


Abstract
Background: Oral squamous cell carcinoma (OSCC) development involves complex machinery of genomic and epigenetic regulations including microRNA (miRNA) expression changes. We aimed to analyze the expression patterns of a set of miRNAs in human TNM stage I and II OSCC samples in an autologous normal mucosa-controlled experimental design. Materials and Methods: Forty samples of OSCC and 40 matched normal tissues were evaluated for miR-21, miR-155, miR-191, miR-146a, miR-221 and miR-222 expression in a LightCycler 480(PCR) system. Results: Our results showed significant overexpression of miR-21, miR-155, miR-191 and miR-221 in paired-sample t-test and the sensitivity/specificity of tests were over 90% in the case of miR-21 and miR-155 on the receiver operating characteristic curve (ROC) analysis. Conclusion: Our results underline the role of miR-21 in OSCC and support the possible causal role of miR-155 and miR-221 in oral carcinogenesis. The overexpression of miR-191 is a novel finding in squamous cell carcinoma.
Cancer of the oral cavity is still among the major public health issues with considerable individual and socioeconomic impact worldwide. According to the WHO Globocan cancer statistics, cancer of the oral cavity occupies the 10th place in cancer morbidity for men on both the world statistics chart and the WHO European regional statistics. It has an age-standardized incidence rate of 3.8 on a world scale and 4.3 in Europe considering both sexes. Age-standardized mortality rates are 1.9 (world) and 1.7 (Europe), almost half of the rate of the incidences (1). Squamous cell carcinoma is responsible for approximately 90% of all malignant histopathological entities of the oral cavity (2). In fact, despite the many advancements made in the field of oral cancer prevention, and the development of multimodality treatments, the five-year survival rate for oral squamous cell carcinoma (OSCC) remains almost unchanged for the past two decades. The poor prognosis is mainly due to a low response rate even to current multimodality therapeutic strategies (2-4). Primary OSCC treatment relies on surgery with or without neck dissection, or combined surgery and radiotherapy with or without molecular target therapy. With these advanced combination therapies, 35-55% of patients develop locoregional or metastatic recurrence within two years (4, 5). During the development of OSCC, clinical disease characteristics are driven by genomic changes in a distinct field of the oral mucosa (6-9). This type of cancer is formed after environmentally-induced genomic changes or intrinsic genetic or epigenetic alterations of the mucosal cells. These molecular events are not necessarily followed by protein dysfunctions or structural changes immediately; therefore they cannot be detected from the cellular or tissue phenotype alone (10-13).
In OSCC, the diagnostic procedures and the therapeutic decisions rely on the standard histological evaluation, which has remained unchanged for the past decades and is focused on cellular and tissue phenotypic alterations. Molecular characterization of a lesion is rarely carried out in practice (14, 15). Application of even a limited set of molecular markers in diagnosis could successfully supplement histopathology.
miRNAs are small, non-coding, 18-23 nucleotide-length molecules, chemically very stable and which can be detected by a range of high-throughput detection methods in tissue, serum, plasma, saliva, urine and feces, and which are also retrievable from formalin-fixed paraffin-embedded tissue samples (16, 17). miRNA expressions are modified by broad-spectrum mechanisms that independently or in combination drive the carcinogenic process: Alterations of miRNA expression occur in structural genetic alterations of their chromosomal location: chromosomal aberrations, genetic mutations and polymorphisms of the miRNA genes; and by epigenetic changes: alterations of DNA methylation, acetylation or altered chromosomal re-arrangement (17-20).
The potential of miRNA expression signatures to distinguish between normal and tumour tissues and discriminate between different tissue-specific sub-groups has already been confirmed by numerous wide-scale studies of human tumours, first in chronic lymphoid leukaemia (CLL) (21); subsequently, breast, colorectal, head and neck and oral cavity squamous cell carcinomas have been also found to involve miRNA regulatory alterations (22-27). In OSCC, several miRNAs with potential biological and clinical relevance have been identified and are being explored as diagnostic, prognostic and predictive markers.
