


Abstract
Background: The aim of the present study was to determine the effects of osthole on cell proliferation and viability, cell-cycle progression and induction of apoptosis in human laryngeal cancer RK33 and human medulloblastoma TE671 cell lines. Materials and Methods: Cell viability was measured by means of the MTT method and cell proliferation by the 5-bromo-2-deoxyuridine (BrdU) incorporation assay. Cell-cycle progression was determined by flow cytometry, and induction of apoptosis by release of oligonucleosomes to the cytosol. The gene expression was estimated by a quantitative polymerase chain reaction (qPCR) method. High-performance counter-current chromatography (HPCCC) was applied for isolation of osthole from fruits of Mutellina purpurea. Results: Osthole decreased proliferation and cell viability of cancer cells in a dose-dependent manner. The tested compound induced apoptosis, increased the cell numbers in G1 and decreased cell number in S/G2 phases of the cell cycle, differentially regulating CDKN1A and TP53 gene expression depending on cancer cell type. Conclusion: Osthole could be considered as a potential compound for cancer therapy and chemoprevention.
- Osthole
- proliferation
- apoptosis
- cell cycle
- high-performance counter-current chromatography
- RK33
- TE671 cells
Natural plant components, coumarins, have attracted great attention because of their pharmacological potential (1). Osthole (7-metoxy-8-isopenthenocoumarin), one of naturally-occurring coumarins isolated from dried fruits of Cnidium monnieri, Angelica pubescens and Peucedanum ostruthium used in traditional Chinese medicine (2), possesses anti-inflammatory (3, 4), anti-viral (5, 6), anti-coagulant (7), anti-convulsant (8, 9) and anti-allergic activities (10-13). Currently, osthole is investigated as a potential drug in the treatment of osteoporosis (14-17), as well as liver (18-22) and central nervous system disorders (23-26). Osthole also possesses anti-cancer properties and has been shown to inhibit cancer cell proliferation in human leukemia (HL-60) (27), cervical cancer U-937 (28) and HeLa (27), MCF7 (29) and MDA-MB-231 (30) human breast carcinoma cell lines, human prostatic cancer hormonosensitive LNCaP and hormonoindependent PC3, DU145 cell lines (31) or non-small cell lung cancer A549 line (32) and colorectal carcinoma (CoLo 205) cells (27). A single study reported in vivo anti-cancer effects of osthole in a murine-ype lymphocytic leukemia, P-388 D1 (33). Osthole also enhanced the cytotoxic activity of paclitaxel in vitro in breast cancer cells with HER2 receptor overexpression (34).
The aim of the present study was to evaluate the anticancer activity of osthole in two human tumor cell lines: laryngeal carcinoma RK33 and medulloblastoma TE671 cells. To determine the viability and proliferation, cancer cells were exposed to either culture medium-alone (control) or medium containing osthole (10-100 μM) for 72 h. High-performance countercurrent chromatography (HPCCC) was used for successful separation of osthol from natural source, a herb of Mutellina purpurea.
Materials and Methods
Apparatus. A Spectrum HPCCC apparatus (Dynamic Extractions, Slough, UK) was employed in the present study. The instrument was equipped with two multilayer PTFE coils: analytical and semipreparative (0.8 mm ID, 22 ml volume and 1.6 mm ID, 136 ml volume), respectively and working with the optimal speed of 1,600 rpm. The effluent was monitored at 320 nm by an ECOM Sapphire UV detector (Prague, Czech Republic). Identification of separated fractions was elucidated on an Agilent 1100 HPLC chromatograph coupled with 250 mm × 4.6 mm stainless steel column packed with 5 μm C18 (Zorbax Eclipse XDB-C18, Agilent Technologies, address) with a diode array detector (DAD) detector.
Reagents. Dichloromethane, n-heptane and ethyl acetate used for preparation of the extract from plant material was of analytical grade and purchased from Polish Reagents (POCH, Gliwice, Poland). Water was purified using a Millipore laboratory ultra pure water system (Simplicity™ system, Millipore, Molsheim, France). Methanol used for HPLC was of chromatographic grade (J.T. Baker Inc., city, The Netherlands).
