


Abstract
15,16-Dihydrotanshinone I (DHTS) is a component of the traditional Chinese medicinal plant Salvia miltiorrhiza Bunge. In this study, DHTS at as low as 2.5 μg/ml concentration significantly inhibited proliferation of human benign (SW480) and malignant (SW620) colorectal cancer cells, as shown by 3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-diphenytetrazoliumromide (MTT) and flow cytometric analysis. Activating transcription factor (ATF)-3, a basic leucine zipper-type transcription factor, was found to be predominantly up-regulated in DHTS-treated SW480 and SW620 cells. The up-regulation of ATF3 was blocked by a c-JUN N-terminal kinase (JNK) or p38 inhibitor. Overexpression of ATF3 resulted in a significant augmentation of DHTS-induced apoptosis of SW480 cells, but resistance to DHTS-induced apoptosis of SW620 cells. These results suggest that DHTS has a strong therapeutic or preventive potential against cancer. In addition, ATF3 has a dual role in DHTS-induced apoptosis, depending on the degree of malignancy of colorectal cancer.
According to a report of annual updates on cancer occurrence and trends in the US, the incidence of colorectal cancers ranked third among both men and women. It is estimated that 143,460 people will be diagnosed with and 51,690 people will die of cancer of the colon and rectum in 2012 in the US (1). The average death rate was 16.7 per 100,000 people per year during 2005-2009 in the US (National Cancer Institute; http://seer.cancer.gov/). Early diagnosis can increase the 5-year relative survival rate to 89.9%, but the survival rate drops to 11.9% when the cancer has metastasized (1). Colorectal cancer incidence and mortality has been increasing rapidly in Taiwan during the last few decades. According to the Taiwan Cancer Registry Annual Report 2009, the incidence and mortality rates of colorectal cancer are 1.4-fold and 1.7-fold, respectively, higher in Taiwan than in the Americas (http://cph.ntu.edu.tw/main.php?Page=N1). Currently, there are several therapeutic methods, including surgery, radiotherapy, chemotherapy, and combinations of these for treatment of colorectal cancer.
Activating transcription factor (ATF)-3 contains a basic region and a leucine zipper (bZIP) motif and belongs to the bZIP superfamily of transcription factors (2). Several transcription factor-binding sites were found within the gene promoter of ATF3, including activating transcription factor/cyclic AMP response element (ATF/CRE), activator protein-1 (AP1), nuclear factor (NF)-κB, v-myc myelocytomatosis viral oncogene homolog (MYC)/MYC associated factor X (Max), and E2F transcription factor (E2F)-binding sites (3). Many signal transduction pathways are also involved in the induction of ATF3, such as mitogen-activated protein kinase (MAPK), NF-κB, ataxia telangiectasia mutated (ATM), nijmegen breakage syndrome protein-1 (NIBRIN), and p53-dependent and p53-independent pathways (4). Research also indicated that ATF3 is a stress-inducible gene. ATF3 is also called liver-regenerating factor (LRF)-1 due to its high expression in the regenerating liver (5). In animal experiments, inducible ATF3 expression was found in conditions of myocardial ischemia, ischemia-reperfusion, hepatic ischemia (especially with a hepatectomy), seizures, and wounds. Treatments with cytokines, genotoxic agents such as UV radiation and ionizing radiation, and drugs also induce ATF3 expression in cell models (6). Microtubule-binding agents, such as taxol and colchicines, induce cell-cycle arrest and apoptosis through ATF3 induction in human MCF-7 breast cancer cells (7). The inhibition of pro-inflammatory tumor necrosis factor (TNF)-α expression by the peroxisome proliferator-activated receptor (PPAR) activator in human umbilical vein endothelial cells was associated with ATF3 induction (8). Although many reports found that ATF3 plays a role in different diseases and conditions, its physiological roles are still unclear. In particular, scientists found a controversial role of ATF3 in tumorigenesis (9). ATF3 acts as a proapoptotic factor that inhibits tumorigenesis of rat sarcoma oncogene (RAS)-transformed mouse embryonic fibroblasts (10). On the other hand, ATF3 serves as an antiapoptotic factor that protects cardiac myocytes from doxorubicin-induced apoptosis (11).
