


Abstract
Aim: The aim of the present study was to evaluate the effects of a diet supplementation with either olive oil, or n-3 or n-6-polyunsaturatedFatty acids (PUFAs) on tumour development and gene expression for lipogenic enzymes in ApcMin/+ mice. Materials and Methods: In the control group, the mice received a standard diet, the OO group was fed on a diet with 12% olive oil, the OM-3 group with 12% salmon fish rich in n-3 PUFAs, the OM-6 group with 12% oenothera oil rich in n-6 PUFAs. Gene expression of lipogenic enzymes was evaluated by real-time reverse transcription polymerase chain reaction. Results: All mice in the treated groups presented a reduction in total intestinal polyp number and load, which was particularly marked in the OM-3 group. Treated mice showed an induction of low density lipoprotein receptor gene expression and a significant reduction of expression of lipogenic gene. Conclusion: Our data provide new insights into the mechanism of cell growth inhibition and apoptotic regulation by dietary olive oil and PUFAs in ApcMin/+ mice.
Lipid metabolism in cancer cells is regulated by the common oncogenic signaling pathways and is believed to be important for the initiation and progression of tumors (1, 2). Alterations in the mevalonate pathway, the main metabolic way used by proliferating cells to gain lipids for cell membrane synthesis, are common in neoplasia (3). The activity of 3-hydroxy-3-methyl-glutaryl CoA reductase (HMGCoAR) is up-regulated by several fold in colonic tumors and does not respond to the inhibitory feedback induced by cholesterol (4).
Cancer cells seem to require an increased concentration of cholesterol and cholesterol precursors and this requirement may be fulfilled by increased HMGCoAR activity. In our previous study, HMGCoAR activity was found to be enhanced in human colorectal cancer that did not express low density lipoprotein receptor (LDL-R), indicating that LDL-R absence, which deprives colonic neoplastic cells of exogenous sterols, is overcome by an increase in endogenous cholesterol synthesis (5).
These observations suggest that the LDL-R has a role in cell growth and tumorigenesis. In this connection, interest has been attributed on factors regulating LDL-R expression. Previously, we demonstrated the efficacy of eicosapentaenoic acid (EPA), belonging to the of omega-3 family polyunsaturated fatty acids (PUFAs), and lovastatin, a potent inhibitor of HMGCoAR, in gene expression regulation of HMGCoA reductase and LDL-R in the HepG2 cell line (6).
Several studies have revealed that PUFAs can regulate the expression of genes involved in several metabolic pathways (7, 8). Feeding with fish oil, rich in n-3 PUFAs, drastically reduced mRNA levels of genes for lipogenic enzymes in rodent liver (9). PUFAs may control cholesterogenic gene expression through their effects on (SREBP)-dependent regulation (7). SREBP-responsive genes include those coding for HMGCoAR and for intermediates in cholesterol synthesis, such as farnesyl pirophosphate synthase (FPPS). PUFAs reduced mRNA levels of lipogenic enzymes in rat hepatoma cells and mouse liver, in correlation with their effects on HMGCoAR (9).
In addition, dietary n-3-PUFAs can reduce tumor load in mice with an (APC) defect (10). There is also clinical evidence that these fatty acids have specific effects on disease prevention, reducing rectal polyp number and size in patients with familial adenomatous polyposis (FAP) (11).
A recent clinical study demonstrated that adherence to a Mediterranean diet reduces the risk of colorectal cancer (12). Another study shows that a Mediterranean diet, supplemented with extra-virgin olive oil or nuts, reduces the incidence of major cardiovascular events (13). The healthy effects of olive oil can be attributed not only to the higher ratio of unsaturated to saturated fatty acids, but also to the antioxidant property of its phenolic compounds, such as oleuropein (OL) and hydroxytyrosol (HT) (14-16). In fact, we have previously demonstrated that HT and OL, the main polyphenols contained in olive oil, have antiproliferative effects and induce apoptosis of human colorectal cancer cells by suppressing fatty acid synthase activity (17). Moreover, we have recently shown that HT causes an evident antiproliferative effect in a dose- and time-dependent manner on two human hepatoma cell lines, HepG2 and Hep3B. This antiproliferative effect was mediated by down regulation of lipogenic enzymes, activation of the cell antioxidant system, and reduction of cellular (IL-6) levels (18).