Plant material. The plant material was collected in the Medicinal Plants Garden, Medical University in Lublin in June 2011. Aerial parts (herb) of Mutellina purpurea were dried at room temperature (RT), powdered and left for further extraction. Voucher specimens No ES17/18-19/2011 were deposited in the Department of Pharmacognosy with Medicinal Plant Unit, Medical University, Lublin. Fruits were air-dried at RT, powdered and 50 g was extracted with 500 ml of petroleum ether under reflux in 30 min. The procedure was repeated three times. The filtrates were combined and concentrated with a rotary evaporator to remove the solvent. The dried crude extract (4.9 g) was stored in a refrigerator for subsequent HPCCC separation.
Selection and preparation of two-phase solvent system. Two-phase solvent systems made of n-heptane, ethyl acetate, methanol and water (HEMWat) in different volume ratios were tested. Partition coefficient K, as a ratio of target substance in stationary to mobile phase, was determined by high-performance liquid chromatography (HPLC) analysis (comparing the peak area of the osthole both in upper and lower phases). A volume ratio 3:2:3:2 was chosen as the most proper for purification of target compounds (K=1.8). Then, the analytical column was first entirely filled with the upper stationary phase. The apparatus was rotated at 1,600 rpm and the lower mobile phase was pumped into the column at a flow rate of 1.0 ml/min. After hydrodynamic equilibrium was reached, 60 mg of extract dissolved in 1 ml of two-phase solvent system was loaded onto the column through a 1-ml injection valve. The solid phase retention was 70%. The effluent from the column was continuously monitored with a UV detector at 320 nm. Each one minute fraction was manually collected into test tubes. At the end of the run, the fractions were evaporated under reduced pressure and redissolved in methanol for subsequent HPLC analysis. Osthole was detected in fractions 40-45 with amount of 0.2 mg. The separation procedure was repeated in order to obtain the desirable amount. The crude methanol extract and each fraction obtained from the HPCCC separations were analyzed by HPLC. During HPLC analysis the flow rate was 1 ml/min, the column temperature was 25°C. A stepwise mobile phase gradient was prepared from methanol (A) and water (B). The gradient was: 0-5 min 50-60% A; 5-25 min 60-80% A; 25-30 min isocratic 80% A; 30-40 min 80-100% A.
The identification of isolated compound was carried out by comparison of retention time and UV-DAD spectra with those obtained by standards under the same conditions. The purity of osthole was 99% (established by the HPLC-DAD method).
Cell lines. Human medulloblastoma (TE671) was obtained from the Department of Medical Biology of Institute of Agricultural Medicine in Lublin. Human larynx carcinoma (RK33) was derived from a patient with diagnosed larynx squamous cell carcinoma. Cancer tissue was removed from the larynx after total laryngectomy and established as stable cell line, as previously described (35). Cell lines were maintained in Dulbecco's Modified Eagle Medium (DMEM) culture medium (Sigma, address) and 1:1 mixture of DMEM and Nutrient mixture F-12 Ham (Sigma) for RK33 and TE671 cells, respectively. All media were supplemented with 10% fetal bovine serum (FBS) (Life Technologies, Karlsruhe, Germany), penicillin (100 U/ml) (Sigma) and streptomycin (100 μg/ml) (Sigma). Cultures were kept at 37°C in a humidified atmosphere of 95% air and 5% CO2.
Cell viability assessment. Tumor cells were placed on 96-well microplates (Nunc, Langenselbold, Germany) at a density of 2×104/ml (RK33), 1x104/ml (A549, TE671). The folloing day the culture medium was removed and cells were exposed to serial dilutions of osthole in a fresh culture medium. Cell proliferation was assessed after 72 h by means of the methylthiazolyldiphenyl-tetrazolium bromide (MTT) method in which the yellow tetrazolium salt (MTT) is metabolized by viable cells to purple formazan crystals. Tumor cells were incubated for 3 h with MTT solution (5 mg/ml, Sigma). Formazan crystals were solubilized overnight in SDS buffer (10% SDS in 0.01 N HCl) and the product was determined spectrophotometrically by measuring absorbance at a wavelength of 570 nm using an Infinite M200 Pro microplate reader (Tecan, Männedorf, Switzerland).
