


Abstract
Background: The Human Epidermal Growth Factor Receptor 2 (HER-2), overexpressed in 25-30% of breast carcinomas (BC), is the therapeutic target for trastuzumab, a recombinant humanized monoclonal antibody. The initial response to trastuzumab is often followed by drug-insensitivity within one year. Several hypotheses have been raised to explain this event, but the mechanisms behind the responses to trastuzumab are still unclear. Aim: To study the effects of short and prolonged trastuzumab treatment on the proteomic profiles of HER-2-overexpressing SKBR-3 BC cells. Materials and methods: Cells were treated with trastuzumab to obtain sensitive and resistant clones. The drug effects were evaluated at the phenotypical and proteomic levels. Results: In the trastuzumab-resistant cells the expression of a large amount of proteins, initially affected by treatment, reverted to levels of the untreated cells. Conclusion: The results obtained so far illustrate for the first time a large-scale differential protein expression between trastuzumab-treated and untreated cells, and between trastuzumab-sensitive and resistant cells. We believe that the results obtained will help to increase the knowledge of the molecular effects of trastuzumab and will be useful to better-understand the drug resistance mechanisms.
The Human Epidermal Growth Factor Receptor 2 (HER-2) belongs to EGF tyrosine kinase receptor family that also includes HER-1, HER-3 and HER-4. Once activated by interaction with their cognate ligands, these receptors form homo- or heterodimers and trigger tyrosine kinase activity, which in turn, initiates a signal transduction cascade, leading to alternative routes towards cellular proliferation, differentiation, and survival (1). HER-2 is considered an ‘orphan’ receptor because it does not seem to interact with any known ligand in the form of a homodimer, while it is able to form heterodimers with other members of the HER family, thus activating multiple transduction pathways.
The overexpression of HER-2, primarily due to gene amplification, occurs in approximately 25-30% of invasive breast carcinomas (2). Several in vivo and in vitro studies have demonstrated that HER-2 overexpression is associated with increased cell proliferation and survival, resistance to apoptosis, activation of cell motility, promotion of angiogenesis and increased tendency for tumor metastasis (3, 4). Hence, HER-2 is considered a reliable prognostic and predictive marker, and a privileged therapeutic target in primary tumors and metastatic sites (5, 6). In addition, since the levels of HER-2 expression in human cancer cells are much higher than those of normal adult tissues, the latter are potentially less sensitive to the toxicity of HER-2-targeting drugs. In 1998 the United States Food and Drug Administration approved trastuzumab for clinical use against metastatic breast cancer that overexpress HER-2 (7).
Trastuzumab (Herceptin®) is a recombinant humanized monoclonal antibody against HER-2 whose main epitope corresponds to a sequence in the extracellular domain of HER-2, adjacent to the cell membrane (8, 9).
According to experimental studies, trastuzumab may act both directly and indirectly in cancer therapy. The direct effects concern: the inhibition of HER-2 internalization and its turnover from the cell membrane (10), the down-regulation of cell proliferation (11, 12), the possible induction of DNA damages and the simultaneous inhibition of DNA repair processes (13), and the blockage of the metalloproteinase-mediated shedding of HER-2 ectodomain (14). Among the indirect effects, the promotion of the antibody-dependent cytotoxicity by interaction with the Fc receptor (CD16) expressed by Natural Killer (NK) cells, has been reported (15).
Unfortunately, the initial response to trastuzumab-based regimens is often followed by drug-insensitivity occurring within one year in the majority of patients with HER-2-overexpressing breast cancer (16). Although several hypotheses have been raised to explain this event, the biochemical and molecular mechanisms behind the responses to trastuzumab are still unclear (17). Major difficulties lie in explaining if trastuzumab-resistant cells are pre-existing subclones of the tumour cell population, or the result of clonal selection occurring during treatment.
The proteomic approach, based on two-dimensional gel electrophoresis, is a powerful instrument to evaluate qualitative and quantitative modifications of protein profiling induced by pharmacological treatments of appropriate cellular systems. The present investigation aimed at studying and comparing the effects exerted by short- and long-term trastuzumab treatment on the proteomic profile of cells overexpressing HER-2. As a model system, we used SKBR-3 cells because of their ERBB2 gene amplification and strong overexpression of HER-2.
Materials and Methods
Cell culture. SKBR-3 cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA), and grown in RPMI-1640 (Gibco, Paisley, UK) with 10% Fetal Bovine Serum (FBS), containing 100 U/ml penicillin and 100 μg/ml streptomycin, at 37°C in an atmosphere of 5% CO2. The cell medium was replaced every 48 h.
The expression level of HER-2 was assessed by western blot analysis, using the 8701-BC cell line which does not overexpress HER-2 as a negative control (Figure 1). The 8701-BC cells were derived from an infiltrating ductal carcinoma and have been well-characterized (18) for their phenotypic and molecular features, including their proteomic profile (19, 20).
Cell proliferation assay. The growth rates of SKBR-3 cells were determined by a colorimetric method (Promega, Madison, WI, USA) using a kit based on 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxy-methonyphenol)-2-(4-sulfophenyl)-2H-tetrazolium (MTS)-to-formazan degradation, according to the manufacturer's instructions. Cell proliferation was recorded at different time intervals according to the experimental design.
Pharmacological treatment. Trastuzumab (Herceptin®; Genentech, South San Francisco, CA, USA) for present research was furnished by the operative unit of Pharmacology (La Maddalena Hospital, Level III Oncological Department), as residual aliquots after the preparation of the drug for patient therapy. The dose responses to the drug were tested at three different concentrations of trastuzumab, namely 1 μg/ml, 5 μg/ml, 10 μg/ml, by the administration of the drug doses at every change of the culture medium for seven days. Untreated SKBR-3 cells (WT) were used as control.
