


Abstract
Background: Hepatocellular carcinoma (HCC) is one of the most common types of malignant tumors worldwide, for which the prevalence and mortality rates are very high in Taiwan. Caveolin-1 (CAV-1) is a main structural protein of caveolae and plays a regulatory role in signaling pathways and tumorigenesis. High expression of Cav-1 in mouse HCC is positively correlated with higher cell invasive capacity, but the contribution of CAV-1 genetic variants during HCC progression is still largely unknown. In this study, we investigated the contribution of CAV-1 variant to the risk of HCC from the analyses of DNA, RNA and proteins. Materials and Methods: We enrolled 298 patients with HCC and 298 cancer-free controls, frequency-matched by age and gender in this case–control study. Firstly, the associations of six single nucleotide polymorphisms (SNPs) of the Cav-1 gene at C521A (rs1997623), G14713A (rs3807987), G21985A (12672038), T28608A (rs3757733), T29107A (rs7804372), and G32124A (rs3807992) with HCC risk in a Taiwanese population were evaluated. Secondly, thirty HCC tissue samples with variant genotypes were tested to estimate for CAV-1 mRNA expression by real-time quantitative reverse transcription. Finally, the HCC tissue samples of variant genotypes were examined by western blotting to estimate their CAV-1 protein expression patterns. Results: There were significant differences between the HCC and control groups in the distributions of the CAV-1 G14713A genotypes (p=0.0124), and these carrying AG and AA genotypes had a higher risk for HCC, compared with those with the GG genotype (odds ratio=1.51 and 1.94, respectively). Patients with CAV-1 G14713A AG or AA genotype had higher levels of mRNA (p=0.0001) and protein (p=0.0019) than those with the GG genotype. Conclusion: Our multi-approach findings at the DNA, RNA and protein levels suggest that CAV-1 may play a critical role in HCC carcinogenesis, and serve as a target for HCC therapy.
- Caveolin-1
- hepatocellular carcinoma
- polymorphism
- genotype
- real-time quantitative reverse transcription
- western blot
Hepatocellular carcinoma (HCC) is the leading cause of malignant cancer death in the world, with most cases occuring in Africa, Western countries, China and Taiwan (1, 2). Limited treatment and poor prognosis of this disease emphasize the importance in developing an effective chemoprevention. However, the exact molecular mechanism of HCC development is still unclear (1).
Three caveolin (CAV) proteins, CAV-1, -2 and -3, serve as the structural components of the caveolae and also function as scaffolding proteins, which are capable of recruiting numerous cascade signaling molecules to the caveolae and regulating their activity. It has been reported for a caveolin-deficient animal model that caveolins play a role in human disease processes, including diabetes, cancer, cardiovascular diseases, atherosclerosis, pulmonary fibrosis and a variety of degenerative muscular dystrophies (3). CAV-1, a protein of 178 amino acids, was initially identified as a tumor suppressor gene (4). It has been demonstrated that CAV-1 is down-regulated in sarcoma, lung carcinoma and ovarian carcinoma (5-7). However, elevated expression of CAV-1 has also been reported to be associated with the metastasis of esophageal squamous cell carcinoma and prostate cancer, and to be negatively correlated with patient survival (8, 9). These findings indicate that the role of CAV-1 may be multi-faceted, depending on the involved tissue. In literature, some epidemiological studies have investigated the association between the CAV-1 single-nucleotide polymorphism (SNP) and the risk for various types of cancer, including nasopharyngeal carcinoma (10), non-small cell lung carcinoma (11), prostate (12-14), breast (15, 16), oral (17), colorectal (18) and bladder cancer (19).
