


Abstract
Background/Aim: Activated Wnt signaling in cancer cells leads to cell proliferation. It has been shown that the Wnt pathway is activated in pancreatic adenocarcinoma cells. Therefore, we tested the effect of Wnt inhibitors in human and murine pancreatic cancer cell lines. Materials and Methods: The Wnt inhibitors ethacrynic acid (EA), ciclopirox olamine (CIC), piroctone olamine (PO) and griseofulvin (GF) were tested in murine and human pancreatic cancer cell lines with the 3-(4,5-dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Results: We showed that the Wnt inhibitors significantly reduced cell viability in murine, as well as human pancreatic cancer cell lines. Conclusion: These results may lead to a new therapeutic option with Wnt inhibitors for patients with pancreatic adenocarcinoma.
Pancreatic cancer is still a lethal disease with a five-year relative survival rate of 5.6% in the United States (1999-2006). In 2008, pancreatic cancer belonged to one of the ten leading cancer-related causes of death worldwide (1).
The pancreas is divided in an exocrine and endocrine part. Most pancreatic cancers occur in the exocrine part and a common type is the adenocarcinoma. Problematically, this cancer is often diagnosed at an advanced stage of disease and consequently it has a poor prognosis. Therefore, new therapies are urgently needed.
Our group already showed some approaches of immunotherapy that might be useful in the treatment of pancreatic cancer (2-4).
Moreover, aberrant Wnt/β-catenin activation has been revealed in pancreatic adenocarcinoma (5). Thus, it is of great interest to develop new therapies targeting the Wnt/β-catenin pathway to treat this type of cancer.
Several reviews describe the canonical Wnt pathway in detail. In general, Wnt signaling regulates cell differentiation and proliferation (6, 7). In healthy cells the canonical signaling pathway is inactivated, whereby β-catenin is degraded. In the cytosol a destruction complex is formed consisting of the adenomatous polyposis coli (APC), axin, casein kinase 1 and glycogen synthase kinase 3 (GSK3β) proteins. With help of this destruction complex, β-catenin can be phosphorylated and is ready for ubiquitination by β-transducin repeat-containing protein (β-TrCP). Finally β-catenin can be degraded by a proteasome (7, 8).
Upon Wnt signaling, Wnt is able to bind on Frizzled and low density lipoprotein receptor-related protein 5 (LRP5) and LRP6 receptors. This signal leads to an inhibition of the degradation of β-catenin. Subsequently, β-catenin accumulates in the nucleus, where it interacts with the transcriptional factors T-cell factor/lymphoid enhancing factor (TCF/LEF) (7, 9). As a result, Wnt target genes like c-myc and cyclin D1 are transcribed (10, 11).
In this study we tested the effect of the Wnt-inhibitors ethacrynic acid (EA) (12), ciclopirox olamin (CIC) (13), piroctone olamine (PO) (14) and griseofulvin (GF) (15) on the viability of a murine and a human pancreatic cell line (PancO2 and DanG, respectively).
Materials and Methods
Cell lines and culture conditions. The human pancreatic cancer cell line DanG (DSMZ, Braunschweig, Germany) and the murine pancreatic cancer cell line PancO2 (kindly provided by Dr Guillermo Mazzolini, University of Navarra, Pamplona, Spain) were incubated at 37°C, 5%CO2, 90% humidity and cultured in RPMI-1640 medium (PAN Biotech, Aidenbach, Germany) complemented with 10% FCS (Invitrogen, Darmstadt, Germany) and 1% penicillin/streptomycin (Invitrogen, Darmstadt, Germany).
Drugs and chemical reagents. In our experiments we used following reagents: griseofulvin (Sigma Aldrich, Taufkirchen, Germany), piroctone olamine (Spinnrad, Bonn, Germany), ciclopirox olamine (Sigma-Aldrich) and ethacrynic Acid (Sigma-Aldrich). The agents were dissolved in dimethyl sulfoxide (DMSO; Carl Roth, Karlsruhe, Germany) or dimethylformamide (DMF; Sigma-Aldrich) at different concentrations.
MTT assay. Cell viability was measured with 3-(4, 5-dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide (MTT) (Sigma-Aldrich). When MTT is added in metabolically active cells, it is metabolized into a water-insoluble dye. The amount of this dye is proportional to the number of viable cells. After solving the dye with an organic solvent, the absorption can be measured at 560 nm.