To obtain the sensitive (TS) clone, the SKBR-3 cells were treated for seven days with 10 μg/ml of trastuzumab, which represents the saturation dose (SD) for SKBR-3 cells (21). Cells surviving this treatment were collected, cultured and submitted to long-term treatment with 10 μg/ml trastuzumab, until the growth rate of the treated cells became synchronous with that of parental WT cells (75 days).
Sample preparation for proteomics. WT-, TS- and TR-SKBR-3 cells used for proteomic experiments were serum-starved for 24 h, harvested with a cell scraper in phosphate buffered saline (PBS) and lysed in M-RIPA buffer (50 mM Tris pH 7.5, 0.1% Nonidet P-40, 0.1% deoxycholate, 150 mM NaCl, 4 mM EDTA) added with a mixture of protease inhibitors [0.01% aprotinin, 10 mM sodium pyrophosphate, 2 mM sodium orthovanadate, 1 mM phenylmethanesulfonylfluoride (PMSF)].
Cellular lysates were centrifuged at 12,000 ×g for 10 min to clear cell debris, and the supernatants were dialyzed against ultrapure distilled water, lyophilized and stored at −80°C until analysis. Protein concentration in the cellular extracts was determined using the Bradford method (22).
2D gel electrophoresis (2D-E). 2D-E was performed essentially as described (23). Aliquots of the cell lysates were solubilized in a buffer containing 4% CHAPS, 40 mM Tris, 65 mM 1,4-dithioerythritol (DTE) and a trace amount of bromophenol blue in 8 M urea. The first dimensional separation was performed at 10°C on commercial sigmoidal immobilized pH gradient strips (IPG), 18-cm long with pH range 3.0-10, (Bio-Rad, Milan, Italy). The strips were rehydrated in 8 M urea, 2% CHAPS, 10 mM DTE and 0.5% carrier ampholytes (Resolyte 3.5-10, Amersham Bioscience, Milan, Italy). Aliquots of 45 μg (analytical gels) or 1.5 mg (preparative gels) of total proteins were applied to each gel strip.
The isoelectric focusing was carried out by linearly increasing voltage from 200 to 3500 V during the first 3 h, after which focusing was continued at 8000 V for 8 h. After the run, the IPG strips were equilibrated with a solution containing 6 M urea, 30% glycerol, 2% Sodium dodecyl sulfate (SDS), 0.05 M Tris–HCl pH 6.8 and 2% DTE for 12 min. The –SH groups were then blocked by substituting the DTE with 2.5% iodoacetamide in the equilibrating buffer. The focused proteins were then separated on 9-16% linear gradient of polyacrylamide gels (SDS-PAGE) with a constant current of 40 mA/gel at 10°C. The gels were stained with ammoniacal silver nitrate, digitized and processed with Image-Master 2D Platinum system (Amerham Bioscience, Milan, Italy) (24).
Mass spectrometry. Mass spectrometric analysis was carried out after in-gel digestion of protein spots, using sequencing-grade trypsin (20 μg/vial), according to the method of Shevchenko et al. (25) with some modifications (26, 27). The tryptic peptide extracts were dried in a vacuum centrifuge and re-dissolved in 10 μl of 0.1% trifluoroacetic acid (TFA). The matrix, a-cyano-4-hydroxycinnamic acid (HCCA), was purchased from Sigma–Aldrich (St. Louis, Missouri, USA). A saturated solution of HCCA (1 μl) at 2 mg/200 μl in CH3CN/H2O (50/50 (v/v), containing 0.1% TFA was mixed with 1 μl of peptide solution on the Matrix-assisted laser desorption/ionization (MALDI) plate and left to dry.
Mass spectra were recorded on a Voyager DE-PRO (AbSciex, Foster City, CA, USA) mass spectrometer, in the 500–5000 Da mass range, using a minimum of 100 shots of laser per spectrum. Delayed extraction source and reflector equipment allowed sufficient resolution to consider MH+ of monoisotopic peptide masses. Internal calibration was carried out using trypsin autolysis fragments at m/z 842.5100, 1045.5642 and 2211.1046 Da.

Western blot assay of proteins extracted from Human Epidermal Growth Factor Receptor-2 (HER-2)-amplified SKBR-3 cells and non-HER-2-amplified 8701-BC cells. Proteins extracted from the two cell lines were separated by a gradient (4-20%T) SDS-PAGE and probed with a mouse monoclonal antibody directed against the full-length HER-2 protein. The 200 kDa reference standard is marked.
Peptide mass fingerprinting was compared to the theoretical masses from the Swiss-Prot or U.S. National Center for Biotechnology Information (NCBI, Rockville Pike, Bethesda, MD, USA) sequence databases using Mascot (http://www.matrixscience.com/). Typical search parameters were as follows: ±50 ppm of mass tolerance, carbamidomethylation of cysteine residues, one missed enzymatic cleavage for trypsin, a minimum of four peptide mass hits was required for a match and methionine residues could be considered in oxidized form.
Western blot. The 1-D SDS-PAGE was performed in 4-20% gradient polyacrylamide mini-gels by loading 40 μg of each protein sample prepared according to Laemmli (28). For immunodetection the 1-D gels were electrotransferred onto nitrocellulose membranes (HyBond ECL; Amersham Bioscience) and stained with Ponceau S (Sigma-Aldrich). The membranes were then probed with the following antibodies from Santa Cruz Biotechnology (Heidelberg, Germany): anti-HER-2 (mouse monoclonal, sc-80898), anti-HER-1 (mouse monoclonal, sc-57092), anti-HER-3 (mouse monoclonal, sc-71067), anti-HER-4 (mouse monoclonal, sc-8050), anti-RAC-alpha serine/threonine-protein kinase (AKT1) (mouse monoclonal, sc-5298), anti-Insulin growth factor-1 receptor (IGF-IR) (mouse monoclonal, sc-81464), anti-Transforming growth factor-beta receptor type II (TGFB-RII) (rabbit polyclonal, sc-1700), anti-p27(KIP1) (mouse monoclonal, sc-56338), anti-p21(WAF1) (rabbit polyclonal, sc-756).