Cell proliferation assay. The Cell Proliferation ELISA, 5-bromo-2-deoxyuridine (BrdU) Kit (Roche Diagnostics, Mannheim, Germany), was applied. Optimized cell amount (2×104) was applied on the 96-well plate (100 μl/well). The cells were treated with 100 μM concentrations of osthole for 72 h, followed by incubation with BrdU (100 μM). Cells were than fixed in FixDenat solution (30 min, RT). Monoclonal anti-BrdU antibodies coupled with horseradish peroxidase were subsequently added (90 min, RT) and detected using TMB (tetramethylobenzidine) substrate (30 min, RT). 1M sulphuric acid was added to stop enzymatic reaction and quantitation was performed spectrophotometrically at 450 nm using the Infinite M200 Pro microplate reader (Tecan).
Assessment of cell death. Measurement of cell death was performed using the Cell Death Detection ELISAPLUS kit (Roche). RK33 and TE671 cells growing on 96-well microplates were subjected to osthole (100 μM) for 24 h, whereupon supernatants were removed and cells lysed with 200 μl of lysis buffer for 30 min on ice. Subsequently, cell lysates were centrifuged at 200 × g for 10 min. and 20 μl of the sample were carefully transferred into the streptavidin-coated 96-well microplate. The immunoreagent (80 μl) containing anti-histone-biotin and anti-DNA-POD mouse monoclonal antibody was added and incubated under gentle shaking (300 rpm) for 2 h at 20°C. The solution was removed by tapping, each plate well was rinsed three times with 250 μl of incubation buffer and, finally, 100 μl per well of substrate, 2,2’-azino-bis(3-ethylbenzthiazoline-sulfonic acid) solution (ABTS), was applied and incubated at RT for 15 min on a plate shaker (250 rpm). Absorbance was measured at a wavelength of 405 nm using an Infinite M200 Pro microplate reader (Tecan).