As for the role of CAV-1 in HCC, most evidence has come from the studies in mouse models. It has been reported that CAV-1 was highly expressed in mouse hepatoma cells with lymphatic metastasis potential and increased their invasive ability by up-regulating glycosylation of CD147 (20). In 2008, the same group found that exogenous expression of CAV-1 in Hepa1-6 cells enhanced cell transformation capability both in vitro and in vivo and prevented actinomycin D-induced apoptosis via the activation of survivin-mediated survival pathway (21). Conversely, down-regulation of CAV-1 in Hca-F cells significantly attenuated cell transformation ability in vitro and in vivo and increased cell sensitivity to actinomycin-D by inhibiting survivin-mediated survival pathway (21). In 2009, the same group reported that highly expressed CAV-1 in mouse hepatoma H22 cells could be suppressed by siRNA of Cav-1, which resulted in a reduced cell migration capacity in vivo and in vitro (22). In addition, down-regulation of CAV-1 can also promote apoptosis of H22 cells in vivo and in vitro (22).
To our knowledge, the association of Cav-1 SNPs with HCC has never been reported. Thus, the objectives of the current study were i) to determine the genotypic frequency of six polymorphisms of the CAV-1 gene at C521A (rs1997623), G14713A (rs3807987), G21985A (12672038), T28608A (rs3757733), T29107A (rs7804372), and G32124A (rs3807992) in a Taiwanese population with HCC; and ii) to investigate the functional phenotype of the CAV-1 G14713A genotype. We assumed that variant CAV-1 genotypes and phenotypes may contribute to HCC susceptibility. To test this hypothesis, our present study was designed to investigate the association of CAV-1 genotypes with risk of HCC in our hospital-based case–control study in a central Taiwanese population. In addition, we investigated the association of CAV-1 mRNA and protein expression patterns with HCC risk, by real-time polymerase chain reaction (PCR) and western blot respectively, to assess the potential functional effect of CAV-1 genotype on HCC risk. To the best of our knowledge, this is the first study to evaluate the relationship of CAV-1 genotype/phenotype and HCC susceptibility using DNA, RNA and protein analyses.
Materials and Methods
Study population. Two-hundred and ninety-eight patients diagnosed with HCC were recruited at the Departments of General Surgery at the China Medical University Hospital, Taiwan, in 2004-2010. Each patient and non-cancerous healthy person, matched by gender, age and individual habits, such as smoking and alcohol drinking, from a random sampling from the Health Examination Cohort of China Medical University Hospital, completed a self-administered questionnaire and provided their peripheral blood samples. Each patient donated 3-5 ml of venous blood and their tumor and non-tumor tissues after providing written informed consent. The study was approved by the Institutional Review Board of China Medical University Hospital.
PCR-restriction fragment length polymorphism genotyping conditions. Genomic DNA was prepared from peripheral blood leucocytes using a QIAamp Blood Mini Kit (Blossom, Taipei, Taiwan, ROC) (23-25), and the genotyping was performed according to published methods (17, 19). Briefly, the following primers were used for CAV-1 C521A (rs1997623): 5’-GTGTCCGCTTCTGCTATCTG-3’ and 5’-GCCAAGATGCAGAAGGAGTT-3’; for CAV-1 G14713A (rs3807987): 5’-CCTTCCAGTAAGCAAGCTGT-3’ and 5’-CCTCTCAATCTTGCCATAGT-3’; for CAV-1 G21985A (12672038): 5’-GGTGTCAGCAAGGCTATGCT-3’ and 5’-CCAGACACTCAGAATGTGAC-3’; for CAV-1 T28608A (rs3757733): 5’-GCTCAACCTCATCTGAGGCA-3’ and 5’-GGCCTATTGTTGAGTGGATG-3’; for CAV-1 T29107A (rs7804372): 5’-GCCTGAATTGCAATCCTGTG-3’ and 5’-ACGGTGTGAACACGGACATT-3’ and for CAV-1 G32124A (rs3807992): 5’-GGTGTCTTGCAGTTGAATG-3’ and 5’-ACGGAGCTACTCAGTGCCAA-3’. The following cycling conditions were performed: one cycle at 94°C for 5 min; 35 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 30 s; and a final extension at 72°C for 10 min. The PCR products were studied after digestion with AvrII, BfaI, HaeIII, Tsp509I, Sau3AI and NlaIII, restriction enzymes for CAV-1 C521A (cut from 485 bp C type into 170+315 bp A type), CAV-1 G14713A (cut from 268 bp A type into 66+202 bp G type), CAV-1 G21985A (cut from 251+43 bp A type into 153+98+43 bp G type), CAV-1 T28608A (cut from 298 bp T type into 100+198 bp A type), CAV-1 T29107A (cut from 336 bp A type into 172+164 bp T type) and CAV-1 G32124A (cut from 213+142+67 bp A type into 142+118+95+67 bp G type), respectively.
