


Abstract
Background: Naftopidil, an α1-adrenoceptor blocker, has been clinically used for the treatment of benign prostate hyperplasia and hypertension. Emerging evidence has shown that naftopidil exhibits an antitumor effect on a variety of cancer types including prostate cancer. The aim of the present study was to investigate naftopidil-induced apoptosis in human malignant mesothelioma cells and to shed light on the underlying mechanism. Materials and Methods: 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay, terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining, western blotting, and enzymatic assay of caspase-3, -8, and -9 activities were carried out on human malignant mesothelioma cell lines NCI-H28, NCI-H2052, NCI-H2452, and MSTO-211H cells. To knock-down α1D-adrenoceptor, siRNA to silence human α1D-adrenoceptor-targeted gene was constructed and transfected into cells. Results: Naftopidil induced apoptosis in all the investigated malignant mesothelioma cells, and a similar effect was obtained with prazosin, another α1-adrenoceptor blocker. α1-Adrenoceptor is linked to Gq/11 protein involving activation of protein kinase C (PKC). Naftopidil-induced reduction in cell viability was inhibited by GF109203X, while prazosin-induced in cell viability was less affected. Knocking-down α1D-adrenoceptor promoted malignant mesothelioma cell proliferation. Both naftopidil and prazosin activated caspase-3 and -8 in all the investigated malignant mesothelioma cells. Conclusion: Naftopidil, as well as prazosin, has the potential to induce apoptosis in malignant mesothelioma cells by activating caspase-8 and the effector caspase-3, regardless of α1-adrenoceptor blocking.
Adrenaline/noradrenaline play a pivotal role in the autonomic nervous system by activating adrenoceptors, e.g., regulation of cardiac motility and blood pressure. Adrenoceptors are classified into three groups: α1-, α2-, and β-receptors (1-3). α1-Adrenoceptor is linked to Gq/11 protein involving phospholipase C activation, and further divided into α1A-, α1B-, and α1D-subtypes. α2-Adrenoceptor is linked to Gi protein involving adenylate cyclase inhibition, and further divided into α2A-, α2B-, and α2C-subtypes. β-Adrenoceptor is linked to the Gs protein involving adenylate cyclase activation, and further divided into β1-, β2-, β3-, and β4-subtypes.
Intriguingly, α1-adrenoceptor antagonists such as prazosin, doxazosin and terazosin have the potential to inhibit cell growth by arresting the cell cycle or induce apoptosis of malignant and non-malignant cells (4-20). In explanation of the apoptotic action of α1-adrenoceptor antagonists, a variety of pathways have been proposed, such as mitochondria-mediated activation of caspase-3/-9 and c-Jun N-terminal kinase (JNK)1/2; recruitment of Fas-associated death domain (FADD) and the ensuing activation of caspase-8; transforming growth factor-β1 (TGF-β1) activation followed by IκBα induction; or antagonistic effect of BCL-2. α1-Adrenoceptor antagonists, alternatively, exhibit anti-angiogenic effects, resulting in suppression of cell growth in human prostate cancer and human bladder cancer (21-25).
Naftopidil, an antagonist for α1-adrenoceptor, with higher selectivity for α1A- and α1D-receptors, has been developed as a drug for treatment of benign prostate hyperplasia and hypertension (26). Naftopidil is still capable of inhibiting prostate cancer cell growth by arresting cells at the G1 phase of cell cycling (27, 28). Moreover, a study showed that an asymptomatic meningioma markedly regressed two years after oral intake of naftopidil (29). Naftopidil might be available for the treatment of various malignant tumors. Malignant pleural mesothelioma, a highly aggressive neoplasm, has been increasing in incidence and is strongly associated with asbestos exposure (30). No efficient therapy or drug for malignant mesothelioma has been yet established, and therefore, patients with malignant mesothelioma cannot escape death.
The present study investigated the antitumor action of naftopidil on human malignant mesothelioma cell lines. We showed that naftopidil induces apoptosis of malignant mesothelioma cells by a mechanism independent of α1-adrenoceptor blocking.
Materials and Methods
Cell culture. Human malignant pleural mesothelioma cell lines such as NCI-H28, NCI-H2052, NCI-H2452, and MSTO-211H cells were purchased from the American Type Culture Collection (Manassas, VA, USA). Cells were grown in RPMI-1640 medium supplemented with 10% heat-inactivated fetal bovine serum (FBS), 0.003% L-glutamine, penicillin (final concentration, 100 U/ml), and streptomycin (final concentration, 0.1 mg/ml), in a humidified atmosphere of 5% CO2 and 95% air at 37°C.