Taking this background into account, the present study was designed to evaluate the effects of a diet supplementation with either olive oil, or n-3-PUFAs, or n-6-PUFAs in the ApcMin/+ mouse model, an experimental model that closely mimics the mechanisms of APC gene inactivation observed in patients with FAP and in most sporadic colon adenomas. Moreover, the study explores the molecular mechanisms and gene modulation involved in these dietary effects.
Materials and Methods
Animals and experimental study design. Five-week-old C57BL/6J mice with an heterozygote mutation for the Apc gene (ApcMin/+) and wild-type mice were obtained from Charles River (Calco, CO, Italy). Mice were maintained in temperature, air- and light-controlled conditions and received food and water ad libitum. Animals did not receive any surgical or hormonal manipulation. All animals received care in compliance with the “Guide for the Care and Use of Laboratory Animals”. The procedures related to animal use have been communicated to the Italian Ministry of Health and approved (art. 7 DL 116/92, RC2011).
ApcMin/+ mice were randomly divided into four groups fed for 10 weeks with isocaloric diets in which the fat source in each was different: Control group, n=10 animals received a standard diet (12.5% protein, 12% soya bean oil, 3% fiber); OO group, n=10 animals received a standard diet in which soya bean oil was substituted with olive oil (12.5% protein, 12% olive oil, 3% fiber); OM-3 group, n=10 animals fed a diet in which soya bean oil was substituted with 12% of salmon fish rich in n-3 PUFAs; OM-6 group, n=10 animals fed a diet in which soya bean oil was substituted with 12% of oenothera oil rich in n-6 PUFAs. To control laboratory conditions, wild-type syngenic mice (n=5) were fed the standard diet for 10 weeks and killed for molecular analyses as described hereafter.
All diets were provided in pellet form (Mucedola Srl, Settimo Milanese, Italy) and immediately divided into daily aliquots and stored at −80°C to prevent PUFA oxidation. Mice were provided with fresh food daily, and body weights were recorded weekly.
After 10 weeks of dietary treatment, animals were killed by cervical dislocation and the entire intestinal tract was immediately removed and washed with cold phosphate-buffered saline (PBS). The small bowel was divided into three segments: proximal, medial and distal intestine, and all segments of the small intestine and colon were cut along the mesenteric insertion, placed on a paper strip at 0-4°C and analyzed by a stereomicroscope in order to evaluate the number, location and size of polyps, calculated considering them as hemispheres (1/2×3/4 π r3). Fresh tissue samples of intestine and colon were collected and stored at −80°C until assayed.
Sequences of amplification primers.
Lipogenic gene expression analysis and apoptotic death assay. To study the effects of the diets on gene expression of lipogenic enzymes and of apoptosis in the intestinal distal tract from treated mice, the mRNA levels of LDL-R, FPPS and HMGCoAR genes, as well as the levels of (Bax) and (Bcl2) were assessed by real-time (RT-PCR). The reactions were performed using a master mix with SYBR Green (iQ SYBR Green Supermix, Bio-Rad, Milan, Italy) and sense and antisense primers for the target genes and the β-actin gene (Table I). Real-time PCR was carried out in an CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Inc.) using the following protocol: 45 cycles at 95°C for 3 min, 95°C for 10 s, 55°C for 30 s followed by a melting curve step at 65-95°C at a heating rate of 0.5°C per cycle for 80 cycles. The PCR products were quantified by external calibration curves, one for each tested gene, obtained with serial dilutions of known copy number of molecules (102-107 molecules). All expression data were normalized by dividing the target amount by the amount of β-actin used as internal control for each sample. The specificity of PCR products were confirmed by gel electrophoresis.