15,16-Dihydrotanshinone I (DHTS) is a component of the traditional Chinese herbal medicine, Salvia miltiorrhiza Bunge (Tanshen), which is used to treat cardiovascular disease, especially angina pectoris and myocardial infarction (12, 13). The bioactive ingredients of S. miltiorrhiza can be classified into two major types: water-soluble phenolic acids and lipophilic tanshinone (14). The lipophilic tanshinone fraction was found to have multiple biological functions, including antibacterial, anti-inflammatory, antioxidant, and anticancer activities (13). DHTS exhibited anti-inflammatory activity by inhibiting nitric oxide (NO) production in mouse BV-2 microglia and RAW264.8 macrophages (15, 16). DHTS also induced hepatoma HepG2 cell apoptosis through induction of reactive oxygen species (ROS) and the p38 pathway (17). Our previous studies demonstrated that DHTS induced cell-cycle arrest at the G1 phase, apoptosis through inhibition of cyclin-dependent kinase (CDK)-2 and CDK4 activities, and activation of the mitochondrial apoptosis pathway (14). In addition, DHTS inhibited prostate cancer cell apoptosis through induction of endoplasmic reticular (ER) stress (18). However, whether DHTS can induce colorectal cancer cell apoptosis and the role of ATF3 in DHTS-induced apoptosis of colorectal cancer cells are unclear.
Materials and Methods
Materials. DHTS was purchased from Xi'an Honson Biotechnology (Xi'an, P. R. China). Its purity was >90% according to high-performance liquid chromatographic (HPLC) analysis. Antibodies against ATF3, glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and α-tubulin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA), and to phospho-extracellular-signal-regulated kinase (ERK), phospho-p38, and phospho-c-Jun N-terminal kinase (JNK) were purchased from Cell Signaling (New England Biolabs, Schwalbach, Germany).
Cell culture and the cell viability assay. SW480 and SW620 cells were purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, ROC), cultured in Leibovitz's L-15 supplemented with 10% heat inactivated fetal bovine serum (FBS), and maintained in a humidified incubator at 37°C without CO2.
Cell viability was determined by an 3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-diphenytetrazoliumromide (MTT) assay as described previously (14).
Flow cytometric analysis. Drug-treated cells were stained with annexin V-Alexa Fluor 488 and propidium iodide (PI), and analyzed by FACScan flow cytometry using the CellQuest 3.3 software (Becton Dickinson, San Jose, CA, USA) as described previously (14).