Semiquantitative RT-PCR analysis for CAV-1 expression pattern. To evaluate the correlation between the CAV-1 mRNA expression and CAV-1genotype, 30 surgically-removed liver tissue samples adjacent to tumors with different genotypes were subjected to extraction of the total RNA using Trizol Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's protocol. The expression pattern of CAV-1 RNA was measured by real-time quantitative RT-PCR using FTC-3000 real-time quantitative PCR instrument series (Funglyn Biotech Inc., Toronto, ON, Canada). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal quantitative control. The primers used for amplification of CAV-1 mRNA were: forward 5’-CTCGAGATGTCTGGGGGCAAATACG-3’ and reverse 5’-GAATTCTATCTCTTTCT-GCGTGCTG-3’, while for GAPDH the primers were: forward 5’-GAAATCCCATCACCATCTTCCAGG-3’ and reverse 5’-GAGCCCCAGCCTTCTCCATG-3’. Fold changes were normalized by the levels of GAPDH expression, and each assay was carried out at least in triplicate.
Western blot analysis. The 30 pairs of liver specimens were homogenized in RIPA lysis buffer (Upstate Inc., Lake Placid, NY, USA), the homogenates were centrifuged at 10,000 ×g for 30 min at 4°C, and the supernatants were used for western blotting. Samples were denatured by heating at 95°C for 10 min, then separated on a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel, and transferred to a nitrocellulose membrane. The membrane was blocked with 5% non-fat milk and incubated overnight at 4°C with a rabbit anti-mouse CAV-1 antibody (1:500; Santa Cruz Biotech, Santa Cruz, CA, USA) in 5% powdered skimmed milk buffer. The membrane was then washed thrice with PBS with 0.1% Tween 20, and incubated with the corresponding secondary antibody of anti-rabbit-horseradish peroxidase (1:2000; Santa Cruz Biotech) for 1 h at room temperature. After reaction with electrogenerated chemiluminescence (ECL) solution (Amersham, Arlington Heights, IL, USA), all the bands were visualized using a chemiluminescence imaging system (Syngene, Cambridge, UK). The optical density of each specific band was measured using a computer-assisted imaging analysis system (Gene Tools Match software; Syngene).
Distributions of selected characteristics between hepatocellular carcinoma cases and controls.
Statistical analyses. To ensure that the controls used were representative of the general population and to exclude the possibility of genotyping error, the deviation of the genotype frequencies of CAV-1 SNPs in the controls from those expected under the Hardy-Weinberg equilibrium was assessed using the goodness-of-fit test. Pearson's Chi-square test or Fisher's exact test (when the expected number in any cell was less than five) was used to compare the distribution of the CAV-1 genotypes between cases and controls. The associations between the CAV-1 polymorphisms and HCC risk were estimated by computing odds ratios (ORs) and their 95% confidence intervals (CIs) from unconditional logistic regression analysis with the adjustment for possible confounders. A value of p<0.05 was considered statistically significant, and all statistical tests were two-sided.
Results
Basic comparisons between the case and control groups. The characteristics of the controls and cases are summarized in Table I. The characteristics of patients and controls are all well-matched. None of the frequency distributions of the two groups were statistically different from each other (p>0.05).