Cell viability assay. Cell viability was evaluated by the method of 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) as previously described (31).
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining. TUNEL staining was performed to detect in situ DNA fragmentation as a marker of apoptosis using an In Situ Apoptosis Detection Kit (Takara Bio, Otsu, Japan). Briefly, fixed and permeabilized cells were reacted with terminal deoxynucleotidyl transferase and fluorescein isothiocyanate (FITC)-deoxyuridine triphosphate for 90 min at 37°C. FITC signals were visualized with a confocal scanning laser microscope (LSM 510; Carl Zeiss Co., Ltd., Oberkochen, Germany).
Construction and transfection of siRNA. siRNA to silence human α1D-adrenoceptor-targeted gene (α1DR siRNA) and negative control siRNA (NC siRNA) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
The α1DR siRNA or the NC siRNA was reverse-transfected into cells using Lipofectamine reagent (Invitrogen, Carlsbad, CA, USA). Cells were used for experiments 48 h after transfection.
Western blotting. Western blotting was carried out on cells using an antibody to α1D-adrenoceptor (Santa Cruz Biotechnology) and to β-actin (Sigma, St. Louis, MO, USA) by the method previously described (31).
Enzymatic assay of caspase-3, -8, and -9 activities. Caspase activity was measured using a caspase fluorometric assay kit (Ac-Asp-Glu-Val-Asp-MCA for a caspase-3 substrate peptide; Ac-Ile-Glu-Thr-Asp-MCA for a caspase-8 substrate peptide; and Ac-Leu-Glu-His-Asp-MCA for a caspase-9 substrate peptide) by the method described previously (31). Briefly, cells were harvested before and after treatment with naftopidil or prazosin, and then centrifuged at 1,200 rpm for 5 min at 4°C. The cell pellet was incubated on ice in cell lysis buffer for 10 min, and reacted with the fluorescently-labeled tetrapeptide at 37°C for 2 h. The fluorescence was measured at an excitation of wavelength of 380 nm and an emission wavelength of 460 nm with a fluorometer (Fluorescence Spectrometer F-4500; Hitachi High-Tec, Tokyo, Japan).
Statistical analysis. Statistical analysis was carried out using unpaired t-test.
Results
Naftopidil induces apoptosis of malignant mesothelioma cells. For all the malignant mesothelioma cell lines examined here, naftopidil reduced cell viability in a concentration (1-100 μM)- and treatment time (24-48 h)-dependent manner, the extent reaching almost 0% of basal levels at 100 μM (Figure 1A-D). Likewise, prazosin, another α1-adrenoceptor antagonist, reduced cell viability in a fashion mimicking the effect of naftopidil (Figure 1E-H).
In the TUNEL staining, naftopidil significantly increased the number of TUNEL-positive cells as compared with that for the untreated control in all the malignant mesothelioma cell lines (Figure 2A-D). A similar effect was obtained with prazosin (Figure 2A-D). These results indicate that α1-adrenoceptor blockers, such as naftopidil and prazosin, are capable of inducing apoptosis of malignant mesothelioma cells.
Naftopidil induces apoptosis of malignant mesothelioma cells, regardless of α1D-adrenoceptor blocking. α1-Adrenoceptor is linked to Gq/11 protein involving phospholipase C activation, to hydrolyze phosphatidylinositol into inositol 1,4,5-trisphosphate (IP3) and diacyl-glycerol allowing protein kinase C (PKC) activation. We postulated that naftopidil and prazosin induce apoptosis of malignant mesothelioma cells by inhibiting PKC as a result of α1-adrenoceptor blocking. If this was true, then PKC inhibitors should enhance the cell death-inducing effect of naftopidil and prazosin. Unexpectedly, the effect of naftopidil on malignant mesothelioma cell death was attenuated by GF109203X, an inhibitor of PKC, for all the cell types examined here (Figure 3A-D). Naftopidil, thus, appears to induce apoptosis of malignant mesothelioma cells by a mechanism independent of α1-adrenoceptor blocking and PKC inhibition.
GF109203X, in contrast, enhanced the effect of prazosin on cell viability, only for NCI-H28 cells (Figure 3E) or inhibited it for NCI-H2052 and NCI-H2452 cells (Figure 3F,G), but did not alter the effect for MSTO-211H cells (Figure 3H). It is even less likely that prazosin induces apoptosis of malignant mesothelioma cells by inhibiting α1-adrenoceptor bearing PKC activation.