The cells were seeded in 96-well plates with 1×104 cells/well in 100 μl medium in duplicates or triplicates. The agent was added in a range of concentrations. One triplet without any agent was provided for each cell line as a negative control. A well with cycloheximide (AppliChem, Darmstadt, Germany) served as a positive control. About 45 min before the end of the incubation time the medium was removed and 55 μl MTT (5 mg/ml solved in phosphate buffered saline (PBS; PAA, Pasching, Austria)) were added to each well. After an incubation time of 45 min to 4 h, the MTT was removed and 100 μl of DMSO were added to each well. The plates were shaken at 350 rpm until the cells were lysed and the absorption was measured at 560 nm.
A triplicate of medium controls without any cells and agent was measured once and used for the final evaluation.
Statistical analysis. For statistical analysis the relative viability was shown as the mean±standard deviation (SD) and the student's t-test was applied. Differences of two values were considered as significant when the p-value was less than 0.05. Statistical analysis was performed with the GraphPad Prism software, version 5.0 (GraphPad Software, La Jolla, CA, USA).
Results
Effect of DMF/DMSO on cell viability. DMSO increased the cell viability in DanG cells to 106.5±18.3%, while in PancO2 cells the cell viability was reduced to 95.6±10.3%. When DanG cells were exposed to DMF the cell viability was reduced to 93.6±18.4% and that of PancO2 cells was decreased to 93.6±14.1% (Figure 1).
Effect of CIC on cell viability. The effect of EA, CIC, GF and PO on the human pancreatic cancer cell line DanG and the murine pancreatic cancer cell line PancO2 was tested. Our results showed that EA, CIC, GF and PO significantly reduced the viability of human and murine pancreatic cell lines (Figures 2, 3, 4 and 5). In particular, CIC reduced cell viability even at low concentrations. CIC had a toxic effect towards the cells in the range of 1 μM-10 μM (Figure 3).
Effect of EA on cell viability. The effect of EA was different in both cell lines (Figure 5). Therefore, a final conclusion on the effect of EA on DanG cells cannot be drawn.
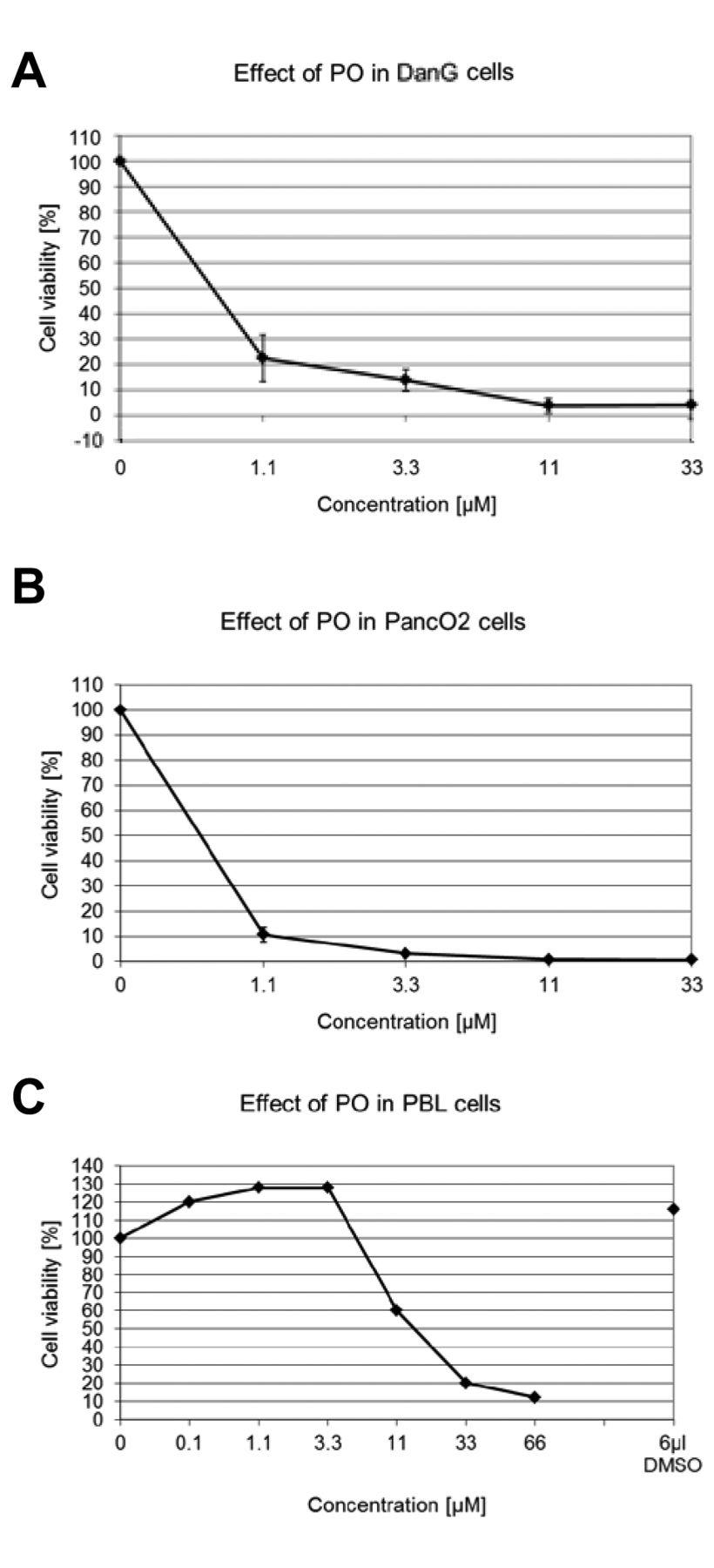
Effect of PO on cell viability. PO reduced the cell viability when used at low concentrations. PO showed a toxic effect even at a concentration less than 1.1 μM (Figure 2).
Effect of GF on cell viability. GF was cytotoxic at concentrations between 10 μM and 50 μM (Figure 4). The determined IC50 values of CIC, EA, PO and GF are shown in Table I.
