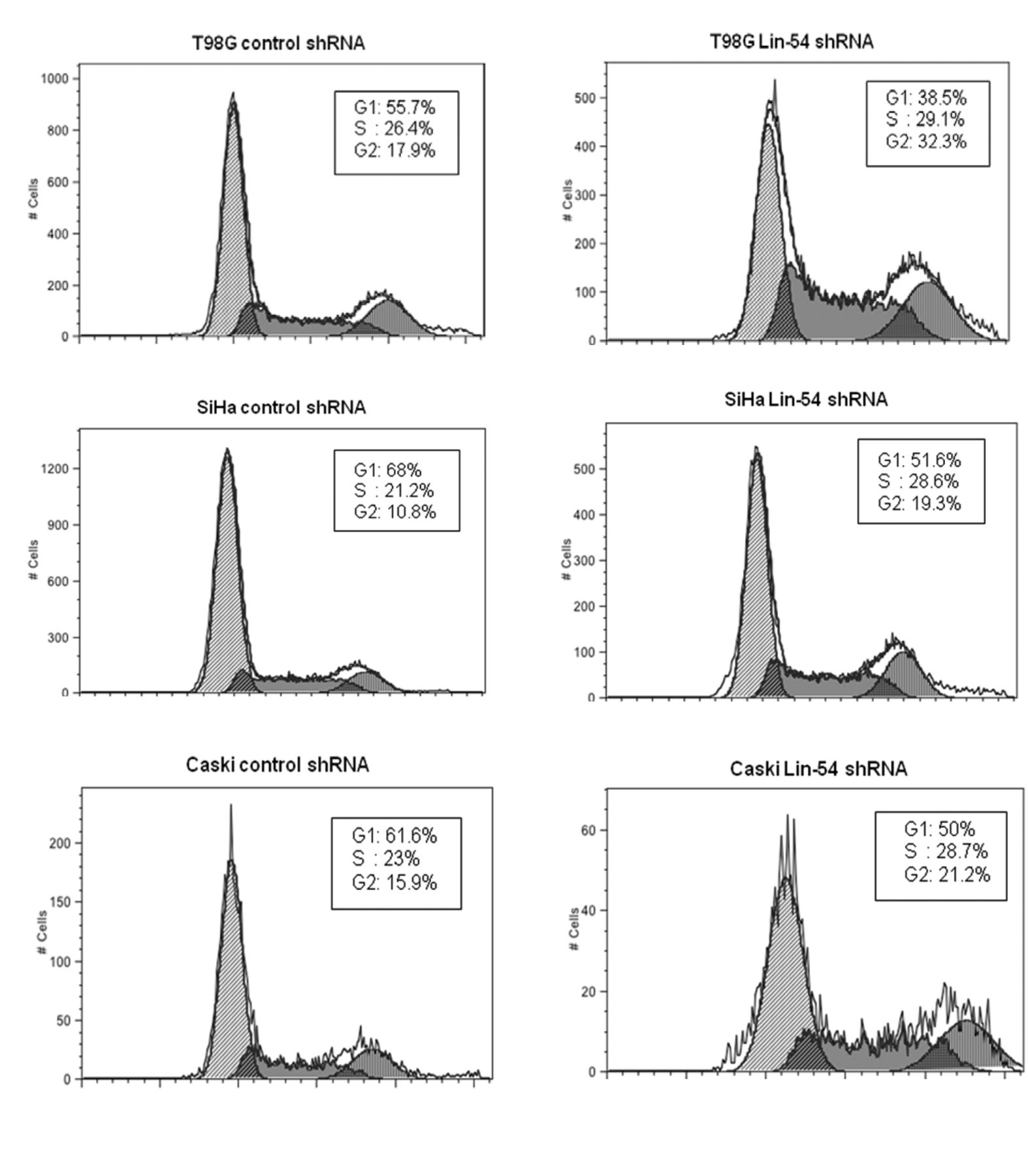
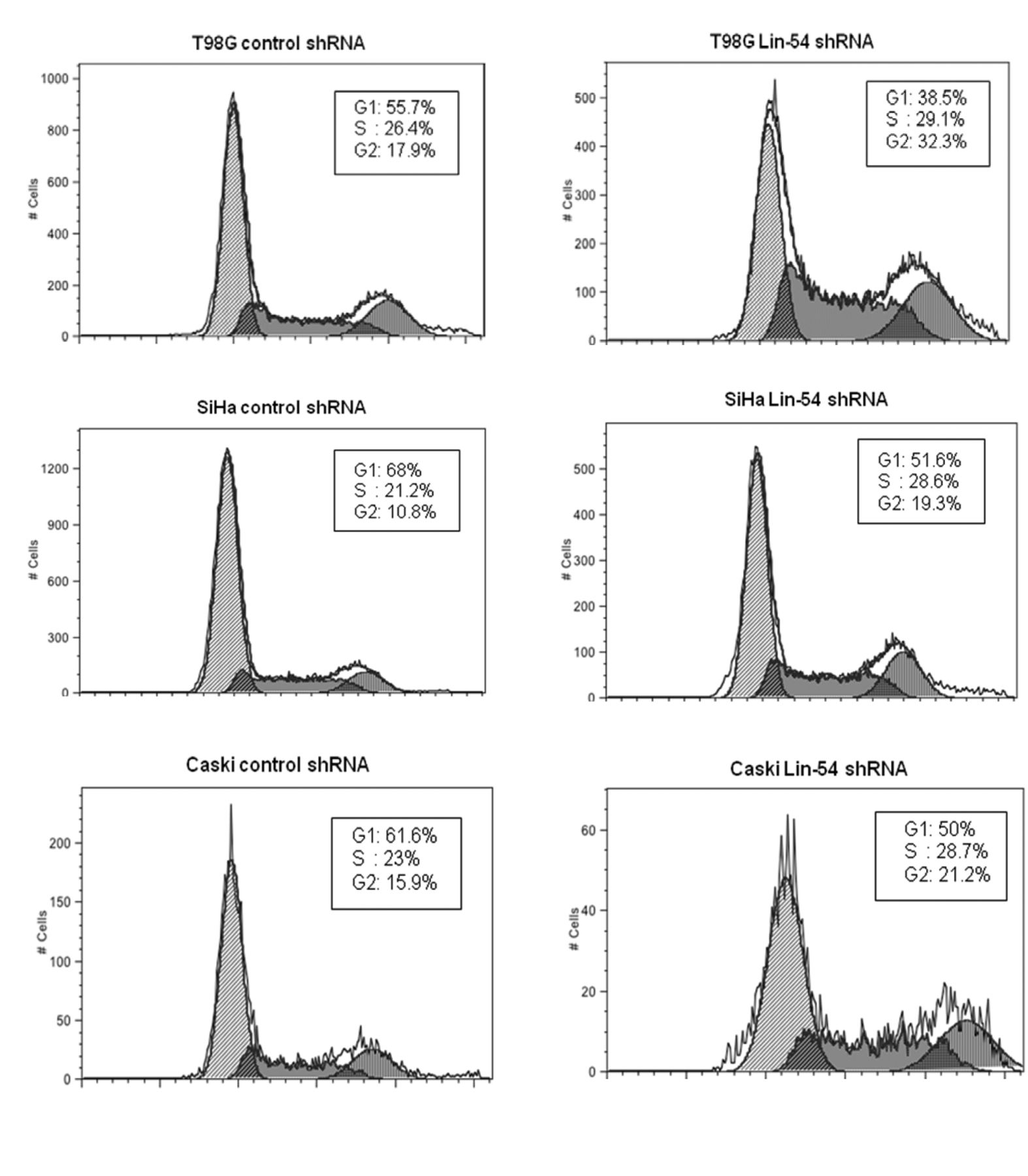
Effects on cell-cycle progression by depleting Lin-54 in T98G, SiHa and Caski cells. Flow cytometry of propidium-iodide-stained T98G, SiHa and CaSki cells tranduced with lentiviruses encoding control shRNA and Lin54 shRNA. The estimated percentages of cells in G1, S and G2/M phases are shown.
Materials and Methods
Cell culture. Human glioblastoma (T98G) and HPV16-positive cervical cancer (SiHa and CaSki) cell lines were maintained in Dulbecco's modified Eagle's medium (DMEM, Invitrogen, Paisley, Scotland) supplemented with 10% FCS, 100 U/ml of penicillin and 100 μg/ml of streptomycin. Incubation was carried out at 37°C in 10% CO2 and 95% humidity.