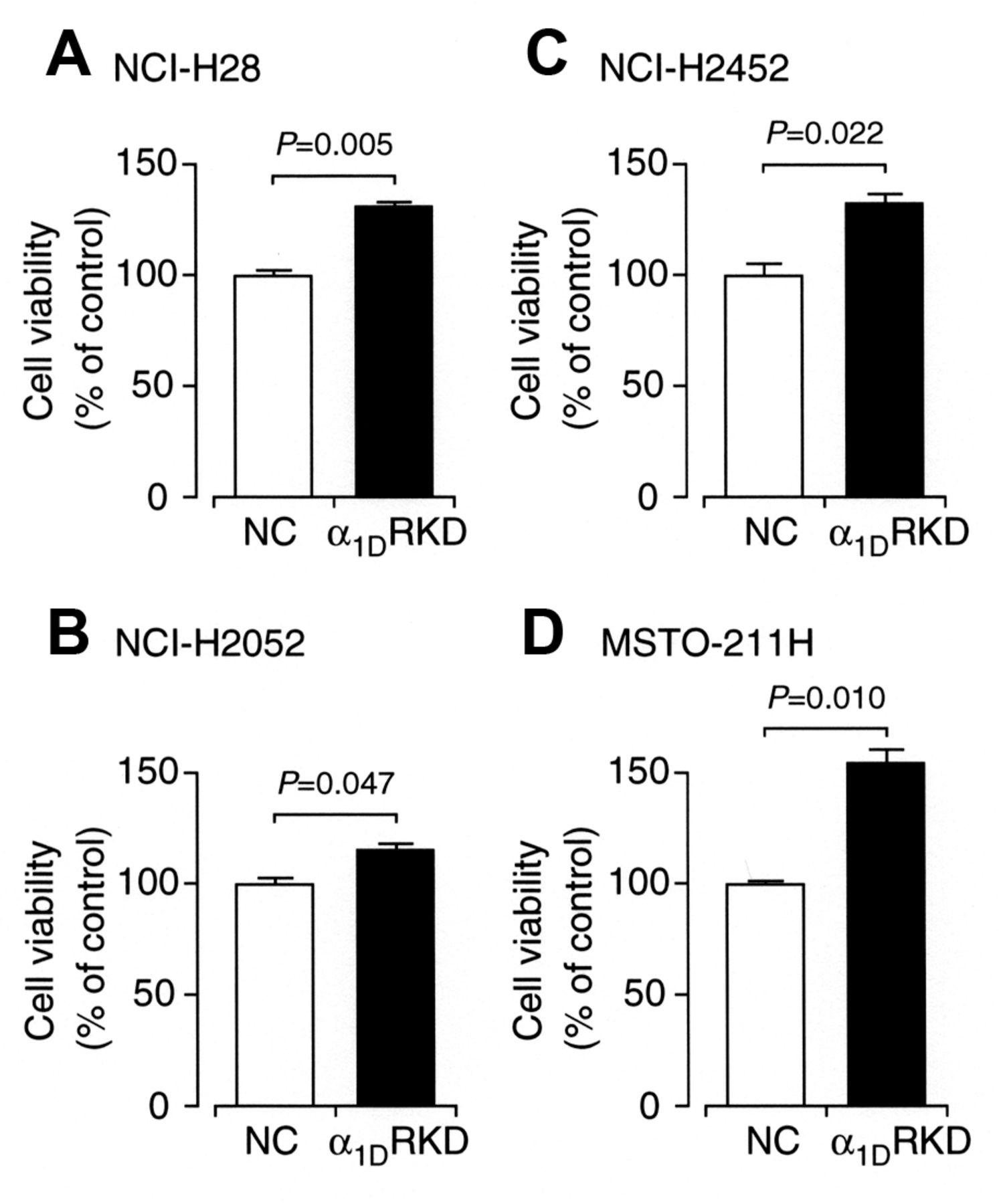
To obtain further evidence for naftopidil- or prazosin-induced apoptosis independent of α1-adrenoceptor blocking, an α1DR siRNA was constructed and transfected into malignant mesothelioma cells. Expression of α1D-adrenoceptor protein for cells transfected with α1DR siRNA significantly decreased as compared with the expression for cells transfected with the NC siRNA (Figure 4A-D), confirming α1D-adrenoceptor knock-down. Cell viability for all the malignant mesothelioma cell lines was not reduced by knocking-down α1D-adrenoceptor, but conversely, it was increased (Figure 5A-D). This indicates that malignant mesothelioma cell apoptosis is not induced by blocking α1D-adrenoceptor; in other words, naftopidil- and prazosin-induced apoptosis of malignant mesothelioma cells is not due to α1-adrenoceptor blocking action.