Western blot analysis. Total cellular proteins (50 μg) were resolved by 8%-12% sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE), transferred onto a polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA), and visualized using enhanced chemiluminescence kits (Amersham, Arlington, IL, USA) as described previously (19).
Transition transfection. An ATF3-overexpressing plasmid, pCI-ATF3, was kindly provided by Professor Shigetaka Kitajima (Tokyo Medical and Dental University, Tokyo, Japan). Cells were seeded in 6-cm dishes and transfected with either pcDNA3, control empty plasmid, or pCI-ATF3 plasmid using Lipofetamine 2000™ (Invitrogen Corp., Grand Island, NY, USA) as described previously (19).
Statistical analysis. Student's t-test was used for the statistical analysis, and differences were considered significant at p<0.05.
Results
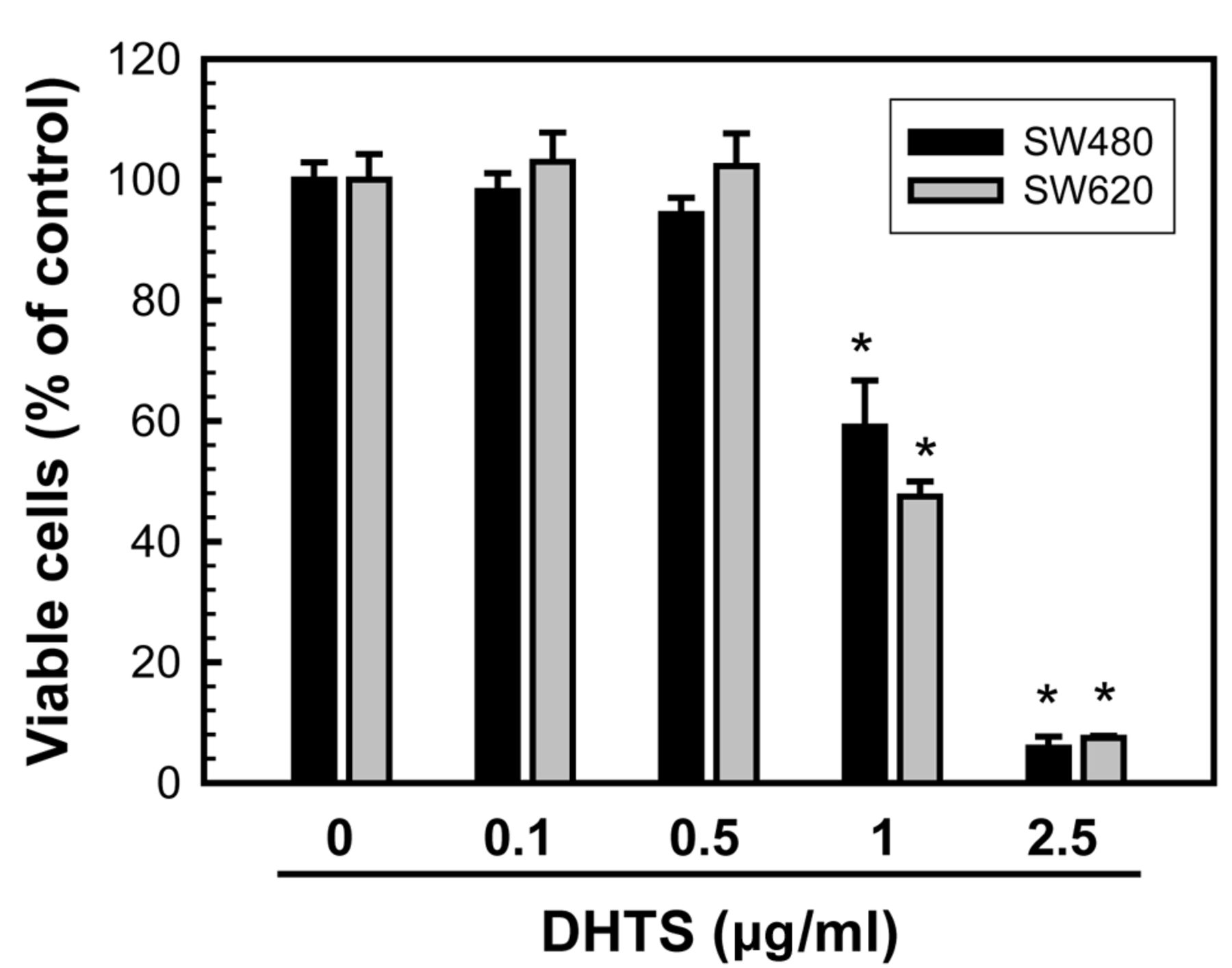
DHTS inhibited cell proliferation and induced cell apoptosis of human colorectal adenocarcinoma cells. To investigate whether DHTS affects the proliferation of human colorectal adenocarcinoma cells, primary colorectal adenocarcinoma SW480 cells and SW480-derived metastatic adenocarcinoma SW620 cells were used in this study. SW480 and SW620 cells were treated with different concentrations of DHTS for 24 h, then the cell number was determined by an MTT assay. As shown in Figure 1, 1 μg/ml DHTS significantly reduced SW480 and SW620 cell numbers by 40.9% and 52.5%, respectively. There was no significant effect on cell numbers at doses of <0.5 μg/ml DHTS for either cell type.
To examine whether DHTS inhibited colorectal adenocarcinoma cell proliferation through inducing cell apoptosis, SW480 and SW620 cells were treated with 0.5, 1.0, or 2.5 μg/ml DHTS for 24 h, and then stained with PI and annexin V-Alexa Fluor 488 followed by quantification of apoptotic cells under flow cytometry. As shown in Figure 2A, 0.5, 1, and 2.5 μg/ml DHTS significantly induced late apoptosis of SW420 cells (upper right) by 7.76%, 20.82%, and 51.27%, respectively. Similar results for DHTS-induced apoptosis were found in SW620 cells (Figure 2B).

Effects of 15,16-dihydrotanshinone I (DHTS) on the viability of human colorectal adenocarcinoma cells. SW480 (A) and SW620 (B) cells were treated with the indicated concentrations of DHTS for 24 h, and viable cells were determined by an 3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-diphenytetrazoliumromide (MTT) assay. Values are presented as the mean±S.E. of four independent experiments performed in quadruplicate. *p<0.05 vs. the control (untreated).