Our aim in the current study was to analyze expression alterations of six candidate miRNAs (miR-21, miR-155, miR-191, miR-146a, miR-221 and miR-222) from the cancer field of 40 specimen of clinically early-stage OSCC (TNM I-II) and compare them with those of the normal mucosa from the same patients. These specific miRNAs were selected based upon our previous investigations and literature review.
Materials and Methods
Patients. Forty oral squamous cell carcinoma samples were obtained from patients undergoing primary oral surgery for verified oral cancer at the Department of Dentistry, Oral and Maxillofacial Surgery in University of Pécs Medical School. The study was approved by the Regional Ethics Committee (permission number: 2012/4682 for investigation entitled: miRNA Expression Analysis of Oral Carcinomas) and all patients provided informed consent. Sample collection was carried out only during R0 resections of I-II TNM stage OSCCs and biopsies were taken from the tumour tissues. For normal controls, biopsies were taken from the remote part of the reconstruction flaps. Resected specimens were routinely processed for histopathological evaluation and additional biopsy samples were snap-frozen and stored at −80°C until laboratory preparation.
Tissue sample preparation and miRNA isolation. Tissue disruption and homogenization of the fresh-frozen samples were automated by using a MagNa Lyzer Instrument (Roche, Mannheim, Germany) with MagNa Lyzer Green Beads (Roche) sample tubes. Tissue homogenization was followed by miRNA isolation with a general chemical extraction followed by solid-phase extraction on silica columns provided by the High Pure miRNA isolation kits (Roche) according to the manufacturer's instructions. The quality of the yielded miRNA fraction was checked by spectrophotometry (MaestroNano, Maestrogen Inc., Hsinchu, Taiwan) and miRNA samples of higher than 1.9 optical density at 260/280 nm were used for further reverse transcription.
Reverse transcription. Samples were reverse-transcribed in 5 μl reactions using the miRCURY LNA™ Universal RT MicroRNA PCR, Polyadenylation and cDNA synthesis kit (Exiqon, Vedbaek, Denmark). Reverse transcription reactions contained 2.5 μl nuclease-free water, 0.25 μl enzyme mix, 1 μl reaction buffer and 1 μl template cDNA corrected for 5 ng/μl concentrations. To provide the interplate calibrators, a reverse transcription reaction mix was prepared with the same components in the same amounts and concentrations but the template RNA was substituted with 0.25 μl RNA spike-in template (UniSP6 CP provided with the kit) mixed with 0.75 μl of the 6.7 ng/μl MS2 RNA (Roche). Reverse transcription reactions were carried out in Light Cycler glass capillaries (Roche) in a Light Cycler 2.0 thermocycler (Roche) at 42°C for 60 min ended with heat inactivation of the reverse transcription enzyme mix at 95°C for 5 minutes. Freshly-synthesized cDNA samples were chilled at 4°C and immediately proceeded to PCR reactions.
RT-qPCR. The PCR amplification was performed in a LightCycler® 480 Real-Time PCR System (Roche) in 96 well plates. Plate design contained two internal controls and an interplate calibrator for each investigated miRNA. MiCury human LNA™ PCR primer sets (Exiqon) were obtained as specific primers for hsa-miR-21, hsa-miR-155, hsa-miR-191, hsa-miR-146a, hsa-miR-221 and hsa-miR-222 UniSP6 CP, U6 snRNA and 5S rRNA. All cDNA templates from the reverse transcription reaction all were 80 fold diluted for RT-qPCR analysis. PCR mix contained 8 μl cDNA template 2 μl of primer mix and 10 μl of LC480 SYBR Green I Master Mix from the LC480 SYBR Green I Master kit (Roche). Plate layout design was maintained for triplicates of the samples according to each miRNA, two reference genes, three wells for negative controls and three wells for the interplate calibrators. The PCR protocol included one cycle of polymerase activation for 10 minutes at 95°C, 45 cycles of amplification with 10 s denaturation at 95°C, 1 minute annealing at 60°C, and 5 s extension at 72°C in each cycle and one cycle for melting curve analysis with 10 s denaturation at 95°C, 1 min annealing at 60°C and a melting step at 98°C at 0.2°C/s ramp rate. Each sample was run in triplicate.