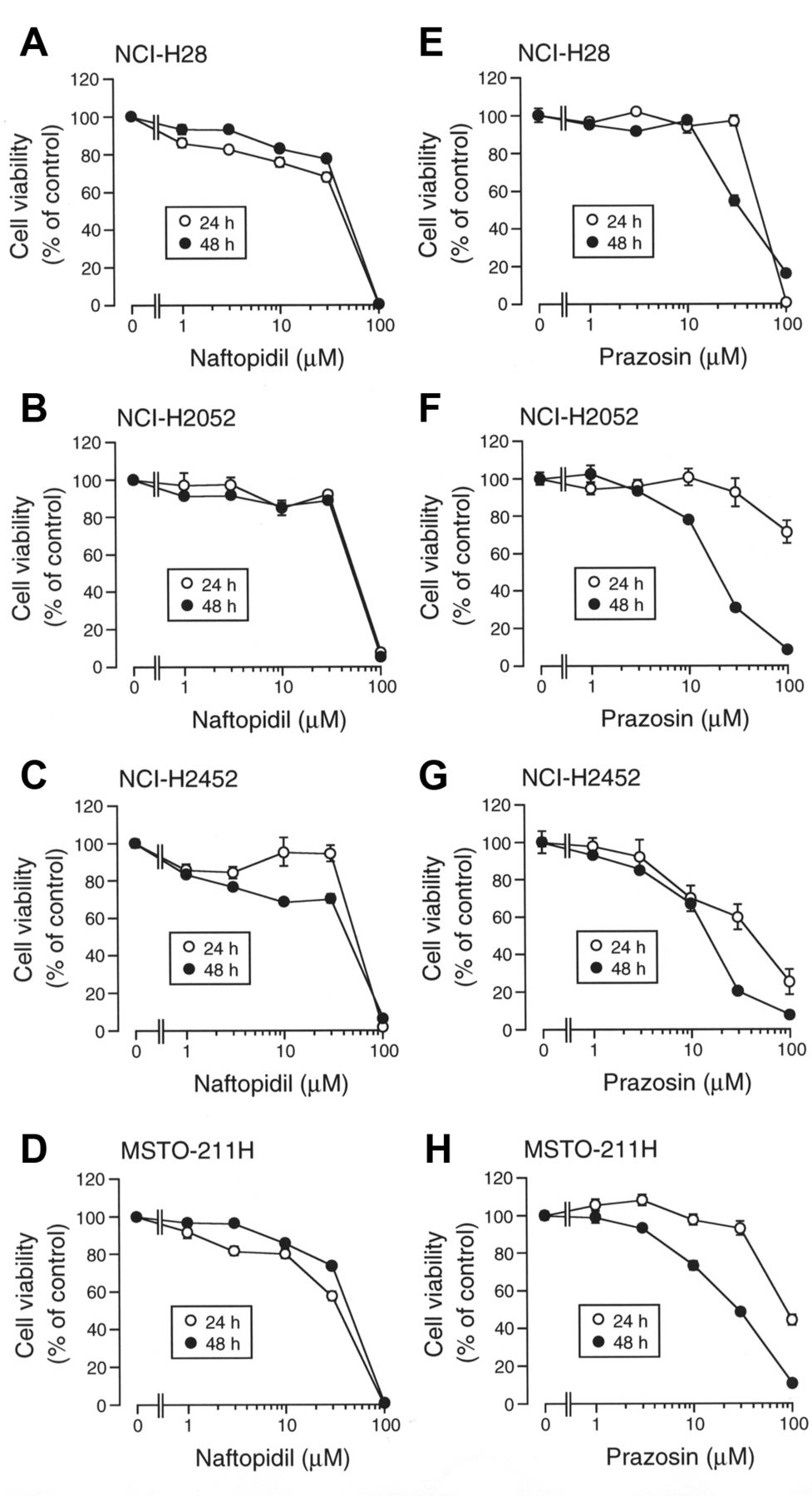
The effects of naftopidil and prazosin on cell viability. 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay was carried out on NCI-H28 (A, E), NCI-H2052 (B, F), NCI-H2452 (C, G), and MSTO-211H cells (D, H) untreated and treated with naftopidil or prazosin, at concentrations, as indicated for 24-48 h. In the graphs, each point represents the mean (−SEM) percentage of control cell viability (MTT intensities of cells untreated with naftopidil or prazosin) (n=4 independent experiments).