DHTS induced ATF3 expression and activated MAPK pathways. It was found that ATF3 is involved in either proapoptotic or antiapoptotic activities which can be induced by various stresses including anticancer drugs (11). Previous studies also indicated that MAPKs are essential upstream signaling pathways in stress-induced ATF3 expression (20). To investigate whether ATF3 plays a part in DHTS-induced apoptosis and whether MAPKs are involved in DHTS-induced ATF3 expression, SW480 and SW620 cells were treated with DHTS, and the expression of ATF3 and the phosphorylation of ERK, JNK, and p38 were determined by western blotting. As shown in Figure 3A, 0.5-2.5 μg/ml DHTS significantly induced ATF3 protein expression in a dose-dependent manner in both cell types. In addition, 1 μg/ml DHTS time-dependently induced ATF3 expression (Figure 3B). Interestingly, ATF3 expression reached a peak at 24 h in SW480 cells but at 9 h in SW620 cells after DHTS treatment. The phosphorylation of JNK was observed at the earliest measured time point of 3 h in DHTS-treated SW620 cells but at 9 h in DHTS-treated SW480 cells. These results suggest that DHTS indeed induced the expression of ATF3 and phosphorylation of ERK, JNK, and p38 in human colorectal adenocarcinoma cells, but ATF3 expression and JNK phosphorylation occurred at different time points in SW480 and SW620 cells.

Effects of 15,16-dihydrotanshinone I (DHTS) on the induction of apoptosis of human colorectal adenocarcinoma cells. SW480 (A) and SW620 (B) cells were treated with the indicated concentrations of DHTS for 24 h, and apoptotic cells were determined by fluorescence-activated cell sorting (FACS) using annexin V-propidium iodide double-staining. Percentages of early apoptotic (lower right) and late apoptotic (upper right) cells are shown in each panel.

Effects of 15,16-dihydrotanshinone I (DHTS) on the protein expression of activating transcription factor (ATF)-3 and phosphorylation of mitogen-activated protein kinases (MAPKs) in human colorectal adenocarcinoma cells. A: SW480 and SW620 cells were treated with the indicated concentrations of DHTS for 24 h. B: SW480 and SW620 cells were treated with 1 μg/ml DHTS for the indicated time periods. Protein expressions of ATF3, phosphor (p)-extracellular signal-regulated kinase (ERK), p- c-Jun N-terminal kinase (JNK), and p-p38 of total cell extracts were analyzed by western blotting.
Overexpression of ATF3 affected the cell fates of DHTS-treated primary and metastatic colorectal adenocarcinoma cells. To investigate the role of ATF3 in DHTS-induced apoptosis of colorectal cancer cells, SW480 and SW620 cells were transitionally transfected with an ATF3-overexpressing plasmid, pCI-ATF3, and treated with DHTS, and then cell viability was determined by the MTT assay. Western blotting showed that the ATF3 protein expression significantly increased in cells transfected with 0.2-0.4 μg of the pCI-ATF3 expression plasmid, clearly indicating ATF3 overexpression in both SW480 and SW620 cells (Figure 5, right panels). Overexpression of ATF3 significantly increased the number of SW480 cells, but inhibited cell proliferation of SW620 cells (Figure 5A and B, first-paired bar). When cells were treated with DHTS, overexpression of ATF3 resulted in enhancement of apoptosis of SW480 cells, but resistance to apoptosis of SW620 cells (Figure 5A and B, third-paired bar). These results suggest that ATF3 plays an important role in regulating apoptosis depending on the degree of malignancy of the tumors.
Discussion
It was demonstrated that DHTS can inhibit cancer cell proliferation through various pathways, and ATF3 plays a role in regulating cell apoptosis. In this study, we found that at very low concentrations DHTS significantly inhibited colorectal cancer proliferation. DHTS also induced ATF3 expression and activated MAPK pathways, and the expression of ATF3 might be associated with activation of JNK and p38. Overexpression of ATF3 enhanced apoptosis of DHTS-treated SW480 cells and boosted resistance to apoptosis of DHTS-treated SW620 cells. These results suggest that DHTS indeed induces apoptosis, the expression of ATF3, and MAPKs in colorectal cancer cells. The present study provides crucial evidence to support the involvement of ATF3 in the induction of apoptosis by DHTS in human colorectal cancer cells.