Following incubation with the appropriate secondary peroxidase-linked antibody (goat anti-mouse, sc-2005; goat anti-rabbit, sc-2030, Santa Cruz Biotechnology), the reaction was revealed by the ECL detection system, using high performance films (Hyperfilm ECL; Amersham Bioscience). ECL films were digitized and processed with Image Master-2D Platinum system (Amerham Bioscience), in order to perform densitometric quantification of the immunoreactive bands.
Results
Effects of trastuzumab on cell proliferation and cell behaviour. The effects of trastuzumab on cell proliferation were tested by administration of incremental doses of the drug, namely 1 μg/ml, 5 μg/ml and 10 μg/ml.

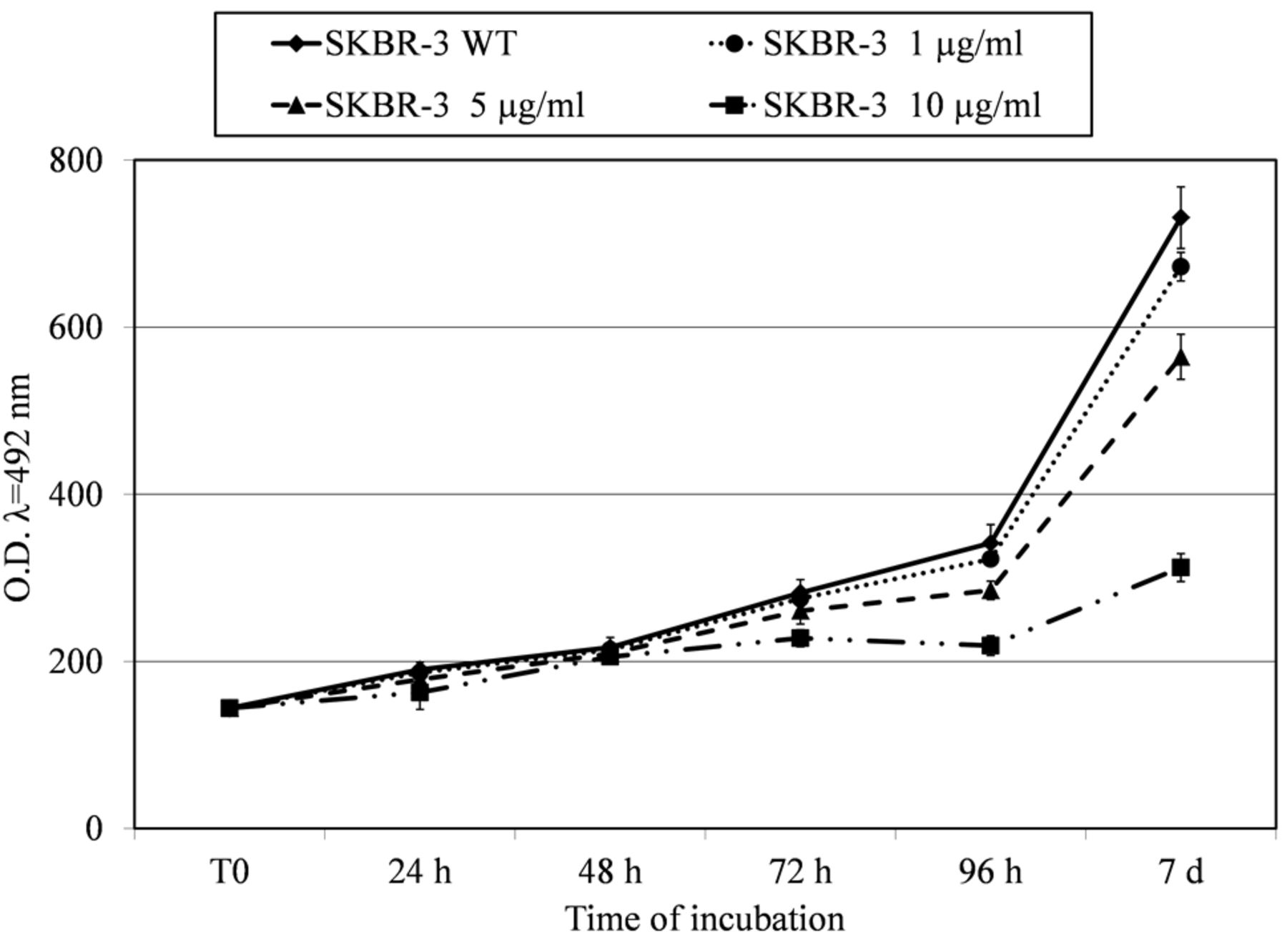
Growth curves of SKBR-3 cells treated with three different trastuzumab doses (1 μg/ml, 5 μg/ml and 10 μg/ml) for seven days and plotted with untreated cells used as reference. On the ordinate the cell concentration is reported as optical density at a wavelength of 492 nm (O.D. λ=492 nm).
As shown in Figure 2 the growth curves of the parental and responsive cells are very similar for the first 48 h. After this point, the rate appears significantly decreased in the cells treated with 10 μg/ml of trastuzumab, reaching about 60% of decrement at the seventh day from seeding, while minor effects were elicited at the other doses. Indeed the dose of 1 μg/ml caused a reduction of about 8% in the growth rate and the 5 μg dose of 23%. As reported by literature data (21) the amount of 10 μg/ml of trastuzumab represents a saturation dose (SD) for SKBR-3 cells. The cell clone sensitive to 10 μg/ml of trastuzumab was named trastuzumab-sensitive SKBR-3 (TS-SKBR-3).