Statistical analysis. The significance of the differences among experimental groups was evaluated with one-way analysis of variance (ANOVA) and Tukey's multiple comparison test. Differences were considered significant at a 5% probability level. The correlation analysis between LDL-R and HMGCoAR mRNA levels was performed by nonparametric correlation test (Spearman correlation).
Results
The body weights of ApcMin/+ mice and wild-type mice did not differ significantly during the study and the final body weights were 22.5±3.1 g and 25±1.5 g, respectively. Average daily food intake of ApcMin/+ mice and wild-type mice did not differ significantly.

A: Number of polyps in the whole intestinal tract from ApcMin/+ mice treated groups: ST, Standard diet; OO, standard diet supplemented with olive oil; OM-3, standard diet supplemented with n-3 PUFAs; OM-6, standard diet supplemented with n-6-PUFAs. Data are expressed as the mean±SE. *p<0.05 vs. ST group (one-way analysis of variance and Tukey's multiple comparison test). §p<0.05 vs. OO and OM-6 groups (Tukey's multiple comparison test). B: Polyp volume (mm3) calculated for all polyps in the whole intestinal tract from mice. Data are expressed as the mean±SE. *p<0.05 vs. ST group (one-way analysis of variance and Tukey's multiple comparison test).
After 10 weeks of dietary treatment, none of the animals fed with experimental diets showed any observable toxicity or any gross changes attributable to liver, kidney or lung toxicity.
Figure 1A shows that the diet enriched with n-3 PUFAs induced a statistically significant reduction in total polyp number along the entire intestinal tract compared with the mice fed the standard diet and with mice treated with olive oil and n-6-PUFAs. As far as polyp volume is concerned, all the dietary-treated groups presented a significant reduction, this being particularly marked in the OM-3 group when compared with mice fed the standard diet (Figure 1B).
Gene expression. Figure 2 shows LDL-R, FPPS and HMGCoAR mRNA levels in the distal intestinal mucosa of wild-type mice. The levels of gene expression detected in wild-type mice are comparable to those detected in ApcMin/+ mice fed on the standard diet.
All dietary-treated groups presented an induction of LDL-R gene expression when compared with ApcMin/+ mice fed on the standard diet. The up-regulation was significantly evident in ApcMin/+ mice treated with n-3-PUFAs (Figure 3). All dietary treatments caused a reduction of FPPS gene expression levels. In particular, diets containing olive oil, as well as n-6 PUFAs had a significant effect on the reduction of FPPS mRNA (p<0.05 by Tukey's multiple comparison test) (Figure 3). For HMGCoAR expression, olive oil treatment elicited a statistically significant reduction of mRNA levels for these enzymes when compared to the group with standard diet (p<0.05 by Tukey's multiple comparison test).