Data analysis. Exor 4 LightCycler 480 software was used to determine the crossing point cycle (Cp) values. Outlier results with Cp standard deviation greater than 0.5 were excluded from further analysis. Interplate calibration factors were calculated for each miRNA and references as the difference interplate calibrator's overall average Cp and the actual plate's interplate calibrator average Cp. Finally each miRNA and reference was corrected for the calibration factor by subtracting the calibration factor from each. Both U6 snRNA and the 5S rRNA were tested as potential reference genes and U6 snRNA was selected for normalization. miRNA expression values were normalized and calculated by using relative quantification with the 2−dCp method as ΔCp=Cp miR of interest −Cp U6 snRNA. Relative miRNA expression results from the sample triplicates were averaged.
Statistical analysis. Expression results were analyzed with the paired sample t-test and receiver operating characteristic (ROC) curve analysis by SPSS version 19.0 (SPSS Inc., Chicago, IL, USA) and p-values <0.05 were considered to be statistically significant. All graphs were also created by SPSS 19.0.
Results
The 40 investigated OSCC samples were classified by TNM stage I and II. Tumour location included the lip, tongue, buccal mucosa and the floor of the mouth (Table I). Nearly half of the tumours were of TNM stage I (53%). In men, the majority of the OSCCs were located on the tongue, and in women the most frequent location was the lip. The average age of the OSCC patients was 63.8 years and the sex distribution was 7:1 male: female ratio.

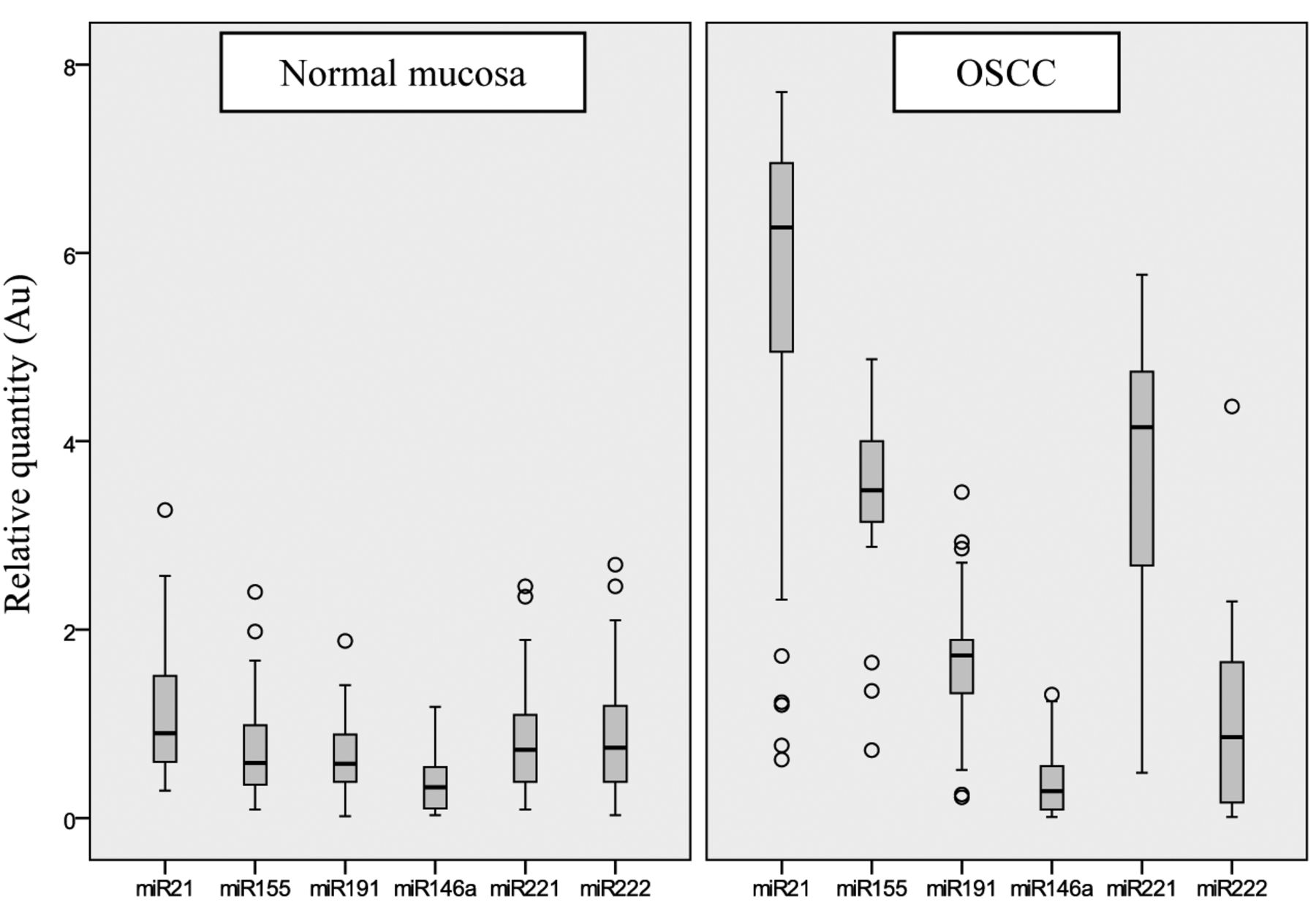