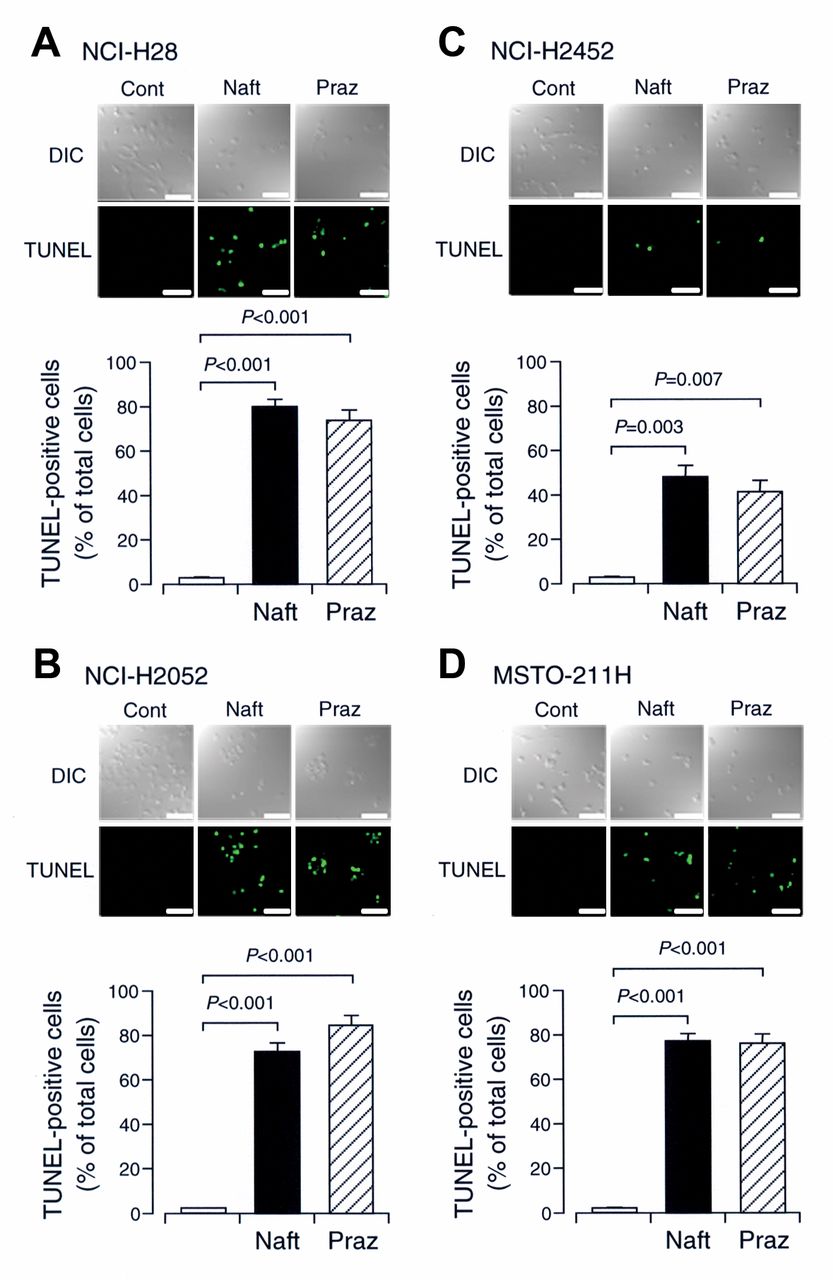
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining. TUNEL staining was carried out on NCI-H28 (A), NCI-H2052 (B), NCI-H2452 (C), and MSTO-211H cells (D) untreated (Cont) and treated with naftopidil (Naft) (100 μM) or prazosin (Praz) (100 μM) for 12 and 24 h, respectively. DIC, Differential interference contrast. Bars=100 μm. TUNEL-positive cells were counted in an area (0.4 mm × 0.4 mm) selected randomly. In the graphs, each column represents the mean (−SEM) percentage of TUNEL-positive cells relative to total cells (n=4 independent experiments). p-Values were defined from unpaired t-test.
Naftopidil activates caspase-3/-8 in malignant mesothelioma cells. For all the malignant mesothelioma cell lines examined here, naftopidil significantly activated caspase-3 and -8, but otherwise did not activate caspase-9 except for NCI-H28 cells (Figure 6A-D). Prazosin also activated caspase-3 and - 8 without affecting caspase-9 activity except for MSTO-211H cells (Figure 6E-H). These results imply that naftopidil and prazosin activate caspase-8 and the effector caspase-3, thereby inducing apoptosis of malignant mesothelioma cells. The results also suggest that naftopidil and prazosin could still activate caspase-9 followed by caspase-3 for certain types of malignant mesothelioma cell.