The cells surviving treatment of trastuzumab for seven days, were collected, cultured and submitted to a long-term treatment with 10 μg/ml trastuzumab, until the growth rate of the treated cells became synchronous with that of the parental WT cells (75 days). This clone was named trastuzumab-resistant SKBR-3 (TR-SKBR-3).
The optical micrographs in Figure 3 show representative fields of the WT-SKBR-3, TS-SKBR-3 and TR-SKBR-3 cultures illustrating the effects of the drug at the SD on the cell morphology from 72 h to 7 days from seeding. As shown in Figure 3B, the cells treated with SD of the drug appear more dispersed and less adherent to the substrate on the seventh day in comparison with the untreated cells (Figure 3A); in addition, the treated cells display a more irregular morphology, with signs of cellular instability, such as the tendency to project several spikes and protrusions. At the same time of culture, cells resistant to the SD of trastuzumab display some characteristics in common with the two other cultures (Figure 3C): indeed the TR-SKBR-3 cells attained a higher density in culture than did TS-SKBR-3 cells, but this was lower with respect to the untreated cells. However, the tendency to form spikes can be seen to have been maintained for a great of number TR-SKBR-3 cells in the field.

Panel of optical micrographs showing the effects of trastuzumab on the density and the morphology (see the insets) of the treated and untreated SKBR-3 cell populations at different times of culture with trastuzumab, from 72 h to 7 days from seeding. A: WT, trastuzumab untreated cells; B: TS, trastuzumab-sensitive cells; C: TR, trastuzumab-resistant cells. Bar=25 μm.
HER-2 levels in treated cells. To test the effect of trastuzumab treatment on the HER-2 expression levels we performed an immunodetection assay in the protein extracts from the TS-, TR- and WT-SKBR-3 cells. The analysis revealed that treatment induced a 2.3-fold decrease of the HER-2 level in both populations of treated SKBR-3 cells, compared to untreated cells (Figure 4).
SKBR-3 cell proteomics. Figure 5 shows a representative proteomic map of WT-SKBR-3. The protein identities are reported with the abbreviated name according to the Uniprot database (www.expasy.ch). A total of 293 proteins were identified, corresponding to 173 genes, due to the fact that many proteins are expressed by the cell in a variety of isoforms. Isoforms are indicated with letters in a progressive manner, starting with ‘a’ for lower pI or heavier MW. The 173 genes corresponding to the identified proteins are reported in Table II.
The known proteins were clusterized into 12 functional classes according to their major biological role in the current ontology database, namely: metabolic enzymes; cytoskeleton structure; cytoskeleton dynamics; detoxification; heat-shock/folding/chaperones; protein biosynthesis and degradation; calcium binding; nuclear proteins; ionic homeostasis; processing of metabolites and amino acids; proteins with extracellular activity; and nucleoside-related processes (Table I).
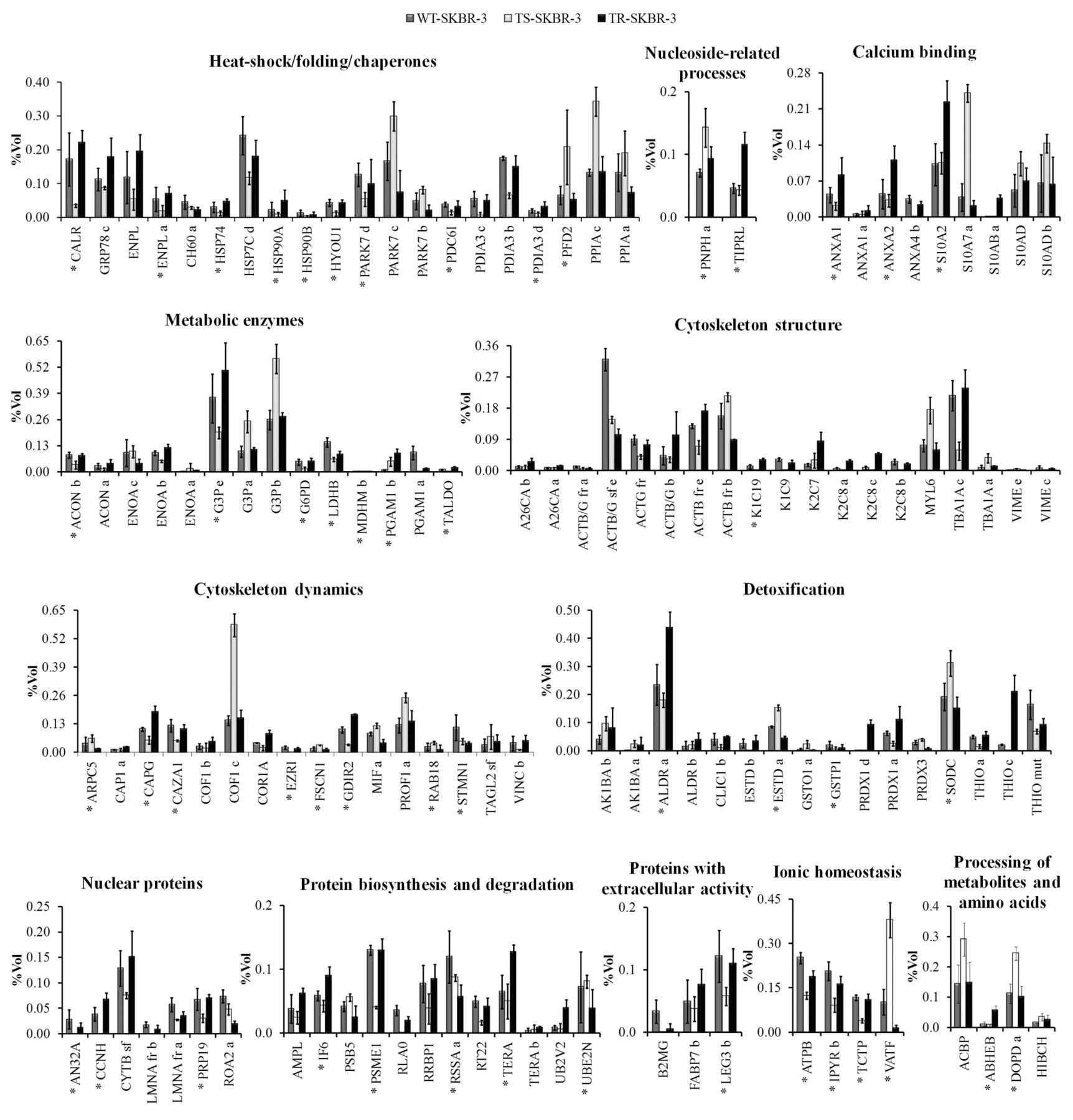
Proteomic profiling of trastuzumab-sensitive and -resistant SKBR-3 cells. The qualitative and quantitative analyses of the proteomic profiles (triplicate replicas, Figure 6) of untreated, sensitive and resistant cells revealed that the short-term treatment induced modulation of 84 out 293 identified proteins, corresponding to 66 genes. The effects exerted by the drug during the long-term treatment involved 41 proteins, corresponding to 33 genes. The modulated proteins are unevenly distributed over the 12 functional categories, with a major concentration in the groups of metabolic enzymes, cytoskeleton, heat-shock/folding/chaperone, calcium binding and detoxification.