Antibodies. Polyclonal Lin-9 antibody (ab46415) used in western blot was from Abcam (Cambridge, UK), B-myb (sc-724), p107 (sc-318), and p130 (sc-317) antibodies were from Santa Cruz, Dallas, TX, USA, β-actin (A-2066) antibody was from Sigma Aldrich and HA (MMS-101R) antibody was from Cov¬ance (Denver, PA, USA). The B-myb LX015.1 monoclonal antibody was described previously (7). The Lin-9 rabbit polyclonal antibody (2) used for immunoprecipitation of DREAM complexes was purified by using a protein A-sepharose column (GE Healthcare, Buckinghamshire, UK).
Lentiviral transduction. Lentiviral particles carrying shRNA genes were generated in the pLKO.1 vector following the supplier's instructions (Cambridge, MA, USA). The four shRNAs used in this study were encoded by the following oligonucleotides inserted into EcoRI and AgeI sites of pLKO.1: Lin-54 (forward, 5’-C C G G C G G C T T C C AT T C A AT G G C AT A C T C G A G TATGCCATTGAATGGAAGCCGTTTTTG-3’, and reverse, 5’-AATTCAAAAACGGCTTCCATTCAATGGCATACTCGAGTATGCCATTGAATGGAAGCCG-3’); and p130 (forward, 5’-C C G G AT G G C C AT G T T T T G G A A G C A A C T C G A G TTGCTTCCAAAACATGGCCATTTTTTG-3’, and reverse, 5’-AATTCAAAAAATGGCCATGTTTTGGAAGCAACTCGAGTTGCTTCCAAAACATGGCCAT-3’). SiHa and CaSki cells (16106) were plated in 10 cm dishes and incubated at 37°C with 10% CO2. The next day, the medium was changed to fresh medium containing 8 mg polybrene ml21. The lentiviral stock was added at 1 ml for each 10 cm dish and the cells were further incubated overnight. Following transduction, fresh medium supplemented with 2 mg puromycin ml−1 was added 24 h post-transduction. Cells were harvested 48 h after puromycin selection for RNA and protein extractions.
Immunoprecipitation and Western blot analysis. Immunoprecipitation was performed as described previously (8). Proteins were resolved by SDS-PAGE, transferred to a nitrocellulose membrane and subjected to Western blot analysis using standard procedure.
Flow cytometry. Fluorescence-activated cell-sorting analysis was performed on ethanol-fixed cells stained with propidium iodide using a FACSCanto II analyser (BD Biosciences, San Jose, CA, USA) and analysed with FlowJo (Tree Star Inc, Ashland, OR, USA) software as described previously (2). At least 10 000 cells were analysed.
RNA extraction and qPCR. Total RNA was extracted by using an RNeasy Mini kit (Qiagen, Hamburg, Germany) and reverse-transcribed using SuperScript II reverse transcriptase (Invitrogen). qPCR was carried out using Absolute SYBR Green ROX (ABgene, Pittsburgh, USA) and an ABI 7900HT sequence detector (Applied Biosystems, Grand Island, NY, USA). Relative quantification was calculated by normalizing against ARPPO gene primers (2). Each PCR was performed on biological triplicates. Primer sequences and conditions were as described previously (2).
Results and Discussion
LIN-54 is necessary for the stability of the DREAM complex. Real-time polymerase chain reaction (PCR) was carried-out in order to investigate the silencing level of LIN-54 at the mRNA level. Three different cell lines were used in this experiment, T98G, SiHa and Caski. As shown in Figure 1a and b, the transduction of T98G and SiHa cells with LIN-54 shRNA resulted in a significant decrease of LIN-54 mRNA by 98.5% and 97.7%, respectively. However, only 58% of LIN-54 mRNA was reduced in Caski cells, as shown in Figure 1c. As the transduced cells were puromycin selected, the inefficient knockdown of LIN-54 in Caski cells should not reflect low transduction. There are several spliced forms of LIN-54 mRNA and it is possible that qPCR detected forms which were not targeted by LIN-54-1 shRNA. To further determine whether LIN-54 knock-down affected B-Myb/DREAM complex association in Caski cells, co-immunoprecipitation with three of the DREAM complexes constituents (Lin-9, Lin-54 and B-myb) was carried-out. The blot was probed against B-MYB, as B-MYB is highly expressed in Caski cells. As demonstrated in Figure 1d, the association of B-MYB with both LIN-54 and LIN-9 was equally decreased by depleting LIN-54 in Caski cells. This suggest that LIN-54 is necessary for the stability of the DREAM complex or its association with B-MYB.