Association of CAV-1 genotypes and HCC susceptibility. The frequencies of the genotypes for CAV-1 C521A, G14713A, G21985A, T28608A, T29107A and G32124A between the controls and patients with HCC are shown in Table II. For the CAV-1 G14713A (rs3807987) genotyping, the ORs for these carrying AG and AA genotypes were 1.51 (95% CI=1.05-2.17) and 1.94 (95% CI=1.13-3.33) respectively, compared to those carrying the wild-type TT genotype. The p-value for trend was significant (p=0.0330). In the dominant model (AG plus AA versus GG), the association between CAV-1 G14713A genotype and the risk for HCC was found to be statistically significant (OR=1.61, 95% CI=1.16-2.25). The small percentage of these with AA genotype caused a border-line effect (OR=1.69, 95% CI=0.99-2.87, p=0.0652) in the recessive model (AA versus GG plus AG). Regarding the results of CAV-1 C521A, G21985A, T28608A, T29107A and G32124A polymorphisms, the distributions of these polymorphisms were in Hardy-Weinberg equilibrium and there was no difference between HCC and control groups in the distribution in the genotypic frequency at these SNPs (Table II). To sum up, the genotyping results indicated that individuals carrying a variant A allele at G14713A maybe at higher risk of HCC.
Correlation of CAV-1 G14713A genotype and the expression level of Cav-1 mRNA and protein. We collected 30 surgically-removed liver tissue samples adjacent to tumors for phenotypical study. These samples were obtained from the patients before any therapy or analysis. The frequencies of the GG, AG, and AA genotypes for CAV-1 G14713A were 22, 5, and 3, respectively. The effects of these three genotypes on the transcriptional expressions of mRNA were measured and evaluated by real-time quantitative RT-PCR (Figure 1). The average level of mRNA for AG, and AA genotypes for CAV-1 G14713A were 1.45-, and 1.43-fold, compared with the GG genotype, respectively. The three samples with the AA genotype were added to the samples of the AG genotype for effective statistical analysis, and a statistically significantly higher level of Cav-1 mRNA expression was identified in samples from patients with the AG/AA genotype than from those with the GG genotype (p=0.0001) (Figure 1).
Distributions and analysis of caveolin-1 (CAV-1) genotypic frequencies among hepatocellular carcinoma cases and controls.
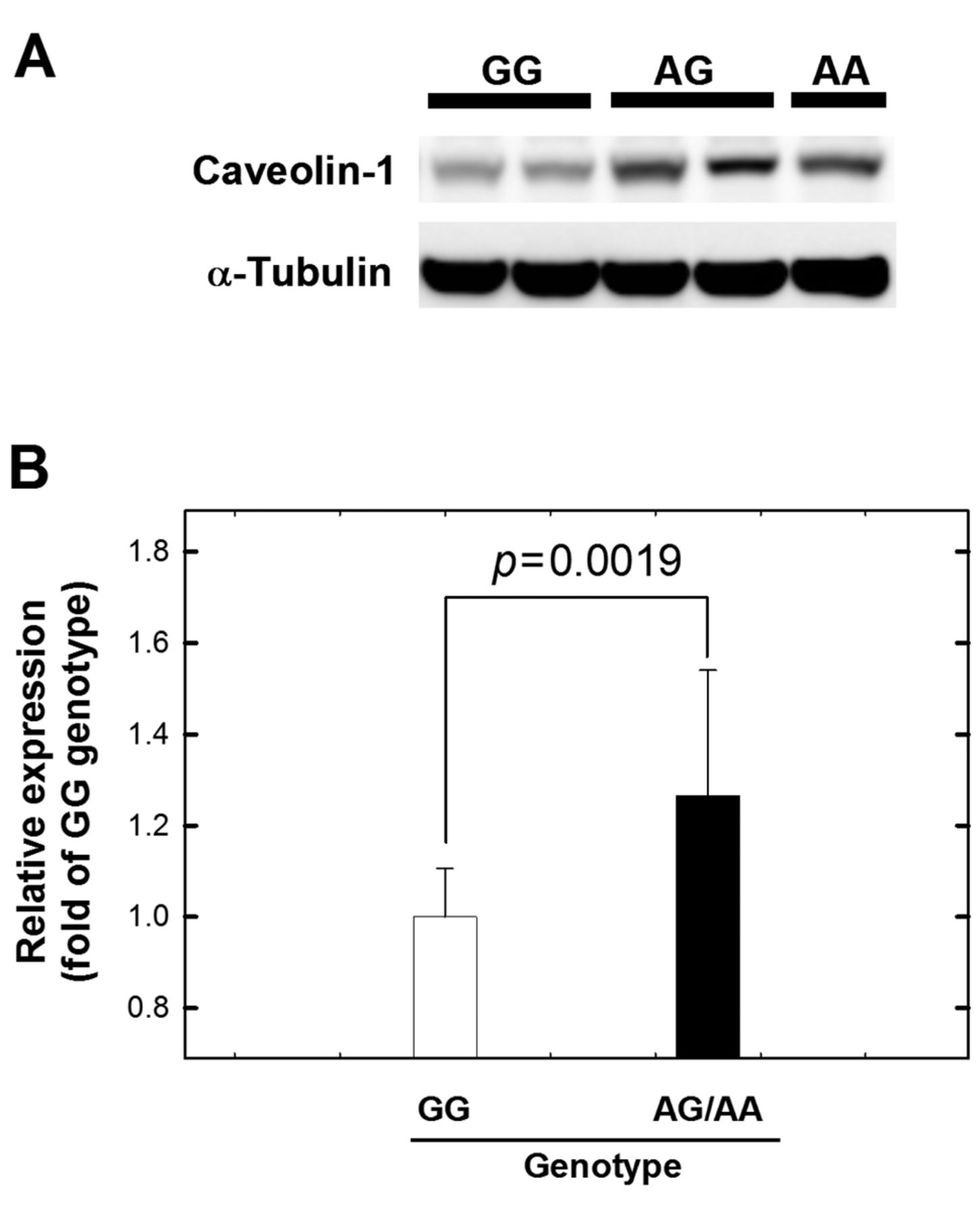
We also examined the CAV-1 protein expression patterns in HCC tumors from patients with GG, AG, and AA genotypes for CAV-1 G14713A (Figure 2). We performed western blot and the results showed that CAV-1 was expressed at a lower level in the tissues of these with GG genotype than those with AG or AA genotypes (p=0.0019) (Figure 2). To sum up, the results at the RNA and protein levels showed that the CAV-1 G14713A genotype may play an important role in HCC etiology.