Flow cytometry analysis. Experiments were performed using the FACSCalibur™ flow cytometer (BD Biosciences, San Diego, CA, USA), equipped with a 488-nm argon-ion laser. For cell-cycle analysis, cells were fixed in 70% ethanol at −20°C. After fixation, the cells were stained with propidium iodide utilizing the PI/RNase Staining Buffer (BD Biosciences) according to the manufacturer's instructions. Acquisition rate was at least 60 events per second in low acquisition mode and at least 10,000 events were measured. Cell-cycle analysis was performed by using a non-commercial flow cytometry analyzing software - Cylchred Version 1.0.2 for Windows (source: University of Wales, Cardiff, Wales, UK) and WinMDI 2.9 for Windows (source: facs.scripps.edu/software.html). The cells were acquired and gated by using dot plot FL-2 Width (X) versus FL-2 Area (Y)-gate to exclude aggregates and analyzed in histograms displaying fluorescence 2-area (yellow-orange fluorescence: 585 nm).
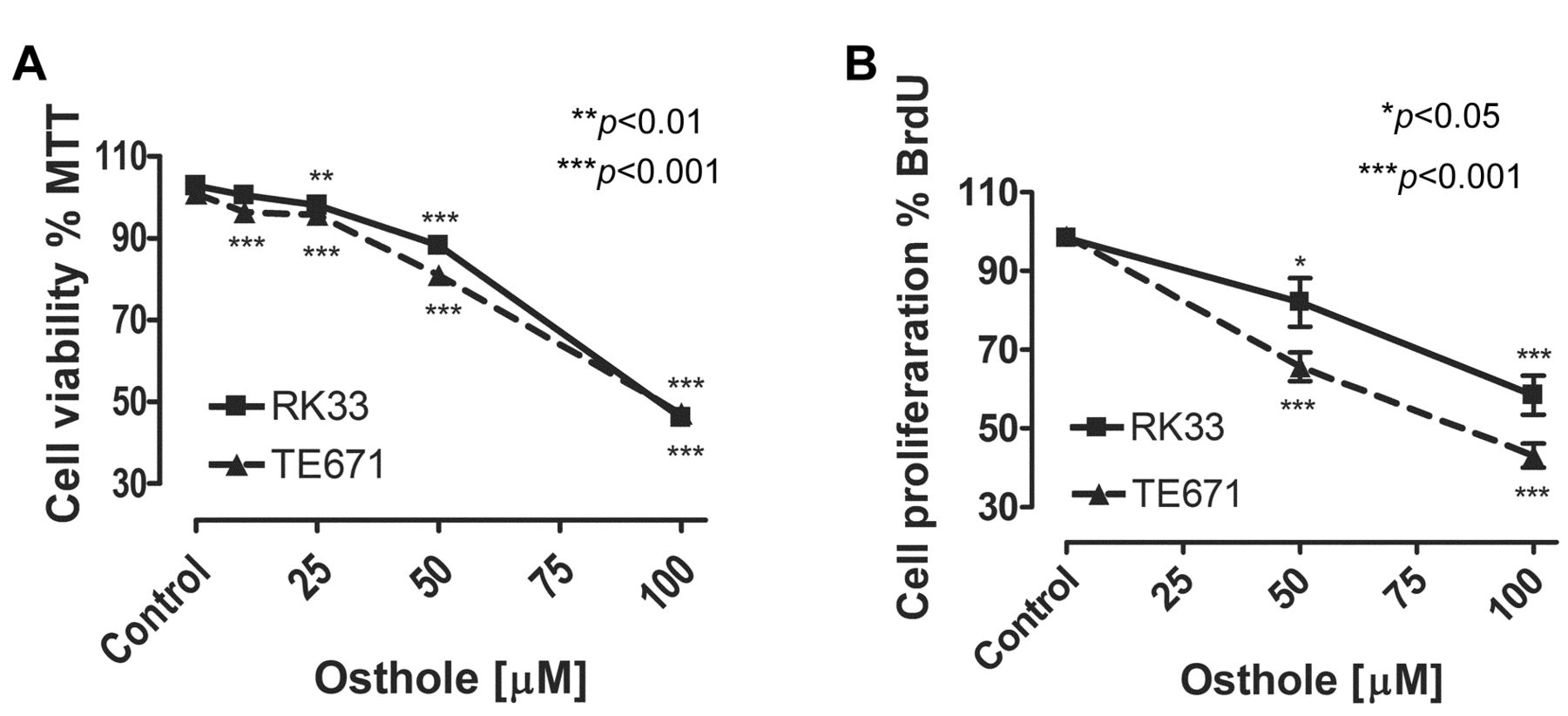
Osthole suppress proliferation of TE671 and RK33 cells. The cells were exposed to either culture medium-alone (Control), or osthole (10-100 μM) for 72 h. Normalized cell viability, measured by the MTT assay (A), and BrdU incorporation (B) is presented as mean±SEM at each concentration. Student's t-test revealed significant effect (*p<0.05, **p<0.01, ***p<0.001) of treatment with osthole compared to vehicle-treated cultures. This effect was also concentration-dependent, as judged by analysis of variance ANOVA test (***p<0.001). n=24 per concentration from three independent experiments.
RNA isolation and cDNA synthesis. Tumor cell lines RK33, A549 and TE671 were incubated on the 6-well microplates with the 100 uM concentration of osthole for 24 hours. Total RNA from the cells was isolated and digested with DNase using the High Pure RNA Isolation Kit (Roche) following the manufacturer's instructions. The RNA concentration was determined spectrophotometrically with a UV-VIS Genesys 10S spectrophotometer at 260/280 nm (Thermo Fisher Scientific, Madison, WI, USA). Three μg of total RNA was reverse transcribed for 30 min at 50°C using an oligo(dT) primer and the Transcriptor High Fidelity cDNA Synthesis Kit (Roche) followed by 5 min enzyme inactivation at 85°C according to manufacturer's instructions.