The panel in Figure 7 reports the histograms of protein level variations following the short- and long-term trastuzumab treatment, as compared with these of the untreated cells. The quantitative level of each protein spot is given as %Vol, calculated as the integration of the intensity of the spot area, normalized by the sum of all spot volumes.
As shown by the graphs, there is a significant difference in the responses elicited by short- and long-term treatment. This is evident both in the number of responsive proteins and also in the quantitative expression of the proteins. The majority of proteins modulated appeared to be down-regulated: indeed out of the 84 responsive proteins in the TS-SKBR-3 cells, 60 displayed diminished intensity levels, while those for 24 increased with respect to the untreated cells. On the contrary, in the TR-SKBR-3 cells, out of the 41 modulated proteins, 24 appeared to be up-regulated and 17 down-regulated.
The group of up-regulated proteins in the TS-SKBR-3 cells comprises of: two calcium binding proteins (S10A7a and S10AD); proteins of the heat-shock/folding/chaperone cluster [two isoforms of protein DJ-1(PARK7) and two isoforms of Cyclophilin A (PPIA)]; several proteins belonging to cytoskeletal dynamics group [Cofilin-1 (COF1), Profilin-1 (PROF1), Fascin (FSCN1) and Ras-related protein Rab-18 (RAB18)]; proteins of the detoxification cluster [two isoforms of Aldo-keto reductase 1 B10 (AK1BA), Glutathione S-transferase omega-1 (GSTO1), Superoxide dismutase 1 (SODC)]; some isoforms of the metabolic enzymes [Glyceraldehyde-3-phosphate dehydrogenase (G3P) and alpha-Enolase (ENOA)] and others belonging to the nuclear proteins (Purine nucleoside phosphorylase, PNPH), to the ionic homeostasis cluster (V-type proton ATPase F, VATF) and to the metabolites and aminoacid processing proteins [Acyl-CoA-binding protein (ACBP) and D-dopachrome decarboxylase (DOPD)].
Among those down-regulated, it is worth noting the decreased level of some protein spots corresponding to some glycolytic enzymes. Among these, G3P is represented in our map by five protein spots. The more basic form, at pI 7.52, has been identified as the ‘canonical’ primary gene product, while the other spots at lower pI, from 7.09 to 7.44, represent protein isoforms. Comparison with the WT-SKBR-3 cells highlighted that in TS-SKBR-3 cells the canonical form was down-regulated by almost two-fold, while three of the more acidic isoforms appeared to be up-regulated. Similarly, other metabolic enzymes, L-lactate dehydrogenase B chain (LDHB), Transaldolase (TALDO), Glucose-6-phosphate 1-dehydrogenase (G6PD), and two isoforms of Aconitase (ACON) appeared to be down-regulated by about 50% in TS-SKBR-3 cells.

A: Western blot immunodetection of the Human Epidermal Growth Factor Receptor-2 (HER-2) expression in short- and long-term-treated cells (indicated as TS- and TR-SKBR-3 respectively) with respect to untreated (WT) cells. B: Histogram of the densitometric quantification of the immunoreactive bands. Trastuzumab administration induced a 2.2/2.3-fold downregulation of HER-2 expression in both TS and TR cells.
Interestingly, on short term there was a drastic down-regulation of several proteins belonging to the heat-shock/folding/chaperone proteins: Calreticulin (CALR), Hypoxia up-regulated protein 1, (HYOU1), Programmed cell death 6-interacting protein (PDC6I), three isoforms of Protein disulfide-isomerase (PDIA), Endoplasmin (ENPL), Heat shock protein HSP 90 alpha and beta, (HSP90A, HSP90B). Among these, ENPL, HSP90A and HSP90B are related to the HER-2 maturation pathway.
Within the cytoskeleton structural proteins, there was a drastic down-regulation of the epithelial cytokeratins K2C8, K1C19 and K1C9, and an evident reduction of some proteins involved in cytoskeletal dynamics, namely: Coronin-1A (COR1A), Ezrin (EZRI), Rho GDP-dissociation inhibitor 2 (GDIR2) and three acting capping proteins [Adenylyl cyclase-associated protein 1, (CAP1), Macrophage-capping protein (CAPG) and F-actin-capping protein subunit alpha-1, (CAZA1)].