Expressions of miR-21, miR-155, miR-191, miR-146a, miR-221 and miR-222 in oral squamous cell carcinoma (OSCC) and autologous normal mucosa. Relative quantification values were calculated according to the second derivate maximum method and normalized to U6 snRNA expression. AU: Arbitrary units.
All investigated miRNAs were overexpressed in the OSCC samples compared to their expression in their adjacent normal tissues. The most abundantly expressed miRNAs were miR-21, miR-221 and miR-155 in the tumours and miR-146a was found to be expressed at very low amounts (Figure 1). The most characteristic difference and clearest segregation of OSCC versus normal controls by expression, were found in the case of miR-155. Expression values of miR-21 and miR-221 exhibited 4- to 5-fold elevation in the OSCC samples compared to normal tissue, although there was a relatively higher scatter in their distribution. Lower elevation of expression was seen in the case of miR-191 and no observable difference was found for miR-222 and miR-146a. Outstanding variant expressions in both OSCCs and normal tissues are indicated in Figure 1 by tumour sample number. Although there were only a few cases, most were detected among tumour expressions of miR-21 and miR-155.
Comparison of OSCC tumour samples with their autologous normal mucosal pair from the same patient paired sample t-test analysis revealed four of the analyzed miRNAs miR-21, miR-155, miR-191 and miR-221 to have statistically significant expression changes in the tumours (Table II).
ROC curve analysis is presented in Figure 2 and Table III. Testing accuracy of the miR expressions is represented as the area under the curve, which was found to be 90% for miR-221 and higher than 90% in case of miR-21, miR-155.
Oral squamous cell carcinoma samples according to tumour number, location and TNM stage.