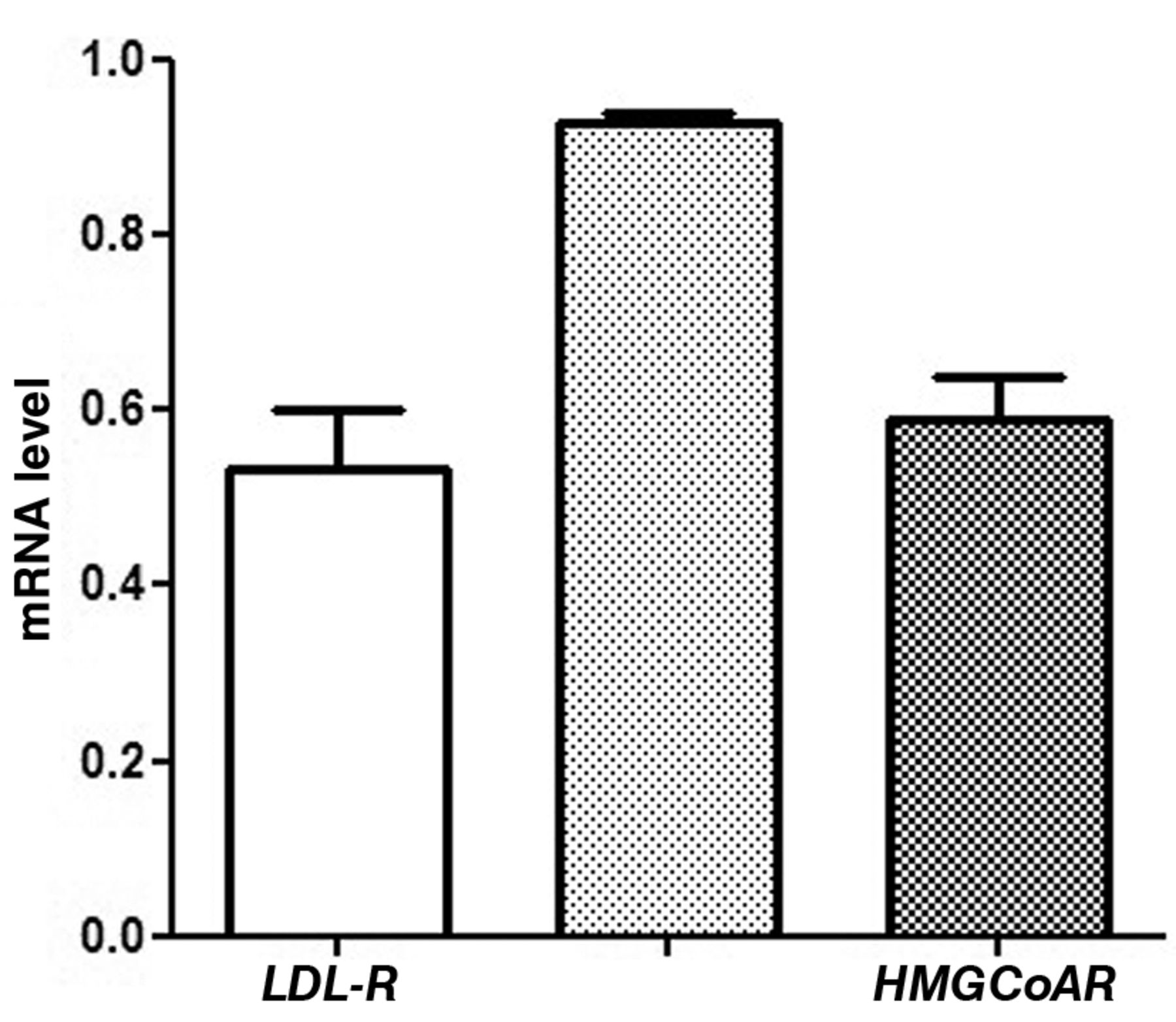
LDL-R, FPPS and HMGCoAR mRNA levels (mean±SE) in distal intestinal mucosa of wild-type mice. Data are expressed as no. molecules mRNA target gene/no molecules mRNA β-actin.
In mice treated with a diet enriched with n-3 PUFAs, a strong negative correlation was observed between HMGCoAR and LDL-R gene expression (Spearman r=−0.9, p-value=0.08) (Figure 4).