Discussion
In this study, the association of CAV-1 polymorphism and HCC risk was investigated in Taiwan, where the prevalence of hepatitis B and C viruses is the highest worlwide. We have genotyped six SNPs, C521A (rs1997623), G14713A (rs3807987), G21985A (12672038), T28608A (rs3757733), T29107A (rs7804372), and G32124A (rs3807992) in HCC cases and non-cancer controls. After the genotyping, we found that individuals carrying the AG and AA genotypes were at higher risk of HCC compared with those carrying the GG genotype for CAV-1 G14713A. As for the other SNPs, there was no differential genotypic distribution between the case and control groups (Table II).

Analysis of Caveolin-1 (CAV-1) mRNA expression levels. A: Quantitative real-time polymerase chain reaction (RT-PCR) for CAV-1 from liver tissue samples of three genotypes was performed and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal quantitative control. Fold changes were normalized by the level of GAPDH expression, and each assay was performed at least in triplicate. B: The groups of AG and AA in (A) were pooled and compared with GG group.
The CAV-1 G14713A polymorphic site mapped to the intron region of CAV-1, which does not directly result in any amino acid coding alteration. After finding that the genotype of CAV-1 G14713A is associated with HCC risk, we were interested in designing a functional investigation to determine whether the Cav-1 G14713A SNP influenced the mRNA and protein level of CAV-1. We collected HCC tumor from patients and selected 22, 5, and 3 of them, which were of the GG, AG and AA genotypes, and performed RT-PCR and western blot. In the results of real-time quantitative RT-PCR, we found that the tissues from these with CAV-1 G14713A AG and AA genotypes indeed had a higher expression of CAV-1 mRNA than those with GG genotype (Figure 1). Following the central dogma of molecular biology, the protein level results also showed that the tissues from these with CAV-1 G14713A AG and AA genotypes indeed had also a higher expression of the CAV-1 protein than those with GG genotype (Figure 2). The A allele of CAV-1 G14713A might somehow code for a higher level of Cav-1 mRNA, which led to an increased expression of CAV-1 protein and elevated HCC risk. To the best of our knowledge, this is the first study of the role of CAV-1 in HCC with findings from multi-faceted DNA, RNA and protein assays.