Representative proteomic analysis of SKBR-3 cells. Protein separation was performed by Immobilized pH Gradient (IPG) gel strips (18 cm, 3.5-10 non-linear gradient) followed by a gradient (9-16%T) sodium dodecyl sulphate-polyAcrylamide gel electrophoresis (SDS-PAGE). The 293 identified protein spots are labelled with the Uniprot AC number.
Trastuzumab effects on TS-SKBR-3 cells include also the down-regulation of proteins involved in disparate functions, such as: detoxification [Chloride intracellular channel protein 1 (CLIC1) and three isoforms of Thioredoxin (THIO)]; the nuclear protein cluster [Acidic nuclear phosphoprotein pp32 (AN32A), Cyclin H (CCNH), two Prelamin-A (LMNA) fragments and Nuclear matrix protein 200 (PRP19)]; protein biosynthesis and degradation [Proteasome activator complex subunit 1 (PSME1), 60S acidic ribosomal protein P0 (RLA0), Ribosome-binding protein 1 (RRBP1) and Ribosomal protein S22 (RT22)]; and ionic homeostasis [ATP synthase subunit beta (ATPB), Inorganic pyrophosphatase (IPYR) and Fortilin (TCTP)].

Miniatures of triplicate 2D silver-stained gels of untreated (WT)- (A), trastuzumab-sensitive (TS)- (B) and trastuzumab-resistant (TR)-SKBR-3 cells' (C) protein extracts.
The comparison of the trastuzumab-resistant cells with the untreated ones revealed a reversion of the responses initially observed in the trastuzumab-sensitive cells. This trend is fully elucidated in the graph in Figure 7, which clearly shows that the expression of about 60% of the proteins modulated on short-term treatment (49 out of 84) reverted to the levels of the untreated cells in the resistant, long-term treated cells.
Finally, we found a small group of 24 protein spots, corresponding to 21 genes, unaffected by the short treatment but responsive to the prolonged administration, 18 of which were up-regulated and six down-regulated. The latter group comprises an isoform of ENOA, Peroxiredoxin-3 (PRDX3), Proteasome subunit beta type-5 (PSB5), 40S ribosomal protein SA (RSSA) and Ubiquitin carrier protein N (UBE2N). Among the 18 proteins positively affected by long exposure to trastuzumab, the remarkable increase of an isoform of Peroxiredoxin-1 (PRDX1) and of the acidic form of S10AB is noteworthy. Less evident but equally interesting is the increase in the expression of other proteins, namely: Annexin-A1 (ANXA1), Annexin-A2 (ANXA2), CAP1, 78 kDa glucose-regulated protein (GRP78) and S10A2.

The panel summarizes the protein level variations (cut off ≥2-fold) among sensitive cells (TS-SKBR-3), resistant cells (TR-SKBR-3) and untreated cells (WT). The protein names, abbreviated according to the Uniprot database are given on the abscissa; on the ordinate the quantitative level of each protein spot is given as %volume (%Vol) derived from three independent replicates. The protein distribution in the graph is performed according to the functional categories. The isoforms are indicated with alphabetical letters and the primary protein (canonical form, according to the theoretical MW and PI) is labeled with an asterisk. For each histogram error bars indicate the standard deviation from the mean of triplicate determinations.
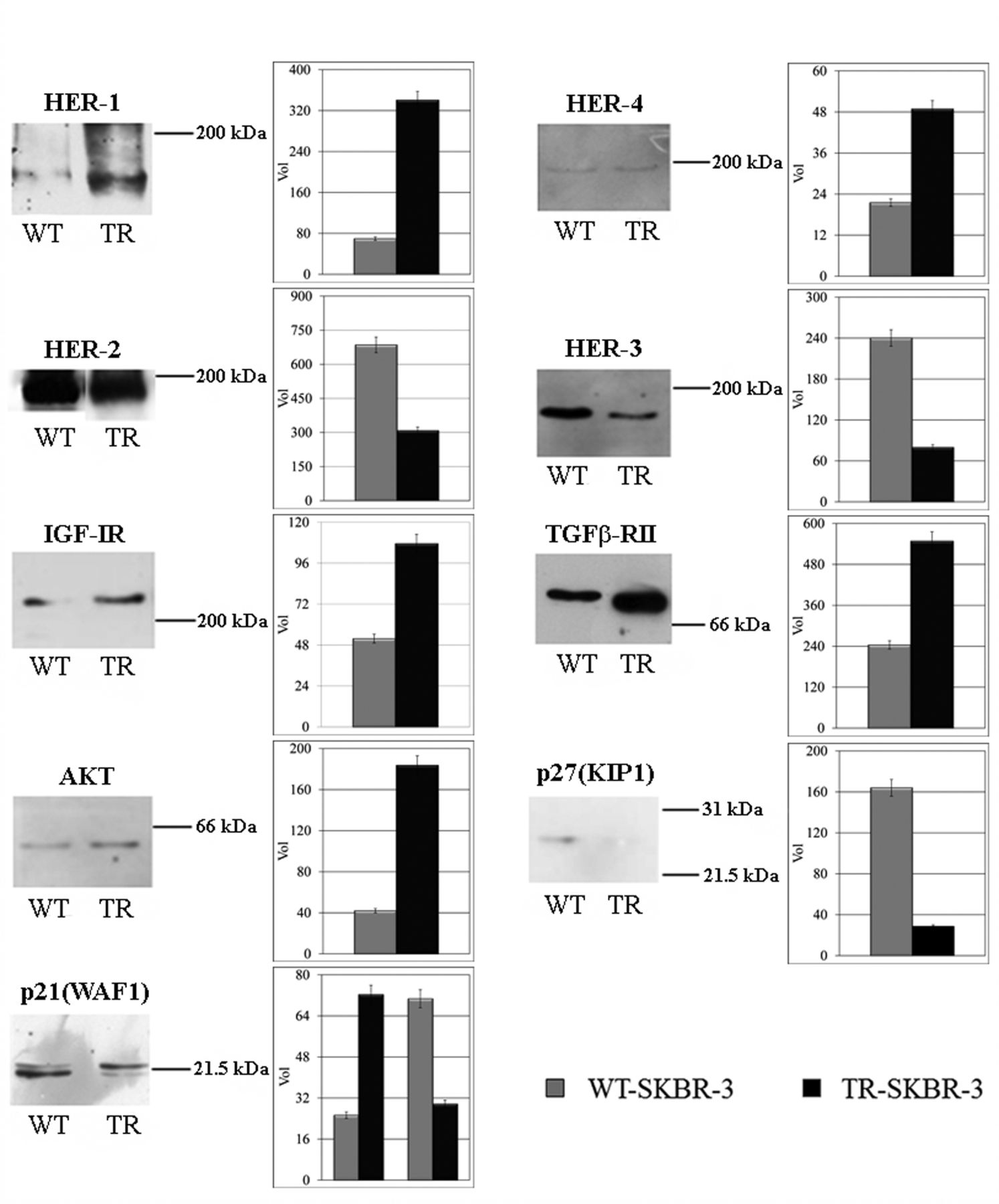
Western blot immunodetection of putative candidates involved in the acquisition of trastuzumab resistance. The respective intensity level of each reactive band in the untreated (WT) and resistant cells (TR) is reported in the histograms. [Human Epidermal Growth Factor Receptors 1-4 (HER1-4), Insulin growth factor 1 receptor (IGF-IR), Transforming growth factor-beta receptor type II (TGFB-RII), RAC-alpha serine/threonine-protein kinase (AKT)].