Discussion
Wnt-signaling plays an important role in the development of different organs (16, 17) and, as of recently, in metastasis (18). It is suggested that this pathway is also important for the development of the pancreas (19, 20).
Mutations involved in the Wnt-pathway lead to aberrant Wnt signaling (21-23). Since such mutations are not common in pancreatic adenocarcinoma cells (23, 24), several groups have detected an abnormal increase of β-catenin in these cancer cells (5, 25, 26). In addition, di Maligano et al. proved that important molecules within the Wnt pathway like Frizzled and Wnt were also up-regulated (25).
In absence of a Wnt signal, β-catenin can be phosphorylated by a destruction complex consisting of APC protein, cytoplasmic serine/threonine kinase GSK-3β, casein kinase 1 and axin. The phosphorylation initiates a cascade ending with the degradation of β-catenin (7, 8). As a result, this essential molecule of the Wnt pathway is not able to accumulate in the cytoplasm and translocate into the nucleus. Upon Wnt-signaling, β-catenin cannot be degraded and therefore de-phoshorylated β-catenin accumulates in the nucleus, where it interacts with the transcription factors TCF/LEF (9) and target genes, like c-myc and cyclinD1, are transcribed (10, 11).
Inhibition of the Wnt pathway results in decreased cell proliferation and apoptosis. The aim of our study was to evaluate the effect of different Wnt inhibitors on the viability of pancreatic adenocarcinoma cells.
EA is a diuretic agent that has been shown to induce cell death in different cancer cells (13, 27, 28). In addition EA has been shown to reduce tumor growth in vivo and prolong overall survival in mice treated with myeloma cells (29).
Aizawa et al. showed that EA is able to increase the level of glutathione (GSH) but when they examined whether this was the reason for its cytotoxic effect, they found that EA was able to induce cell death in a colon cancer cell line, independently of the GSH level. Moreover, this group showed that EA was able to alter the mitogen-activated protein kinase (MAPK) signaling (30).
Since Lu et al. demonstrated that EA could interact with LEF-1, which results in a destabilization of the LEF-1/β-catenin complex (12), another mechanism through which EA might be able to reduce cell viability is revealed.
CIC, GF and PO are pyridone derivatives with anti-fungal effects. They have also been proven to be cytotoxic to myeloma and lymphoma cells (14, 15, 31, 32).