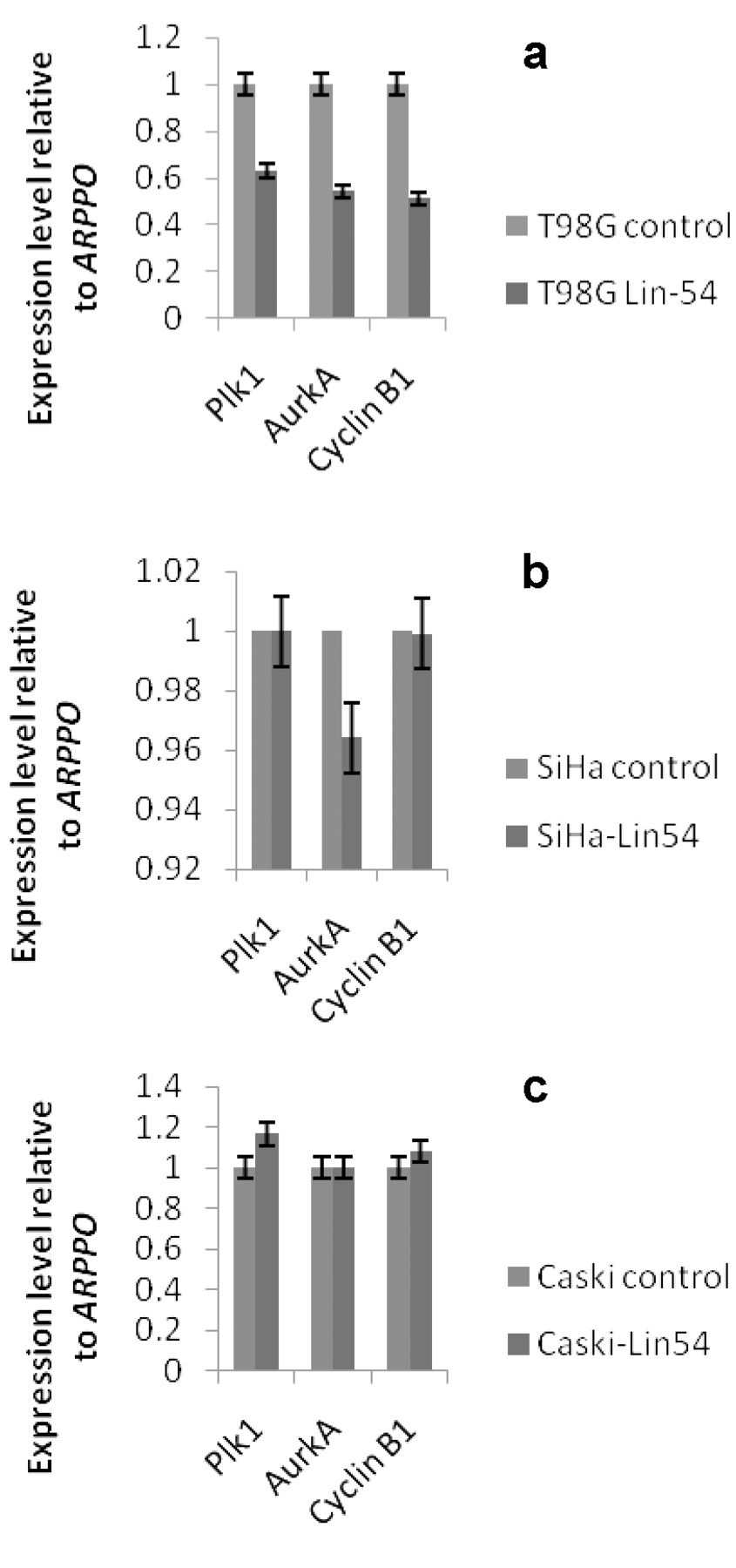
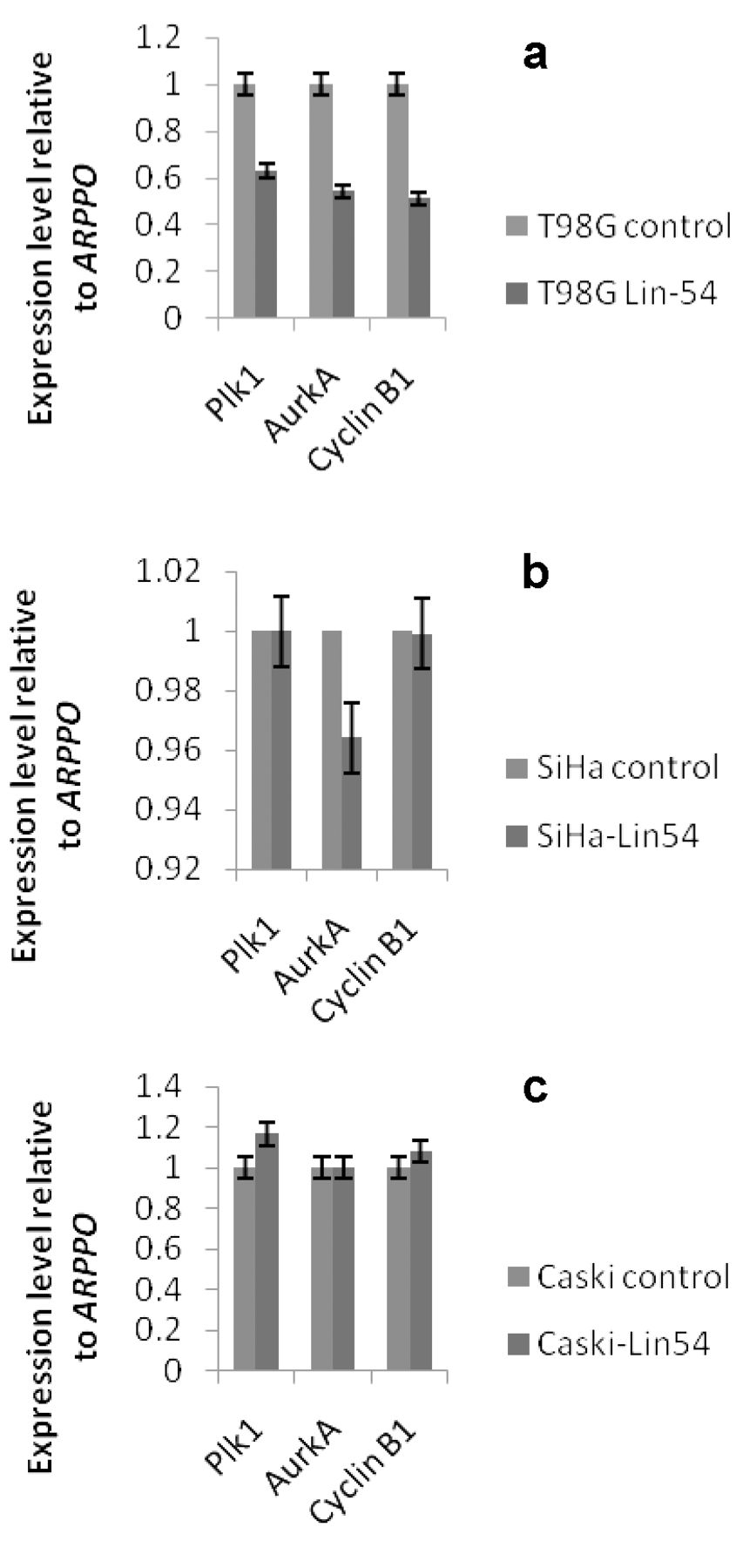
Analysis of G2/M gene suppression in Lin-54-depleted cells (a) T98G, (b) SiHa and (c) Caski. Quantitative RT-PCR analysis of B-myb target genes in Lin-54-depleted cells relative to control cells (scrambled shRNA). Expression was normalized to ARP PO.