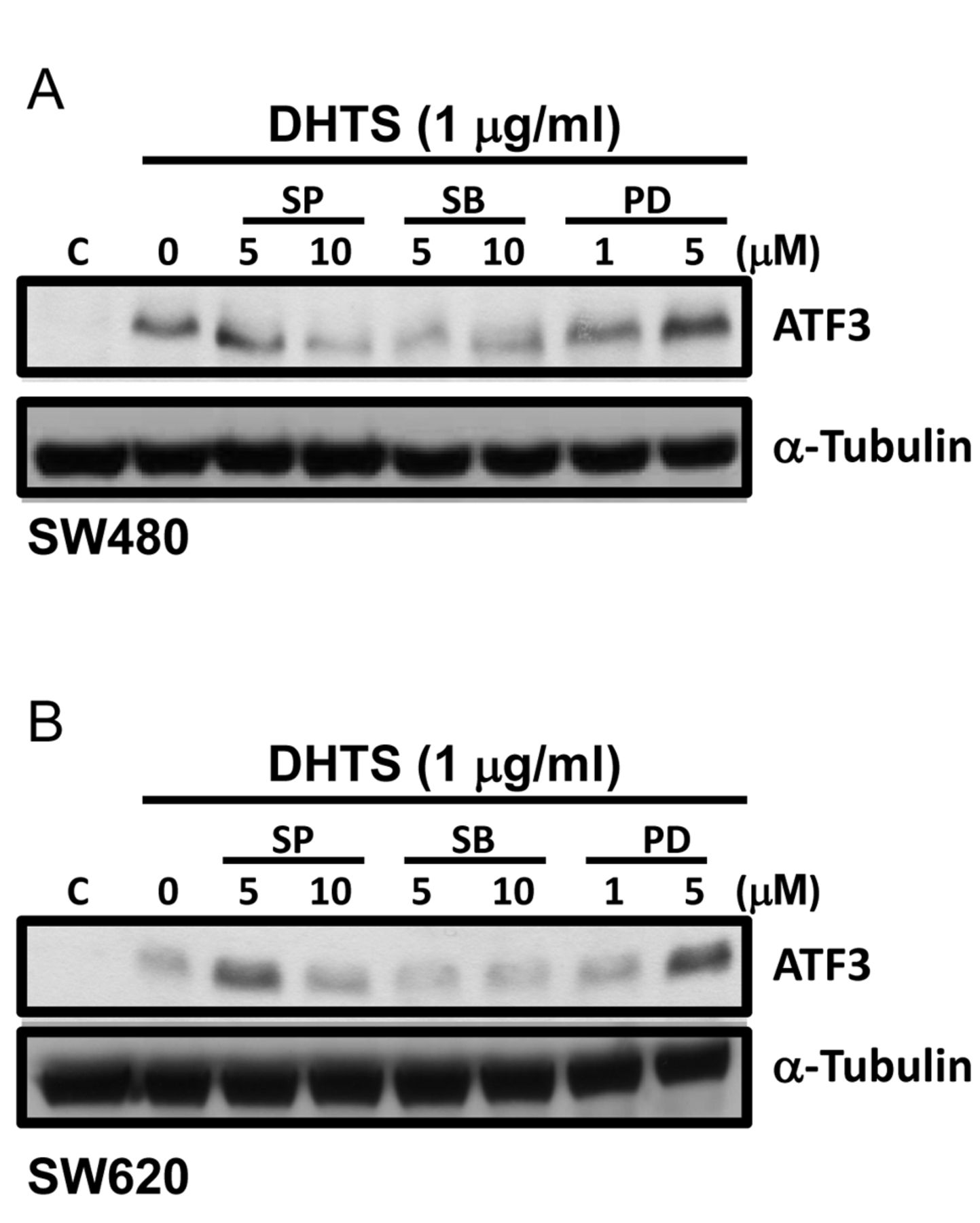
Effects of mitogen-activated protein kinase (MAPK) inhibitors on activating transcription factor (ATF)-3 expression in 15,16-dihydrotanshinone I (DHTS)-treated human colorectal adenocarcinoma cells. SW480 (A) and SW620 (B) cells were pre-treated with the c-Jun N-terminal kinase (JNK) inhibitor, SP600125 (SP), the p38 inhibitor, SB203580 (SB), and the extracellular signal-regulated kinase (ERK) inhibitor, PD98059 (PD), for 1 h followed by 24 and 9 h of co-treatment with 1 μg/ml DHTS in SW480 and SW620 cells, respectively. Protein expression of ATF3 in total cell extracts was analyzed by western blotting.