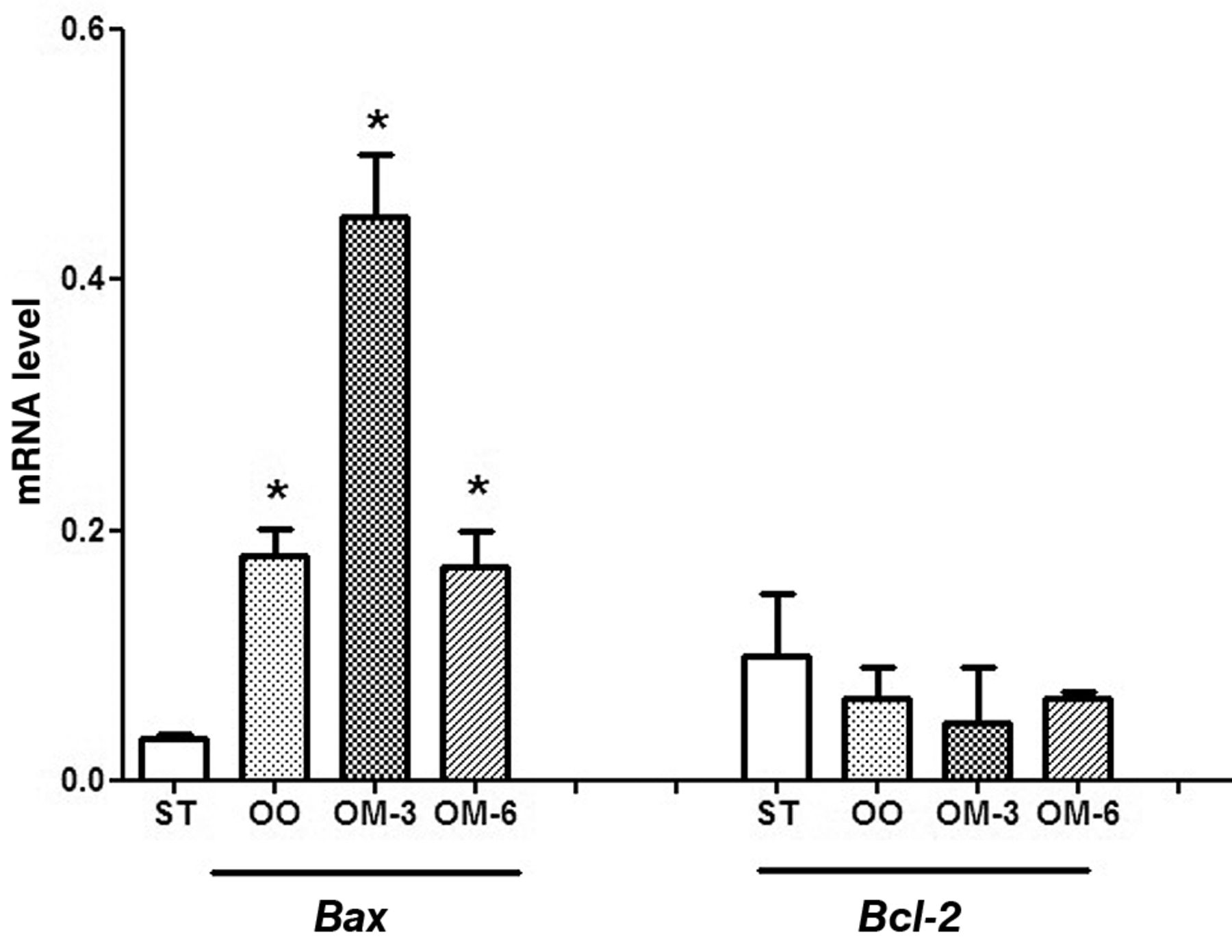
To examine the effects of dietary treatment on apoptosis, Bax and Bcl2 gene expression were evaluated in intestinal distal tract samples. We observed a striking significant increase of Bax mRNA levels, especially in animals receiving n-3 PUFA, associated with a modest decrease of Bcl2 mRNA levels, compared to the control group (Figure 5).