The effect of protein kinase-C inhibitor GF109203X on cell death induced by naftopidil or prazosin. NCI-H28 (A, E), NCI-H2052 (B, F), NCI-H2452 (C, G), and MSTO-211H cells (D, H) were treated with naftopidil (50 μM) or prazosin (50 μM) for 24 h in the presence and absence of GF109203X (GF) (100 nM), and then 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay was carried out. In the graphs, each column represents the mean (−SEM) percentage of control cell viability (MTT intensities of cells untreated with naftopidil or prazosin in the absence of the inhibitor) (n=4 independent experiments). p-Values were defined from unpaired t-test. NS, Not significant.
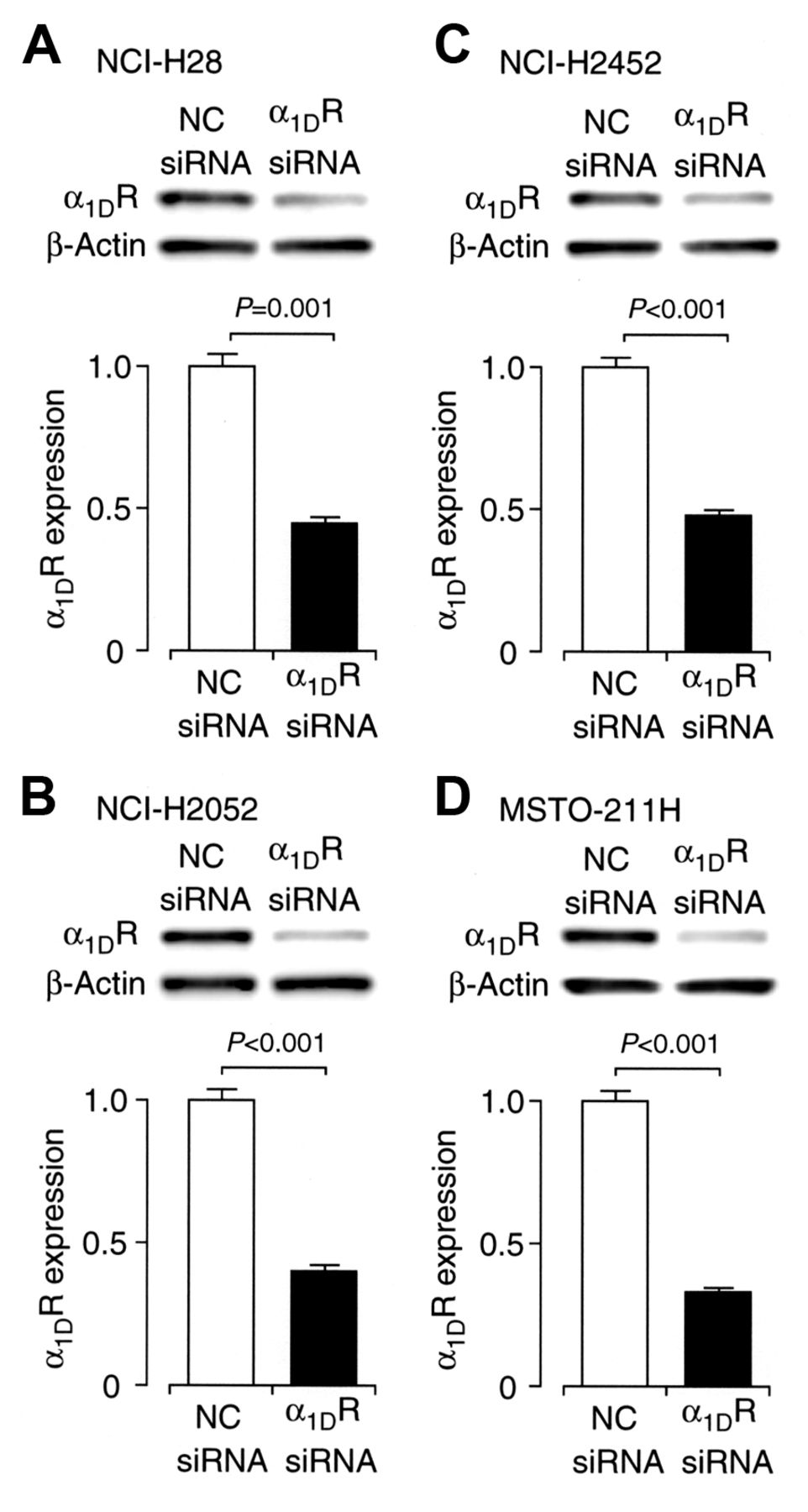
α1D Adrenoceptor knock-down. Western blotting for NCI-H28 (A), NCI-H2052 (B), NCI-H2452 (C), and MSTO-211H cells (D) transfected with negative control (NC) siRNA or α1DR siRNA 48 h after transfection. Signal intensities for α1D-adrenoceptor protein were normalized by those for β-actin. In the graphs, each column represents the mean (−SEM) α1D-adrenoceptor protein intensity (n=4 independent experiments). p-Values were defined from unpaired t-test.