Quantitative polymerase chain reaction (qPCR). Quantitative real-time expression analysis was performed using a LightCycler® 480 II instrument (Roche). Analysis was performed utilizing Universal ProbeLibrary (UPL) hydrolysis probes specific for the TP53 and CDKN1A genes labeled with FAM in duplex with probe for reference gene GAPD labeled with Yellow 555 (Roche). The primers and probe sets were as following: TP53 (For 5’-CCCCAGCC AAAGAAGAAAC-3’, Rev 5’-AACATCTCGAAGCGCTCAC-3’, Probe 5’-GGATGGAG-3’); CDKN1A (For 5’-TCACTGTCTTGTACCCTTGTGC-3’, Rev 5’-GGCGTTTGGAGTGGTAGAAA-3’, Probe 5’-CCTGGAGA-3’); GAPD (For 5’-CTCTGCTCCT CCTGTTCGAC-3’, Rev 5’-GCCCAATACGACCAAATCC-3’, Probe 5’-CTTTTGCGTCGC-3’). Amplification was performed in 10 μl of reaction mixture containing cDNA amount corresponding to 12.5 ng of total RNA, 1 x LightCycler® 480 Probes Master (Roche) and appropriate set of 0.4 μM primers and 0.2 μM UPL hydrolysis probes for each target and reference duplex. After 10 min of initial incubation at 95°C, cDNA was amplified in 45-55 cycles consisting of 10 s denaturation at 95°C, 30 s annealing at 60°C and 10 s elongation at 72°C. Obtained fluorescence data was calculated using a relative quantification method with efficiency correction.
Statistics. The calculations were done by means of the Student's t-test and analysis of variance (ANOVA) test for multiple comparisons. Data are expressed as the mean±standard error of the mean (SEM) (*p<0.05, **p<0.01, ***p<0.001).
Results
Osthole was purified with application of the powerful technique of HPCCC in a very short time (40-45 min). We showed that osthole significantly inhibited cell proliferation and cell viability of TE671 and RK33 cells in a dose-dependent manner, as measured by means of the MTT assay. The threshold concentrations required to elicit antiproliferative effect in tumor cell lines were 10 μM for TE671 (p<0.001) and 25 μM for RK33 (p<0.01) cells. Incubation of RK33 and TE671 cell lines with 100 μM concentration of osthole caused a decrease in cell viability below 50% compared to non-exposed cells (Figure 1A). The effect of osthole on cancer cell proliferation was attributed to decreased cell division, as determined by decreased incorporation of BrdU. Incubation of tumor cells with 100 μM of osthole resulted in more than 50% reduction of BrdU incorporation (Figure 1B), which mimics the inhibition of cell viability measured by the MTT assay. Of note, solvent (70% ethanol) did not influence the assays (not shown).

Osthole induces apoptotic cell death in TE671 and RK33 cells. Enrichment of oligonucleosome fragments (fold increase) in the absence (Control) and following 24 h of osthole (100 μM) exposure (n=9 per concentration from three independent experiments, ***p<0.001 versus control, Student's t-test).
The mechanism responsible for inhibition of cancer cell growth mediated by osthole is not fully-resolved. An increase of apoptosis was demonstrated after incubation with osthole in HeLa cells (33), as well as a slowdown of the cell-cycle progression at G2/M phase in A549 lung adenocarcinoma cells (32). Osthole was effective in decreasing the migration and invasion of breast cancer cells probably through inhibition of matrix metalloproteinase-2 (MMP2) promoter and MMP2 enzyme activity (36) or hepatocyte growth factor (HGF)-induced decrease of E-cadherin and increase of vimentin expressions (37). More detailed mechanism of action of osthole was linked to up-regulation of the ratio of Bax/Bcl-2 proteins in a lung cancer line (33) and inhibition of Akt kinase activity in lung (33) and breast cancer cells (34). The influence of osthole on the HGF/Met pathway associated with MCF-7 breast cancer cells metastasis and invasiveness was also suggested (37). In our study, we also focused on the potential mechanism of anticancer activity of osthole. To assess if decrease in cell proliferation was connected to apoptosis induction and/or alterations of cell cycle progression, tumor cells were exposed to the selected concentrations of osthole for 24 h. We showed that after treatment with 100 μM of osthole the apoptosis rate in RK33 and TE671 cells increased by 1.5-fold and 2.5-fold, respectively, as measured by mono- and oligonucleosomes release to cytosol (Figure 2).
We also determined that osthole influences cell-cycle progression. FACS analysis showed an increased amount of TE671 cells entering the G1 phase of the cell cycle upon incubation with osthole in a concentration-dependent manner, being mirrored by a decrease in the S/G2 cell population (Figure 3A) (Table I). Minor changes were noticed in the RK33 cell line, however, in contrast to the TE671 cells, treatment with osthole resulted in a slight increase of cell amount in S and G2 phases of the cell cycle (Figure 3B) (Table I).