MAPK pathways form a complex kinase network that is involved in multiple physiological processes including cell proliferation, differentiation, and apoptosis. Three major members of MAPKs were identified: ERK, JNK, and p38 (20). The ERK pathway is well-known to be associated with cell proliferation, cell growth, and survival and is mainly activated by mitogens and growth factors. However, the JNK and p38 pathways can mediate apoptosis in response to environmental stresses or drug treatment (21). Interestingly, sustained and prolonged JNK activation can mediate apoptosis, whereas early JNK activation promotes cell survival (22). Evidence also demonstrated that the stress-activated JNK and p38 pathways can negatively regulate ERK pathway activity, and result in apoptosis induction (21). Our results showed that DHTS significantly increased the phosphorylation of JNK for long periods of time, and induced p38 phosphorylation. Together, these activations might contribute to DHTS-induced apoptosis. Although DHTS induced the ERK growth signal, activated JNK and p38 could negatively-regulate ERK pathway activity, and ultimately result in apoptosis. In addition, activated JNK and p38 might mediate ATF3 induction in DHTS-treated cells, since the JNK inhibitor, SP600125, and the p38 inhibitor, SB203580, blocked ATF3 expression (Figure 4).
Previous studies have demonstrated that NF-κB is an antiapoptotic factor and is expressed at a high level by colorectal cancer cells (23). Inhibition of cyclooxygenase (COX)-2 prevents proliferation of colorectal cancer cells (24). However, DHTS did not inhibit NF-κB activation or COX2 expression of SW480 or SW620 cells (data not shown), suggesting that DHTS inhibited the proliferation of colorectal cancer cells through a mechanism other than inhibition of NF-κB and COX2.
ATF3 induction was linked to drug-induced apoptosis of cancer cells, such as after treatment with curcumin (25), progesterone (26), and a COX inhibitor (27). In this study, we first found that DHTS increased ATF3 expression in dose- and time-dependent manners in both non-malignant SW480 and malignant SW620 colorectal cancer cells. Interestingly, we found that ATF3 had opposite effects on non-malignant SW480 cells and malignant SW620 cells. Similarly, another study indicated that ATF3 had opposite effects on untransformed and high-grade malignant breast cancer cells (28). Under stress, overexpression of ATF3 was deleterious in both non-malignant colorectal cells and breast cells, but protected against cell apoptosis in both malignant colorectal and breast cells. The controversial role of ATF3 might be associated with the cellular context, cell type, and degree of malignancy (10, 28), but this still needs to be clarified. ATF3 is a transcription factor that can regulate expression of several genes and results in regulation of cell proliferation, apoptosis, and survival. Activated ATF3 may bind to the gene promoter of cyclins, induce cyclin expressions, and then stimulate cell-cycle progression (2, 29). Hypoxia-inducible factor-2a and p15 PCNA-associated factor (PAF) respectively participate in cell apoptosis and DNA repair, and are induced by ATF3 in human epidermal cells subjected to UV radiation (30). In human colorectal cancer HCT-116 cells, the ribotoxic stress agent, anisomycin, induced macrophage-inhibitory cytokine-1 expression through ATF3, and then induced apoptosis (31). In this study, it is still unclear which downstream signals are induced by ATF3 in DHTS-treated colorectal cancer cells.

Effects of overexpression of activating transcription factor (ATF)-3 on apoptosis of 15,16-dihydrotanshinone I (DHTS)-treated human colorectal adenocarcinoma cells. SW480 (A) and SW620 (B) cells were transfected with 0.3 μg of the ATF3-overexpressing plasmid, pCI-ATF3, or the control empty plasmid, pcDNA3, for 36 and 24 h, respectively, and then treated with DHTS for another 24 h. Viable cells were determined by the MTT assay as described in Materials and Methods. Values are presented as the mean±S.E. of quadruplicate tests. *p<0.05 vs. individual pcDNA3-transfected cells. SW480 and SW620 cells were transfected with 0.2-0.4 μg of the ATF3-overexpressing plasmid, pCI-ATF3, for 24 h, and protein expression of ATF3 of total cell extracts was analyzed by western blotting (right panels).
Acknowledgements
This work was supported by grants from New Taipei City Hospital (TPCH99-a3) and Taipei Medical University's Wan Fang Hospital (99TMU-WFH-08).
Footnotes
-
↵* These Authors contributed equally to this work.
- Received June 3, 2013.
- Revision received June 23, 2013.
- Accepted June 25, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.