Discussion
Naftopidil, an inhibitor of α1A- and α1D-adrenoceptors, has been clinically used for the treatment of benign prostate hyperplasia and hypertension (26). Interestingly, recent evidence has shown that naftopidil exerts an antitumor action on prostate cancer cells (27, 28). In the present study, naftopidil induced apoptosis in the human malignant mesothelioma cell lines NCI-H28 and NCI-H2052 sarcomatoid cells, NCI-H2452 epithelioid cells, and MSTO-211H biphasic cells. A similar effect was obtained with prazosin, another α1-adrenoceptor blocker. α1-Adrenoceptor blockers such as naftopidil and prazosin, thus, might exert an antitumor action on malignant mesothelioma cells.

The effect of α1D-adrenoceptor knock-down on cell viability. 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay for NCI-H28 (A), NCI-H2052 (B), NCI-H2452 (C), and MSTO-211H cells (D) transfected with negative control siRNA (NC) or α1DR siRNA (α1DR KD) 48 h after transfection (n=4 independent experiments). In the graphs, each column represents the mean (−SEM) percentage of control cell viability (MTT intensities for cells transfected with NC siRNA at 48 h after transfection)(n=4 independent experiments). p-Values were defined from unpaired t-test.
α1-Adrenoceptor linked to Gq/11 protein engages PKC activation. Naftopidil and prazosin, therefore, should inhibit PKC following α1-adrenoceptor blocking. Surprisingly, naftopidil- and prazosin-induced cell death of all the malignant mesothelioma cell lines and some cell lines, respectively, was attenuated by the PKC inhibitor GF109203X. In addition, malignant mesothelioma cell death was not induced by knocking-down the α1D-adrenoceptor; conversely, cell proliferation was promoted. Taken together, these results show that naftopidil and prazosin are likely to induce apoptosis of malignant mesothelioma cells by a mechanism independent of α1-adrenoceptor blocking. In support of this, α1-adrenoceptor antagonists have been shown to modulate differentiation and death of human erythroleukemia cells, regardless of α1-adrenoceptor blocking (32).
Naftopidil and prazosin activated caspase-3 and -8 in all the investigated malignant mesothelioma cell lines. This indicates that they activate caspase-8 and the effector caspase-3 to induce apoptosis of malignant mesothelioma cells. Naftopidil and prazosin also activated caspase-9 for some malignant mesothelioma cell lines. This suggests that naftopidil and prazosin could still ultimately activate caspase-3. Caspase-8 is recognized as being activated through death receptors such as tumor necrosis factor receptor-1 (TNFR1), FAS/apoptosis antigen-1 (APO1)/CD95, death receptor-3 (DR3)/APO3/WSL-1/lymphocyte-associated receptor of death (LARD)/TRAMP, DR4/TNF-related apoptosis-inducing ligand receptor-1 (TRAIL-R1), DR5/TRAIL-R2/TNF-related apoptosis-inducing ligand receptor inducer of cell killing-2 (TRICK2)/KILLER, and DR6 (33). FAS, activated by FASL, recruits the adaptor protein FAS-associated protein with death domain (FADD) to aggregate procaspase-8, which cleaves to initiate the active form of caspase-8 (34). TNFR1, activated by TNF-α, alternatively, forms a complex of TNFR1-associated death domain (TRADD)/receptor interacting protein-1 (RIP1)/FADD/procaspase-8 to activate caspase-8 (35). In contrast, caspase-9 is activated in concert with mitochondrial damage, allowing cytochrome c efflux from the mitochondria into the cytosol, to form an apoptosome complex with apoptotic protease activating factor 1 (APAF-1) or dATP (36-38). How naftopidil or prazosin activates caspase-8 or caspase-9 in malignant mesothelioma cells remains to be explored. To address this question, we are currently carrying out further experiments.