Immunodetection of the main HER-2 pathway co-actors. Due to the drastic reversion of the responses to trastuzumab of the resistant cells, we investigated about the major co-actors of the HER-2 pathways, probably related to cellular mechanisms underlying the resistance to trastuzumab, through western blot immunodetection.
Figure 8 reports the comparative levels of the HER family members, of two other tyrosin-kinase (TK) receptors (IGF-IR and TGFB-RII) and three downstream proteins [AKT, p27(KIP1) and p21(WAF1)]. Interestingly, trastuzumab resistance is related to the increment of two other HER family members receptors, i.e. HER-1 (5-fold) and HER-4 (2.2-fold), of the two other TK receptors, IGF-IR (2-fold) and TGFB-RII (2.2-fold), and of the downstream kinase AKT (4.5-fold). The treatment also induced the drastic down-regulation of HER-3 receptor (3-fold) and of p27(KIP1) protein (4.5-fold), a cell-cycle modulator. Moreover, the immunodetection of the protein p21(WAF1), which participates in p53-mediated cell proliferation control, revealed the presence of two immunoreactive bands: the higher MW band is identifiable as the phosphorylated form of p21, whereas the lower MW band may represent the non-phosphorylated protein. Our results indicate that prolonged treatment with trastuzumab induce a 3-fold increase of the higher MW band and a 2.5-fold decrease of the lower MW band.

Histograms representing the expression levels of the 28 proteins related to the regulation of cell death affected by short- and long-term treatment with trastuzumab. The protein names, abbreviated according to the Uniprot database are given on the abscissa; on the ordinate the quantitative level of each protein spot is given as %volume (%Vol) derived from three independent replicates. The isoforms are indicated with alphabetical letters and the primary protein (canonical form, according to the theoretical MW and PI) is labeled with an asterisk. For each histogram error bars indicate the standard deviation from the mean of triplicate determinations.
Discussion
The present study is, to our knowledge, the first large-scale proteomic identification of protein clusters involved in the responses to trastuzumab treatment in vitro. As a cellular model, we used the well-characterized SKBR-3 cells overexpressing the drug target HER-2. Cells were submitted to short- and long-term treatment, in order to obtain sensitive and resistant clones. The comparative proteomic profiling of the untreated and treated SKBR-3 clones highlighted interesting differences in both the number and expression levels of identified proteins. The proteins mainly involved in drug responses belong to the categories of metabolic enzymes, cytoskeleton, detoxification, heat-shock/folding/chaperones, protein biosynthesis and degradation and calcium binding.
In some instance, it was observed that when more isoforms of the same protein were present, these were not always equally affected by the treatment. A peculiar example is G3P which is present in our map in five isoforms. One of them, representing the primary gene product since it was the most intense and the more basic among the group, appeared to be the only one significantly repressed by trastuzumab. This result suggests that one of the effects of trastuzumab treatment may be the down-regulation of glycolytic enzymes, which in neoplastic cells, especially HER-2-positive cells, are overexpressed according to the well-known Warburg effect (3). More interestingly, this drug effect was observed under short-term treatment, while it appears to have been lost in the long-term-treated cells. The same result, even though at lower levels, was observed for other metabolic enzymes (LDHB, TALDO, G6PD, and two isoforms of ACON).
The observed involvement of several proteins related to HER-2 maturation and HER-2 pathways was also of note. Among these, nine cytoskeletal proteins (five genes) and three HSPs (two genes) were mostly represented. HSPs included two isoforms of Endoplasmin (ENPL) (also called GRP94) and HSP90A, which are chaperones involved in cancer progression at multiple levels. In particular, HSP90A and its endoplasmic reticulum homolog ENPL, are known as ‘capacitors for evolution’ for their capacity to stabilize the conformations of mutated forms of oncogenic proteins arising during neoplastic transformation (29).
On the other hand, it must be recalled that probably each of the modulated proteins, in addition to their canonical primary function, are able to perform, or to participate in, several additional roles in the life and death of the cell. In particular, at least 28 of the differentially expressed proteins, each included in a specific cluster, are involved in the regulation of apoptosis (Figure 9). Among these, two proteins deserve particular attention. The first is COF1, a cytoskeleton-associated protein, which was 2.5-fold increased in the short-term treatment, while its expression was at basal levels in the resistant cell clones. This seems relevant when considering the additional cellular functions of COF1 unrelated to actin-assembly regulation. Among these, its mitochondrial translocation and induction of cytochrome c release in the initiation phase of apoptosis (30) and its chaperoning of actin to the nucleus to enable nuclear functions of actin (31) are among the most crucial cellular roles. Hence, it can be hypothesized that Cofilin may be one of the key players in trastuzumab treatment, whose function is probably lost in resistant cells.