Discussion
According to recent knowledge, approximately 30-50 miRNAs have been linked to OSCC based on their altered expression related to this histopathology found by multiple, independent in vitro and in vivo experiments and human studies (26-33). However, only a few have already been proven to be causally involved in carcinogenesis of OSCC (34). For causal correlation to OSCC development, miRNAs need to fulfil the following suggested requirements: i) molecular structural alteration leading to their altered expression in OSCC (DNA structural aberration or epigenetic modification); ii) manipulation of the miRNA gene in an in vitro system leads to verifiable phenotypic alteration; iii) develpomental involvement in OSCC; iv) at least one cellular cancer gene as its target (34). To correspond with current conceptual statements and to provide comparable but partly novel data from our autologous normal tissue-controlled experimental design, miR-21 which satisfies the causality criteria for carcinogenesis of OSCC and another five unconfirmed miRNAs (miR-146a, miR-155, miR-191, miR-221 and miR-222) were the focus of the analysis. Although miR-146a, miR155, miR-191, miR-221 and the miR-222 have not yet been found to be causal in OSCC, they do belong to the group of miRNAs that we still do not have enough information about, but previous research findings support their promising relevance in head, neck and oral cancers (35-37). All miRNAs in our selection belong to the oncomir functional class based on their role in target mRNA regulation.
Paired-sample two-tailed t-test of miRNA expression differences between tumour and normal tissues presenting p-values and 95% confidence intervals for each investigated miRNA (miR-21, miR-155, miR-191, miR-146a, miR-221 and miR-222).
Statistical analysis for the receiver operating characteristic (ROC) curve.
miR-21 is one of the most extensively studied miRNAs in oral and head and neck squamous cell carcinomas, as well as in many other tumour entities, due to the experimental evidence on its inhibition of multiple tumour suppressor targets such as phosphatase and tensin homolog deleted on chromosome 10 (PTEN), tumor suppressor gene, tropomyosin-1 (TPM1), and programmed cell death-4 (PDCD4) (38, 39). One of the most crucial signalling pathways contributing to apoptosis evasion in OSCC is the rat sarcoma viral oncogene (RAS)/phosphoinositide 3-kinase (PI-3K)/PTEN/protein kinase B (AKT) signalling cascade on which miR-21 represses tumour suppressive apoptosis-promoting genes (40). Elevated expression of miR-21 strongly correlated with the progression of pre-malignant leucoplakia to invasive oral carcinoma in a study by Reis et al., where miR-21 was found to negatively-regulate PDCD4, a member of the ATP-dependent DNA helicase (DEAD)-box protein family, acting as translational regulator of eukaryotic translation initiation factor-4A and G1 (eIF4A, eIFG1). By preventing eIF4A helicase activity and assembly of the eIF4F complex, PDCD4 activity results in inhibition of cell transformation. 3’ untranslated region (UTR) down-regulation of PDCD4 by miR-21 has been detected in oral cancer cell lines (41). In our current analysis, we found miR-21 to be most markedly up-regulated in the OSCC samples, with high tumour:normal rate of expression and an increase in expression in the overall miRNA pattern of the malignant lesions. This was also one of the three miRNAs in our panel that surpassed the 90% specificity and sensitivity on ROC analysis for the tumour tissues, which indicates miR-21 to be a useful candidate biomarker for cancer detection.