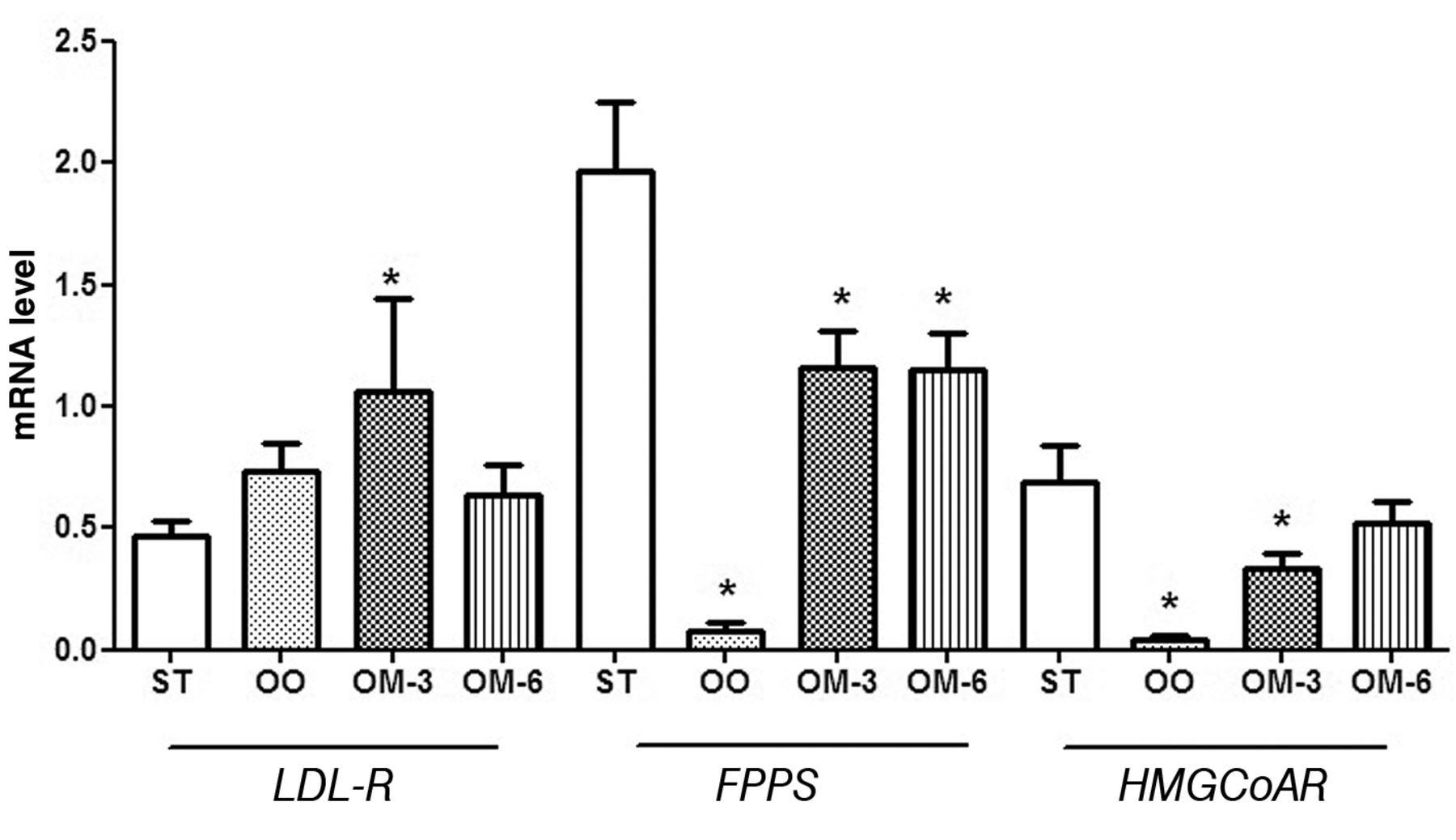
LDL-R, FPPS and HMGCoAR mRNA levels (mean±SE) in distal intestinal mucosa from ApcMin/+ mice treated groups: ST, Standard diet; OO, standard diet supplemented with olive oil; OM-3, standard diet supplemented with n-3 PUFAs; OM-6, standard diet supplemented with n-6 PUFAs. Data are expressed as no. molecules mRNA target gene/no. molecules mRNA β-actin. *p<0.05 vs. ST group (one-way analysis of variance and Tukey's multiple comparison test).