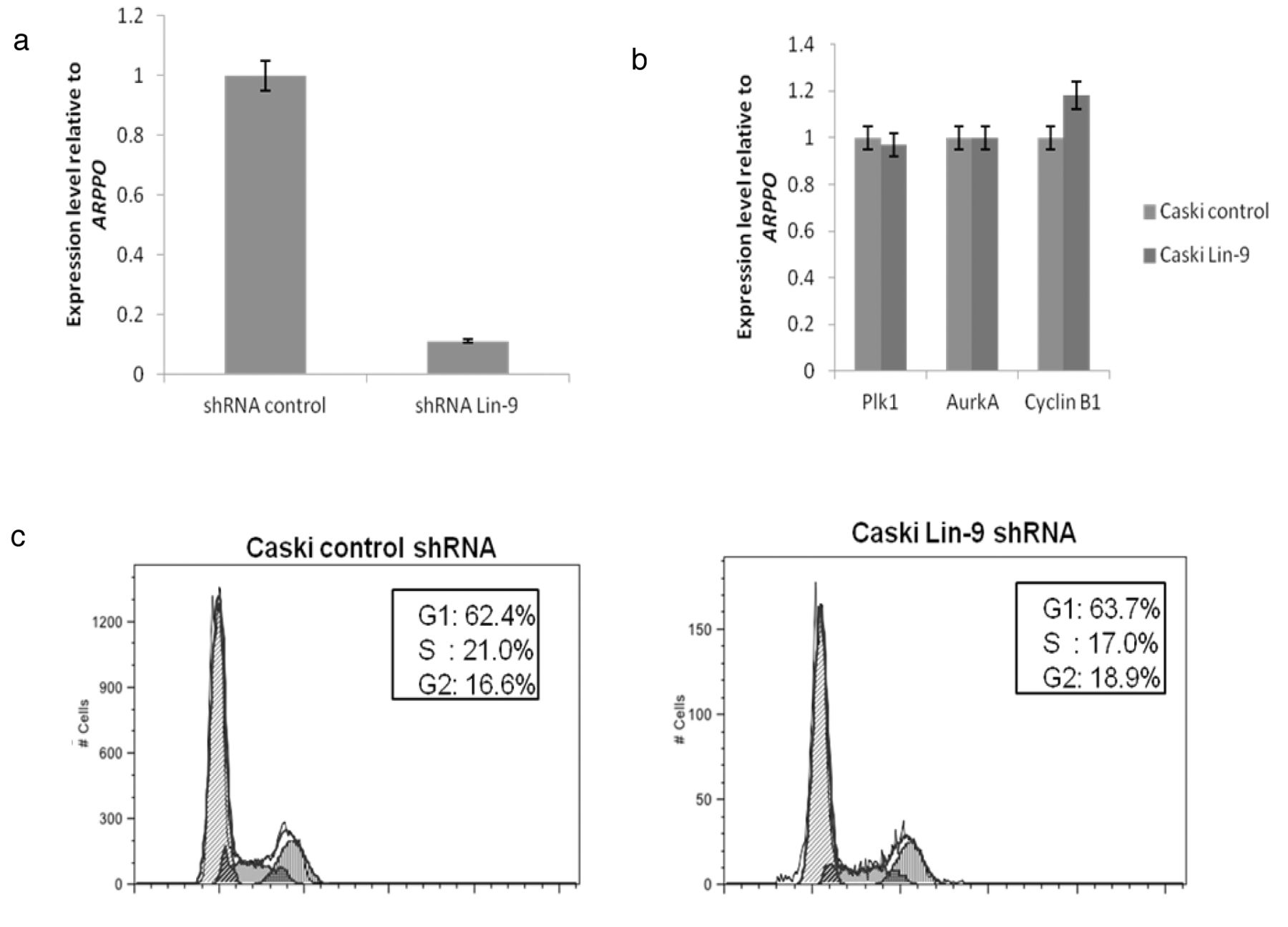
Depletion of Lin-9 in CaSki cells. (a) Quantitative RT-PCR analysis of Lin-9 expression in Lin-9-depleted cells. Expression was normalised to ribosomal ARPPO. (b) Quantitative RT-PCR analysis of Lin-9 target genes in Lin-9-depleted cells relative to control cells (scrambled shRNA) and expression was normalised to ARPPO. (c) Flow cytometry of propidium iodide-stained Lin-9-depleted CaSki cells. The estimated percentages of cells in G1, S and G2/M phases are shown.