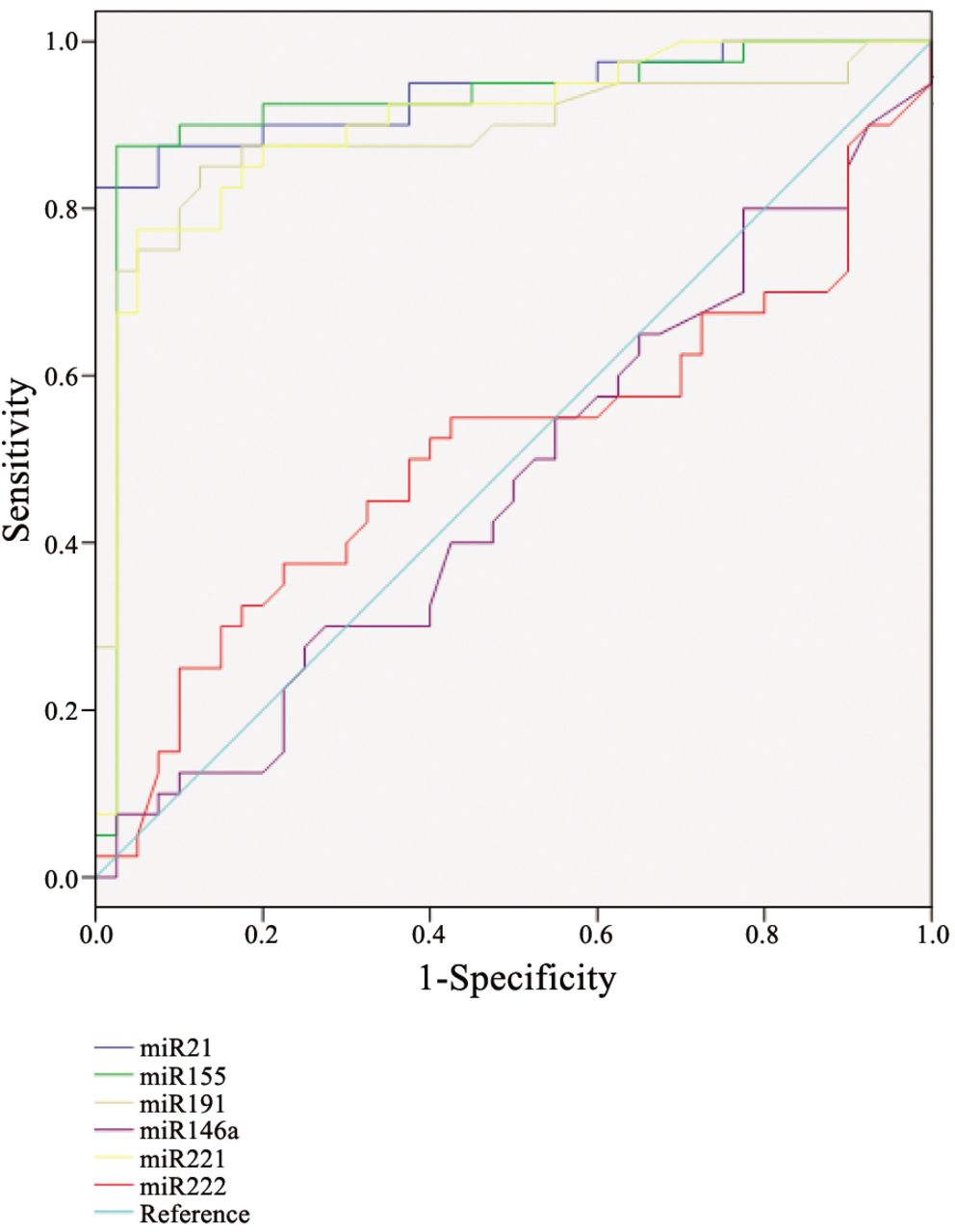
Receiver operating characteristic curve (ROC) analysis.
The role of miR-221 and miR-222 in carcinogenesis was underpinned by several studies (22, 43). miR-221 and miR-222 both act as oncomirs by targeting PTEN and tissue inhibitor of metalloproteinase-3 (TIMP3) tumour suppressors, induce tumour necrosis factor-related apoptosis inducing ligand (TRAIL) resistance and enhance cellular migration through the activation of the AKT pathway and metallopeptidases. MET oncogene was also demonstrated to be involved in miR-221 and miR-222 activation, through the c-JUN transcription factor in different non-small cell lung cancer cell lines (43). miR-221 is known to suppress p27 and p57 transcripts of the retinoblastoma tumour suppressor family (44, 45). Although several studies identified overexpression of miR-221 in head and neck squamous cell carcinomas including cancer of the oral cavity there is a scarcity of reports on functional analysis of these miRNAs in OSCC. Our results demonstrated a characteristic expressional activity of miR-221, confirming its role in oral carcinogenesis while no alterations of miR-222 regulation were observed using our current examinational protocol.