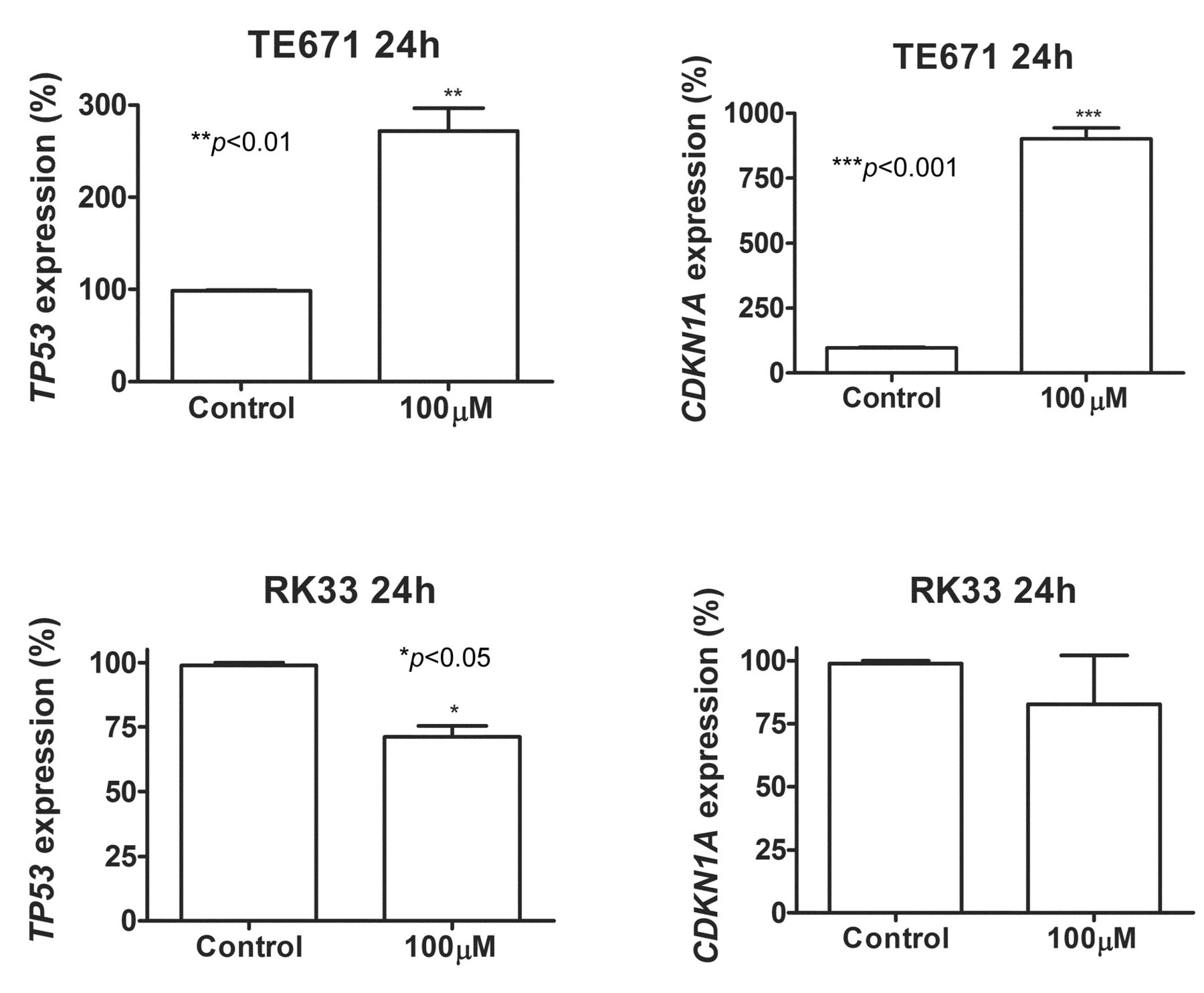
Next, we investigated whether osthole treatment influences gene expression involved in the control of cell-cycle progression. We analyzed the expression of TP53 and CDKN1A in TE671 and RK33 cells. After a 24-h incubation of TE67 cells with osthole, qPCR revealed increased expression of p53 and p21waf1/cip1 (TP53 and CDKN1A) mRNAs by 2.5- and 9-fold, respectively. Such effect was not observed in RK33 cells (Figure 4).