CIC can affect the cell in different ways; it can function as an iron chelator (33) being thus able to inhibit ribonucleotide reductase resulting in cell death (34). This might be one of the mechanisms of CIC action as an anticancer drug. In addition, CIC has been shown to slow down cell cycle progression between the G1/G0- and the S-phase (35). Also, Zhou et al. showed that, upon treatment with CIC, the concentrations of some cyclin and cyclin-dependent kinases (CDK), responsible for the regulation of cell-cycle progression, were down-regulated. The level of p21Cip1 was up-regulated, suggesting that this might lead to an accumulation of cancer cells in the G1/G0 cell cycle. Moreover, the same group demonstrated that apoptosis in cancer cells exposed to CIC may be due to a decrease in the concentration of antiapoptotic proteins (Bcl-xL, survivin) and an increased cleavage of Bcl-2.
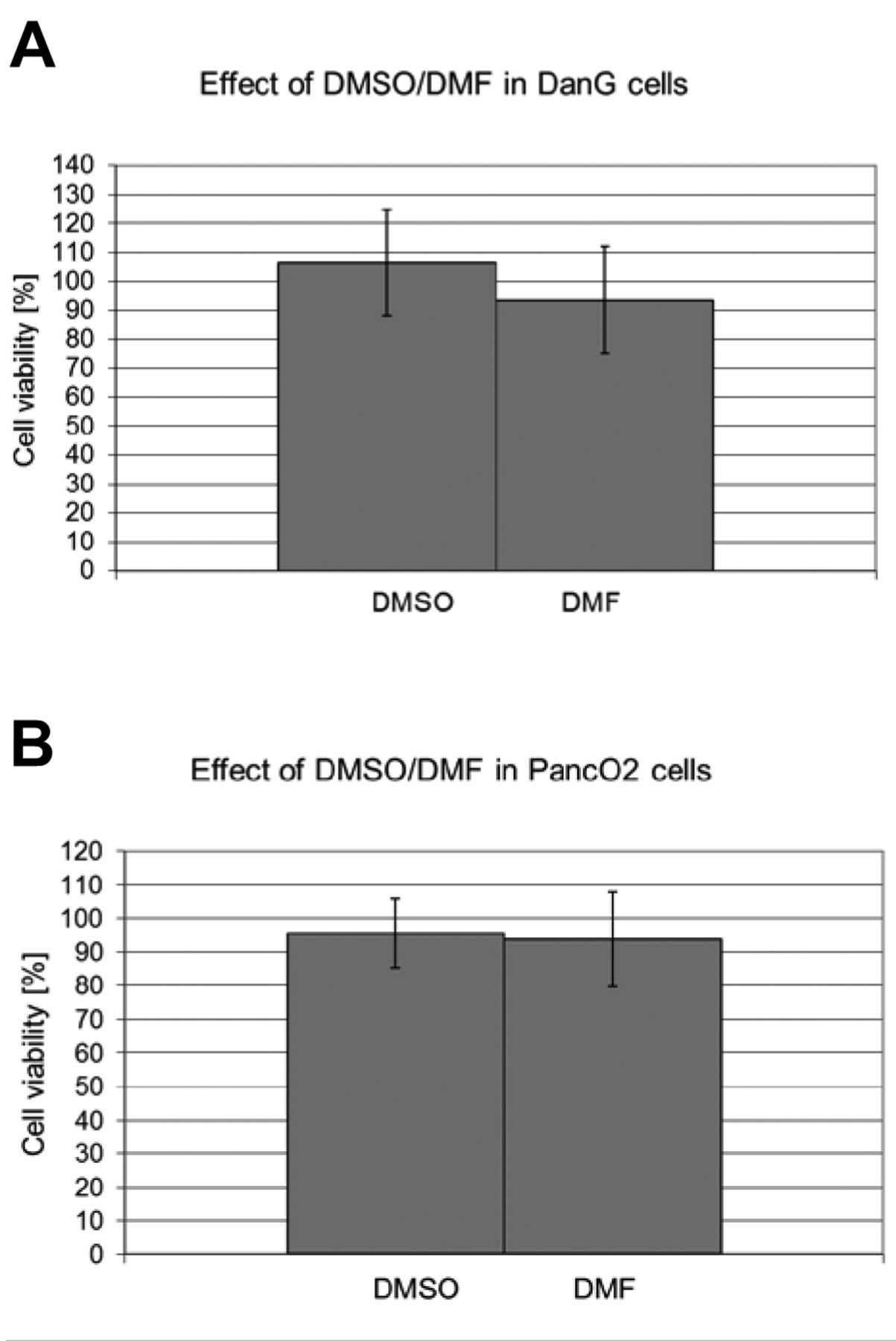
Effect of dimethyl sulfoxide/dimethylformamide (DMSO/DMF) on viability of (A) DanG and (B) PancO2.