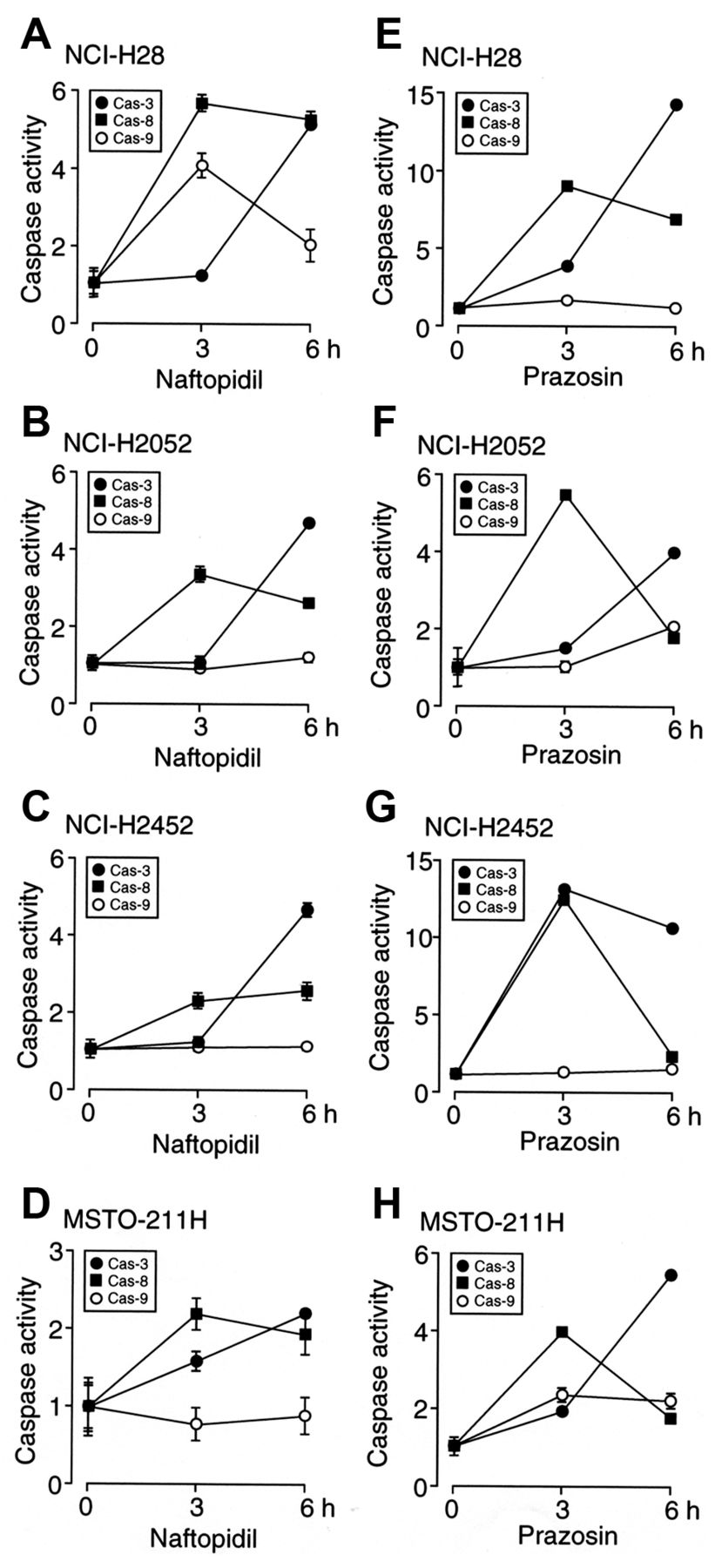
Activities of caspase-3, -8, and -9. NCI-H28 (A, E), NCI-H2052 (B, F), NCI-H2452 (C, G), and MSTO-211H cells (D, H) were treated with naftopidil (100 μM) or prazosin (100 μM) for 3-6 h, and then activities of caspase-3, -8, and -9 were enzymatically assayed. In the graphs, each point represents the mean (−SEM) ratio to basal caspase activities (before treatment with naftopidil or prazosin) (n=4 independent experiments).
Conclusion
The results of the present study show that naftopidil and prazosin have the potential to induce apoptosis of malignant mesothelioma cells by activating caspase-8 and the effector caspase-3, regardless of α1-adrenoceptor blocking. Naftopidil, which has been permitted for clinical use, may have a future role in treatment of human malignant mesothelioma.
Footnotes
-
↵* These Authors contributed equally to this work.
-
Conflicts of Interest
None of the Authors have any potential conflicts of interest.
- Received December 13, 2012.
- Revision received January 28, 2013.
- Accepted January 30, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}