Discussion
Our results indicate that the cellular and molecular mechanism of osthole's anticancer action remains different in the analyzed cell lines. Although osthole inhibited cell proliferation at a similar level in the sudied cell lines, it differentially affected induction of apoptosis and cell-cycle progression of TE671 and RK33 cells. The influence of osthole on TE671 cells could be connected with cell-cycle arrest in the restriction point G1/S, thus slowing-down the cell cycle progression by increased expression of TP53 and CDKN1A coding genes, followed or accompanied by apoptosis. This scheme could not be applied for RK33 cells, where osthole exerts its anticancer mode of action by an unknown mechanism. It has been shown, that osthole down-regulated the expressions of cyclin B1 and p-Cdc2, thereby inducing G2/M arrest in human lung cancer A549 cells (33) and G1/S arrest of cell cycle in vascular smooth muscle cells (38). Together with our data, it seems that the influence of osthole on cell-cycle progression depends on the examined cell type. However, how the mechanism of action of osthole is connected to or dependent on cancer cell type is not known. Coumarin interactions with surface receptors have been suggested (1), therefore the different mode of action of osthole could be related to the type and amount of specific receptors present on different cancer cells. It was found that osthole could abrogate HGF-induced cell scattering, migration and invasion in MCF-7 breast cancer cells (37), thereby osthole blocked the growth factor–induced effect, which supports this hypothesis. However, this issue requires further studies.

Flow cytometric analysis of propidium iodide-stained nuclei revealed higher percentages of (A) TE671 cells in G1 and lower percentages of these cells in S and G2 phases, and (B) lower percentages of RK33 cells in S, and higher in G2 phases of the cell cycle following exposure to 100 μM concentration of osthole compared to controls (*p<0.05, **p<0.01, ***p<0.001, Student's t-test, n=9 per concentration from three independent experiments).
Cell-cycle analysis of TE671 and RK33 treatment by osthole.
It was shown that osthole is a more cytotoxic substance in comparison to other coumarins against the cervical cancer HeLa line, lymphocytic lymphoma HL-60 line and colon cancer CoLo line (27), thereby could be used as the basic compound for new drug development. The modifications of osthole structure led to synthesisof many different osthole derivatives with enhanced anticancer activity (1, 30, 39-41), including a new type of drugs containing osthole and inhibitors of histone deacetylases (HDAC) in one molecule, which were more selective towards histone deacetylase (HDAC) than other HDAC inhibitors (39-41).