The effect of griseofulvin (GF), ciclopirox olamine (CIC), ethacrynic acid (EA) and pirocton olamine (PO) on the pancreatic adenocarcinoma cell lines DanG and PancO2 was tested. The agents were added at various concentrations for three days. Cell viability was measured using the MTT assay and the IC50 value was determined in μM. n=6-11 experiments.

Effect of piroctone olamine (PO) on viability of (A) DanG (n=6) and (B) PancO2 (n=6). 1×104 cells were cultured for 72h with various concentrations of ciclopirox olamine (CIC). Viability was measured with the MTT assay. Data are shown as mean±SD. (C) Experiments with PBL (n=2) were generated as described in the Materials and Methods section.
In addition, Kim et al. showed that β-catenin was down-regulated when CIC was added in lymphoma cells (31), while a 96-well plate-based TOPflash reporter system identified CIC as a Wnt/β-catenin inhibitor (13).

Effect of ciclopirox olamine (CIC) on viability of (A) DanG (n=11) and (B) PancO2 (n=6). 1×104 cells were cultured for 72 h with various concentrations of ciclopirox olamine (CIC). Viability was measured with the MTT assay. Data are shown as mean±SD. (C) Experiments with PBL (n=2) were generated as described in the Materials and Methods section.
Schmeel et al. reported that, in the murine myeloma cell line MPC11, the LEF-1 level was slightly down-regulated when exposed to CIC (36).

Effect of griseofulvin (GF) on viability of (A) DanG (n=6) and (B) PancO2 (n=9). 1×104 cells were cultured for 72 h with various concentrations of ciclopirox olamine (CIC). Viability was measured with the MTT assay. Data are shown as mean±SD. (C) Experiments with PBL (n=2) were generated as described in the Materials and Methods section.
Interestingly, several groups have revealed a role of iron in the β-catenin-dependent transcription. For example, it has been shown that a decrease of iron correlates with an attenuated pathway (37). Another group has shown that an iron chelator was able to suppress the Wnt/β-catenin signaling by inhibition of the transcription of β-catenin target genes. Thus, they suggested that iron containing proteins take part in β-catenin dependent transcriptions in a tissue-correlated way (38).

Effect of ethacrynic acid (EA) on viability of (A) DanG (n=9) and (B) PancO2 (n=6). 1x104 cells were cultured for 72 h with various concentrations of ciclopirox olamine (CIC). Viability was measured with the MTT assay. Data are shown as mean±SD. (C) Experiments with PBL (n=2) were generated as described in the Materials and Methods section.
PO is commonly used in cosmetic products and also this agent is able to inhibit energy metabolism in mitochondria by forming complexes with iron ions (14).
Since GF and PO have similar chemical features compared to CIC, these agents have been tested on lymphoma and myeloma cell lines and it is suggested that they are able to inhibit the Wnt pathway as well (14, 15). In accordance with other studies, we showed that these agents are able to reduce the viability of cancer cells. The fact that these agents are cytotoxic towards pancreatic adenocarcinoma cells in vitro calls for more studies, including experiments in vivo, to further determine the value of these promising findings.
Acknowledgements
The Authors we kindly recognize the support of Sabine Blaum-Feder and Clara Jäkel, Bonn, Germany.
- Received June 26, 2014.
- Revision received July 30, 2014.
- Accepted July 31, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- In Vitro Apoptosis Induction by Fenofibrate in Lymphoma and Multiple Myeloma
- Griseofulvin Efficiently Induces Apoptosis in In Vitro Treatment of Lymphoma and Multiple Myeloma
- Clofibrate Demonstrates Efficacy in In Vitro Treatment of Lymphoma and Multiple Myeloma
- Integrated Genomic Analysis of Pancreatic Ductal Adenocarcinomas Reveals Genomic Rearrangement Events as Significant Drivers of Disease
- In Vitro Efficacy of Naftifine Against Lymphoma and Multiple Myeloma
- Flunarizine Exhibits In Vitro Efficacy Against Lymphoma and Multiple Myeloma Cells
- In Vitro Efficacy of Cinnarizine Against Lymphoma and Multiple Myeloma