G2/M phase cells increased in LIN-54-depleted cells. To determine the effects of LIN-54 suppression on cell-cycle progression, FACS analyses were performed. In this experiment, T98G, SiHa and Caski cells were employed and the transduced cells were harvested 48 h post-transduction. These cells were stained with propidium iodide and analyzed with flow cytometry. The results (Figure 2) demonstrated that LIN-54 depletion caused an increased proportion of G2/M cells in T98G and SiHa cells, with 32.3% and 19.3%, with 4n content, when compared to control, 17.9% and 10.8%, respectively. Depletion of LIN-54 in Caski cells also showed accumulation in G2/M phase with 21.2% with 4n content compared to control 15.9%.
To investigate how critical the B-myb/DREAM complex function is in Caski cells, we further determined the effect of LIN-54 knockdown on mRNA levels of certain S/G2 genes, such as cyclin B, aurora kinase A and Polo-like kinase-1. Theoretically, since depleting LIN-54 in Caski cells resulted in B-myb/DREAM complexes being down-regulated; S/G2 gene expression may be reduced in their expression. However, the expression level of these genes was marginally increased in Caski cells when compared to control shRNA cells (Figure 3). Whereas LIN-54 depletion resulted in expression of all three S/G2 genes being decreased in T98G cells, in SiHa cells only Aurora kinase A gene was decreased.
CaSki cells lack pocket proteins/DREAM complexes and the results demonstrate that in this context B-myb/DREAM complexes are not critical in regulating these S/G2 genes.
In addition, to better test the B-myb/DREAM complex function, we further knocked-down the other main components of core DREAM complex, which is LIN-9. We showed that the results were consistent with LIN-54 depletion in CaSki cells as shown in Figure 4; the mRNA levels of cyclin B, aurora kinase A and polo-like kinase-1 were increased when compared to control shRNA. Flow cytometry analysis showed that Lin-9 Caski-depleted cells is increased by 2% compared to 16.6%, in control cells as depicted in Figure 4c. These results showed that the B-myb/DREAM complex is not critical in regulating CaSki cells.

Depletion of B-myb in CaSki cells. (a) Quantitative RT-PCR analysis of B-myb expression in B-myb-depleted cells. Expression was normalised to ribosomal ARPPO. (b) Quantitative RT-PCR analysis of B-myb target genes in Lin-9 depleted cells relative to control cells (scrambled shRNA) and expression was normalised to ARPPO. (c) Flow cytometry of propidium iodide-stained B-myb depleted CaSki cells. The estimated percentages of cells in G1, S and G2/M phases are shown.
However, these results were further confirmed by depleting the B-MYB itself in CaSki cells. As anticipated, the results in Figure 5 showed the same pattern of cell cycle and mRNA level for S/G2 genes when compared to LIN-54 and LIN-9 depleted cells. Therefore, we can suggest that the B-myb/DREAM complex is not critical in cell-cycle regulation in CaSki cells.
Our results are contrary to those of Knight et al., in which they showed that, in embryonal stem cells, B-MYB recruited LIN-9 to activate transcription of G2/M genes in undifferentiated embryonal carcinoma cells (2). They also demonstrated that the B-myb/DREAM complex is vital for progression through mitosis in cells lacking a G1/S checkpoint. For cervical carcinomas, the B-myb/DREAM complex might not be as critical as the p130/DREAM for the survival of the HPV virus.
Acknowledgements
NNR was supported by the Ministry of Higher Education Malaysia under the Malaysia-Imperial Doctoral Program. The project was funded under grants, (BK009-2013) and (FP011-2012A). We thank Stefan Gaubatz for the Lin-54 antibody.
Footnotes
-
Authors' Contributions
RW and NNR contributed to the design of the study, NNR performed the experiments, NNR and RW carried out the analysis and interpretation of data, NNR drafted the manuscript, RW and RY contributed to the revision of the manuscript. All authors read and approved the final manuscript.
-
Coflicts of Interest
None to declare.
- Received June 25, 2014.
- Revision received August 7, 2014.
- Accepted August 11, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}