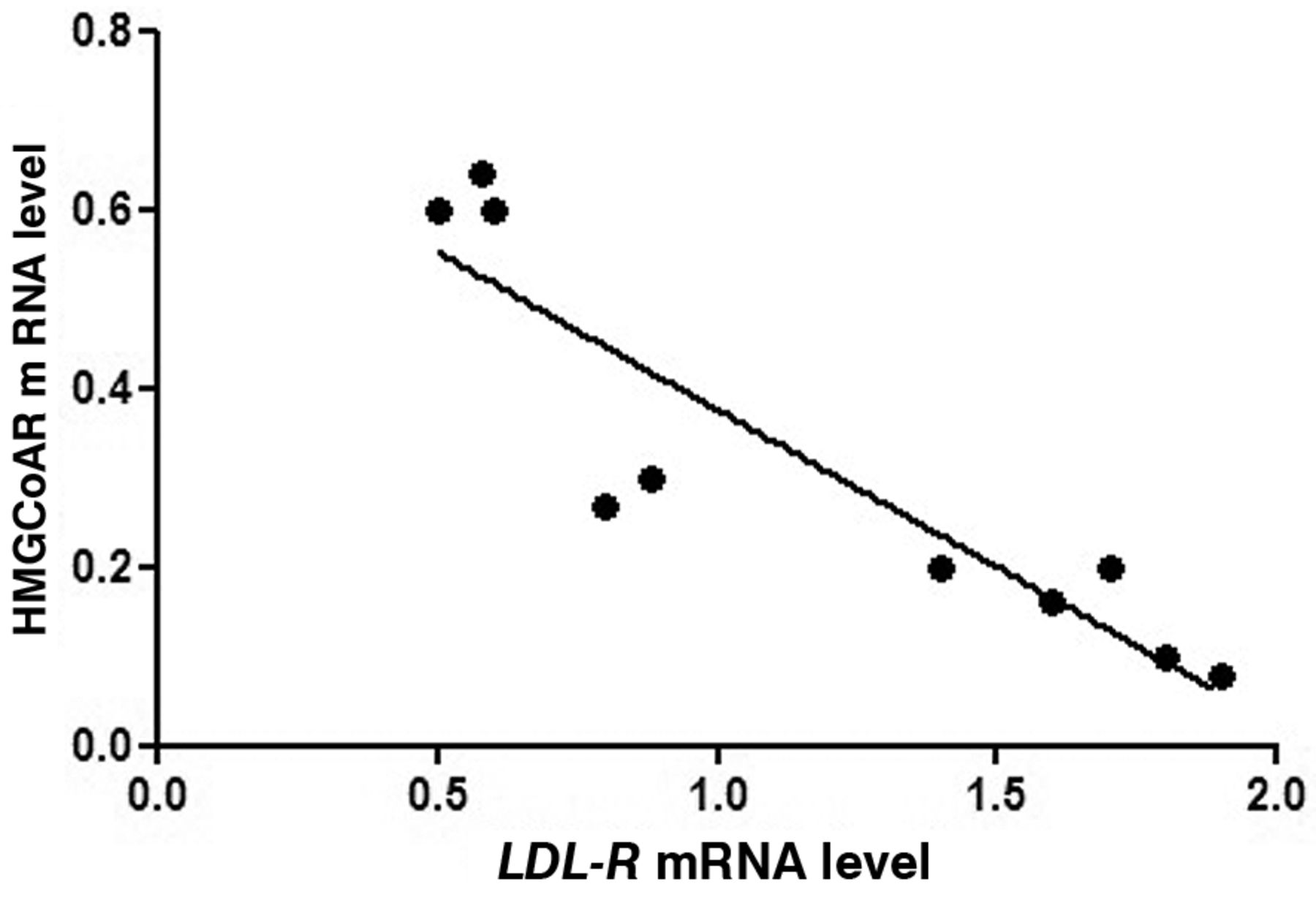
Spearman correlation between HMGCoAR and LDL-R mRNA levels in ApcMin/+ mice from OM-3 group fed with standard diet supplemented with n-3 PUFAs (Spearman r=−0.9, p-value=0.08).