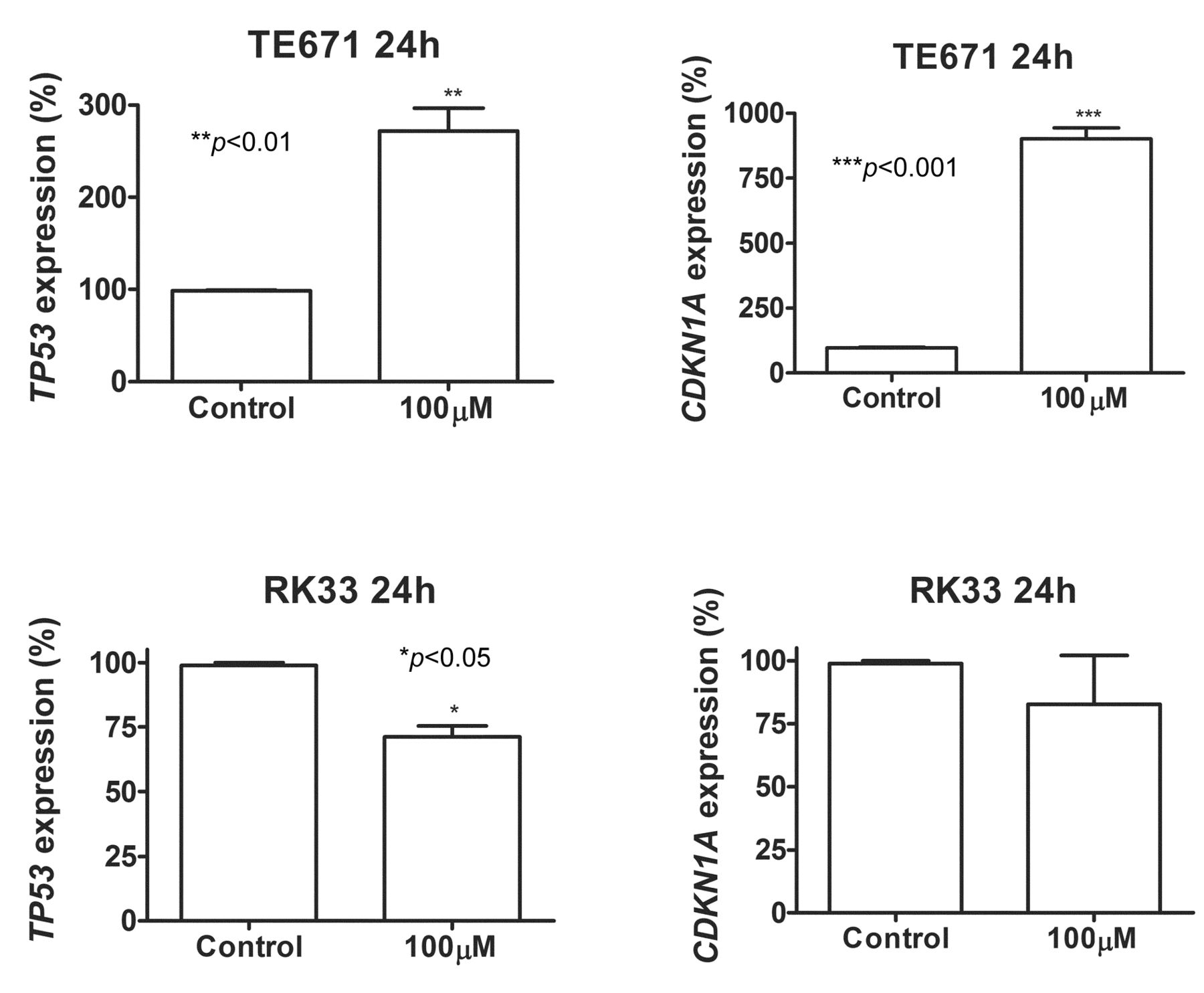
Osthole induces changes in gene expression in cancer cells. Quantification of TP53 and CDKN1A gene expression by means of qPCR method inTE671 and RK33 cells exposed (24 h) to 100 μM concentration of osthole compared to controls (*p<0.05, **p<0.01, ***p<0.001, n=9 measurements from 3 separate experiments, Student's t-test).
In contrast to other chemotherapeutics, coumarin derivatives are relatively low toxic for normal cells. It was demonstrated that osthole was less toxic against normal fibroblast cultures isolated from cervix than against HeLa cervical cancer cells (33). Osthole was also proved to have quite low cytotoxicity against other normal cells such as osteoblasts, where harmful concentrations exceeded 670 μM (15). The concentrations used in our experiments (up to 100 μM) were relatively low, especially when compared with other studies (32) but still able to elicit significant anti-cancer effect.
Conclusion
HPCCC was used for fast separation of minor osthole from the crude extract of the herb Mutellina purpurea. The purified compound was examined for nhibition of cell proliferation and viability, cell cycle progression and induction of apoptosis in human laryngeal cancer RK33 and human medulloblastoma TE671 cell lines. Together with additional beneficial features of osthole treatment, including inhibition of bone resorption (37), protection against liver damage (20, 22) and neuroprotective activities (42), we suggest that osthole may have a therapeutic application in the treatment or chemoprevention of human cancers.
Acknowledgements
This study was supported by a Medical University of Lublin DS440/2012-2013 grant and partially supported by grant no N N405 617538 from the National Science Centre in Krakow, Poland.
- Received July 4, 2014.
- Revision received August 4, 2014.
- Accepted August 6, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Synthetic Betulin Derivatives Inhibit Growth of Glioma Cells In Vitro
- Combination of Osthole and Cisplatin Against Rhabdomyosarcoma TE671 Cells Yielded Additive Pharmacologic Interaction by Means of Isobolographic Analysis
- Additive Interaction of Cisplatin and Histone Deacetylase Inhibitors Combined Treatment in Rhabdomyosarcoma Cells - An Isobolographic Analysis