The expression of caveolin-1 (CAV-1) in HCC tissues from patients of different CAV-1 genotypes. Representative photographs show that different expression levels were observed in cancerous tissues from cases with different CAV-1 genotypes. A: Western blot analysis of CAV-1 expression in tumor tissues from cases with GG, AG and AA CAV-1 genotypes. B: Quantification of the western blot data from Figure 2A. α-Tubulin was used as the loading control. Data are averaged from at least six replicates from the tissues of each group with 15 μg total sample protein for each lane.
In the literature, the role of CAV-1 in tumor biology remains controversial. Several studies supported the idea that it may act as a tumor suppressor. Firstly, down-regulation of CAV-1 has been found in many types of cancer cells, and suppression of CAV-1 expression was sufficient to induce transformation of NIH 3T3 cells (6, 20, 26-29). Secondly, up-regulation via exogenous expression of CAV-1 in human breast cancer cells suppressed cellular transformation and survival capability (30, 31). On the contrary, there is also mounting evidence suggesting that CAV-1 also acts as a tumor promoter in certain types of cancer cells. Firstly, CAV-1 was highly expressed in prostate cancer, and the expression of CAV-1 is positively correlated with tumor grade and stage (32, 33). Secondly, similar findings have been reported for bladder and esophageal cancer (8, 34). Thus, the CAV-1 gene seems to act as both a tumor suppressor gene and an oncogene.
Functional investigations of CAV-1 in HCC are scarce and most came from studies in mouse models (21, 22, 35, 36). Accumulating epidemiological studies have reported that the CAV-1 genotype is associated with human diseases, including kidney transplant fibrosis and allograft failure (37), nasopharyngeal carcinoma (10), non-small cell lung carcinoma (11), prostate (12-14), breast (15, 16), oral (17), colorectal (18) and bladder cancer (19). Here, we reported that the genotypes of CAV-1 were also associated with HCC. Consistent with previous findings, the A allele of CAV-1 G14713A was associated with a higher cancer susceptibility (10, 14, 16-19). We also further examined the expression levels of CAV-1 mRNA and protein in HCC tissues, finding the A allele to be positively correlated with a higher level of mRNA and protein expression (Figures 1 and 2).
In conclusion, our study firstly found that the Cav-1 G14713A AG and AA variant genotypes were associated with a higher HCC susceptibility of Taiwanese, and that the A allele may serve as a predictor not only for higher CAV-1 mRNA and protein expression, but also of higher HCC risk.
Acknowledgements
This study was supported by research grants from the Terry Fox Cancer Research Foundation and China Medical University and Hospital (DMR-102-066). The assistance from Ping-Fang Wang in data collection, and from Liang-Yi Lin, Yi-Ting Chang, Hong-Xue Ji in genotyping was highly appreciated by the Authors.
Footnotes
-
↵*These Authors contributed equally to this work.
- Received December 8, 2012.
- Revision received January 12, 2013.
- Accepted January 14, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- The Contribution of Matrix Metalloproteinase-7 Promoter Genotypes to Hepatocellular Carcinoma Susceptibility
- The Contribution of Matrix Metalloproteinase-1 Genotypes to Hepatocellular Carcinoma Susceptibility in Taiwan
- Tumor Necrosis Factor-{alpha} Genotypes Are Associated with Hepatocellular Carcinoma Risk in Taiwanese Males, Smokers and Alcohol Drinkers
- Evaluation of the Contribution of Methylenetetrahydrofolate Reductase Genotypes to Taiwan Breast Cancer
- Association of Caveolin-1 Genotypes with Gastric Cancer in Taiwan
- A Novel Caveolin-1 Biomarker for Clinical Outcome of Sarcopenia
- Significant Association of Caveolin-1 (CAV1) Genotypes with Upper Urothelial Tract Cancer