miR-191 has been found to be dysregulated in a large number of different types of human tumours, including colorectal, breast and prostate cancer. Target genes of the mature miRNA sequence have not been characterized, and it is not known which factors lead to its dysregulation in certain tumour cells. The expression profile of miR-191 was suggested to be implemented in prognosis of acute myeloid leukaemia, with higher-than-average levels of miR-191 suggesting a lower survival probability (46). No data confirm any role of miR-191 in OSCC. We observed a slight but statistically significant up-regulation of expression of this miRNA. Our miRNA expression panel included an inflammatory regulator miR-146a of whom predicted targets include nuclear factor of kappa-B (NFκB), interleukin-1 receptor-associated kinase (IRAK1), epidermal growth factor receptor (EGFR), and metastasis-associated protein-2 (MTA2) (47). miR-146a showed no expression changes in our analysis.
miR-155 has also been identified in oral carcinogenesis by some studies that have found miR-155 to be up-regulated in oral cancer compared to normal oral tissue. On the other hand, miR-155 was found to be down-regulated in the presence of human papilloma virus (HPV)-16 in a series of HPV-positive squamous cell carcinoma cell lines of the head and neck (32, 48). Interestingly miR-155 overexpression was confirmed on malignantly-transformed oral leucoplakia and primary human oral cavity tumours, where the tumour location was the tongue (49, 50). miR-155 acts in transforming growth factor (TGF)β-induced epithelial–mesenchymal transition by targeting the Rho family small GTPase RhoA transcript (36). We found miR-155 to be characteristically up-regulated miRNA in our early-stage OSCC samples. It had the highest integrity in the overexpression values; moreover, tumour expressions exhibited the strictest segregation from normal mucosal tissue expression values. ROC curve analysis confirmed statistical significance with similarly high biomarker confidence compared to that for miR-21 in our matched controlled sample set. Comparing our results with the above mentioned studies, it is worth pointing out, that one-third of our OSCC samples (16 out of the 40) were obtained from the tongue and two-thirds from other locations in the oral cavity (bucca, floor of the mouth and the lip). We did not observe any changes or differences in expressions due to tumour location, which could be explained by the relatively low sample numbers representing certain locations.
The significance analysis of microarray and prediction analysis of microarrays from six types of solid tumour (lung, breast, colon, gastric, prostate and endocrine pancreatic tumours) revealed miR-21 and miR-155 to be at the top of the oncomir list (22). With the qRT-PCR results for our focused panel, we achieved miRNA expression results with high reproducibility and minimal genomic background noise due to comparison of the OSCC samples with the adjacent normal mucosal tissues. Our data confirm the role of miR-21, miR-155 and miR-221 in OSCC. Furthermore, based upon our results, functional analysis of miR-155 and miR-221 would be useful to complete the picture of their causal association to OSCC, because they also seem to be very promising genomic biomarkers and potentially useful for early-cancer detection. Our work also provides novel results about the significant de-regulation of miR-191 in OSCC. Providing evidence for the causal nature of a set of miRNAs would open the possibility for their standardized application for prognostic and diagnostic purposes.
- Received February 14, 2013.
- Revision received March 8, 2013.
- Accepted March 12, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- MicroRNA-based cancer mortality risk scoring system and hTERT expression linked with risk-adjusted treatment strategy in early-stage oral squamous cell carcinoma
- Comparative miRNA Expression Profile Analysis of Squamous Cell Carcinoma and Peritumoral Mucosa from the Meso- and Hypopharynx
- Next-generation Sequencing for microRNA Profiling: MicroRNA-21-3p Promotes Oral Cancer Metastasis
- Human and Viral microRNA Expression in Sjogren Syndrome