Discussion
Our data confirm a role for increased gene expression of LDL-R and for reduced gene expression of FPPS and HMGCoAR as a partial explanation for the antitumor effect of natural dietary treatments on ApcMin/+ mice. Moreover, it appears that enhanced apoptosis, after n-3-PUFA administration, may be responsible for preventing increased tumor number and load in these mice.
An important feature of colon malignant transformation of the colon is the loss of the cholesterol feedback inhibition mechanism that regulates cholesterol synthesis. The down-regulation of the LDL-R expression in neoplastic cells makes tumors dependent on endogenous cholesterol synthesis for their proliferation, resulting in an increase of HMGCoA reductase activity in the cells and consequently in an increase of cholesterol and mevalonate-derived metabolites, such as isoprenoid compounds.

Bax and Bcl2 mRNA levels (mean±SE) in distal intestinal mucosa from ApcMin/+ mice for four groups of treatment (ST, standard diet; OO, standard diet supplemented with olive oil; OM-3, standard diet supplemented with n-3 PUFAs; OM-6, standard diet supplemented with n-6 PUFAs). Data are expressed as no. molecules mRNA target gene/no. molecules mRNA β-actin. *p<0.05 vs. ST group (one-way analysis of variance and Tukey's multiple comparison test).
Several studies have elucidated the role of isoprenoid and the cholesterol biosynthetic pathway in colonic tumorigenesis (19-21). Inhibitors of the enzymes of the mevalonate pathway are considered to be potential antitumor agents because of their ability to inhibit cellular growth and proliferation (22). Our previous study demonstrated that exposure of DLD-1 cells to a selective inhibitor of FPPS, promoted the apoptotic process, reinforcing the role of isoprenoids in the regulation of cellular metabolic processes (23).
It is well-established that some key dietary components are able to influence cellular dynamics in the colonic epithelium (24). This is noteworthy because the balance between apoptosis and proliferation is critical to the maintenance of steady-state cell populations in the intestine. The promotion of apoptosis is an important aspect of chemoprevention.
In literature there is unequivocal morphological evidence that PUFAs induce apoptosis in vitro (24-26). The mechanism of induction of apoptosis by PUFAs could involve the disruption of membrane fluidity or the involvement of any of the multiple products of PUFA metabolism directly or by releasing a second messenger. In fact, the antitumor effect of dietary n-3-PUFAs detected in this study seems to be associated with enhanced apoptosis and the regulation of lipid metabolism.
For olive oil, the same mechanism can be also hypothesized. It is generally accepted that phenolic compounds of olive oil may exert their cancer-preventive activity by interfering with proliferation and apoptosis of cancer cells (27). Several human and animal studies have shown that olive oil phenols are bioavailable and they are absorbed in the small intestine (28-30). The antiproliferative and proapoptotic effects of olive oil detected in this study are mediated by lipid metabolism modulation.
All natural compounds studied seem to be able to elicit their effects through the down-regulation of HMGCoAR and FPPS gene expression, associated with the induction of LDL-R mRNA, even if this induction is statistically significant only for mice treated with n-3 PUFA.
LDL-R has important implications in the control of cell survival and cell death, having an inhibitory effect on tumor cell growth (5, 6). Moreover, in mice treated with n-3 PUFA, LDL-R and HMGCoAR gene expression appear to be strongly correlated, underlying the importance of dietary treatment in controlling LDL-R up-regulation and HMGCoAR down-regulation.
In conclusion, our data provide new insights into the mechanism of cell growth inhibition and apoptotic regulation by dietary olive oil and PUFAs. These effects associated with lipogenic enzyme expression modifications strongly suggest a causative role for these enzymes in regulating tumor cell growth; therefore, the use of these natural compounds in an anticancer therapeutic strategy should be encouraged.
Acknowledgements
The Authors thank Mr. Vito Spilotro for his excellent technical assistance.
- Received June 18, 2013.
- Revision received June 28, 2013.
- Accepted July 1, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}