The other protein which was significantly incremented in the short-term-treated cells and reduced in the long-term-treated cells is the Cyclophilin A (PPIA), which is primarily involved in the regulation of protein conformation. Several groups have reported that overexpression of PPIA protects cancer cells against cellular stress, including hypoxia and cisplatin treatment, through its antioxidant function (32-35), in general suggesting a positive function of PPIA toward progression in many solid tumors.
The fact that 18 proteins appear to be overexpressed only in the long-term-treated cells is also of interest. Two of these were de novo expressed, namely the acidic isoform of S10AB and the more basic isoform of PRDX1. The latter, besides its pivotal role in stress responses, is involved in multiple pathways of cell metabolism and cytoskeletal dynamics (36, 37). This suggests a novel role for PRDX1 as a possible common node downstream of multiple trastuzumab-resistance pathways. The other de novo expressed protein is S10AB, a small calcium-binding protein involved in multiple cell pathways regarding cell proliferation, differentiation and cancer. S10AB represents a candidate prognostic marker for a wide number of tumors, both for its correlation with histological grade and for its dramatic increase during the progression of many tumor types (38). Moreover, other S100 family proteins were reported to be closely related to cancer progression (39): S10A7 (40), S10A4 (41), S10A18 (42), S10A9 (43) and S10A8 (44).
Most relevant is the evidence that long-term treatment with trastuzumab induces a significant decrement not only of HER-2 (its primary target), but also of its heterologous partner HER-3. Indeed, the heterodimer HER-2-HER-3 plays a pivotal role in tumour progression through de-regulation of the G1-S transition and through favouring uncontrolled cell proliferation (45).
In contrast, resistance to the drug is associated with a significant increase of the expression levels of other HER family receptors, HER-1 and HER-4, as well as of other TK receptors, IGF-IR and TGFB-RII, as demonstrated by the immunodetection assays.
Functional classes of the 293 proteins identified in the SKBR-3 proteome, grouped according to the major biological function, as reported by the current ontology databases. Number of proteins represents the total proteomic profile, independent of drug-induced variations.
It is known that IGF-IR, TGFB-RII and HER-2 may cross-talk by common downstream pathways, such as Mitogen-activated protein kinase (MAPK) and Phosphoinositide 3-kinase (PI3K) (46, 47). On this basis, the observed increment of IGF-IR and of TGFB-RII in long-term responses to trastuzumab could trigger survival and mitogenic pathways in spite of HER-2 targeting by the drug. This is also supported by the 4.5-fold increase of AKT expression in the resistant cells. It has been reported that the deregulation of the AKT-PTEN (Phosphatidylinositol 3,4,5-trisphosphate 3-phosphatase) network may be related to the evasion of the pharmacological action of trastuzumab (11, 48). Once activated, AKT stimulates cell survival, cell-cycle progression, angiogenic pathways and responses to environmental stress (e.g. hypoxia and nutrient depletion).
Finally, our results indicate a marked down-regulation of the expression of p21 and p27 in cells treated long-term with trastuzumab. p27 induces cell-cycle arrest in the G1 phase by inhibiting the formation of the complexes Cyclin-dependent kinase 2 (CDK2)-cyclin E and CDK2-cyclin A in the presence of DNA alteration and damage. It has been shown that the p27 protein levels decrease in a wide number of cancer types as a result of its interaction with a component of the signalosome complex and subsequent degradation via the proteasome (49). With regards to p21 protein, its action is dependent on its subcellular localization: as a nuclear protein it exerts an antiproliferative role, while as a cytoplasmic protein, it has anti-apoptotic activity. p21 cellular destiny is closely related to the AKT kinase activity and is associated with poor prognosis in patients with neoplasms (50).
Lists of the 173 genes encoding for the 293 protein spots identified in the 2D maps of the proteome of SKBR-3 cells. The table reports: the protein name, Uniprot AC number, gene number and number of identified isoforms.
Conclusion
The present data suggest that the primary effect of trastuzumab at the SD is the slowing down of the growth rate of sensitive SKBR-3 cells in concomitance with metabolic regression, as judged by the down-regulation of key metabolic enzymes belonging in the glycolytic pathway, and the decrement of several proteins related to HER-2 maturation and HER-2 pathways.
Once insensitivity to trastuzumab is achieved, following prolonged administration, the surviving cell clones appear to revert their phenotype to match that of untreated parental cells. Moreover, prolonged exposure to the drug induces the overexpression of alternative TK receptors and of downstream effectors [(AKT), p21(WAF1)] which may compensate for the HER-2 block. The de novo expression of two proteins with crucial role in tumour progression, the acidic isoform of S10AB and the more basic isoform of PRDX1, may also play a crucial role in the resistant cell progression.
Conclusively, the large-scale proteomic approach applied to the study of the effects induced in target cells by trastuzumab highlights new complex scenarios, as it could be expected when molecular perturbation is induced into cells. Many interrelated pathways have been found to be involved in the responses to trastuzumab. We suggest that the present results may improve the knowledge of the molecular effects of trastuzumab and could be useful for understanding resistance mechanisms while improving the efficacy of the treatment response.
Acknowledgements
The present work was supported in part by Ph.D. funds to G.D. by the University of Palermo, and in part by Italian Government contributions to C.OB.S. as a distinguished non-profit research association [‘5x1000’ contribution, as provided in paragraphs 337-340 of the Italian Government Finance Act for 2006 (Law of 23 December 2005, no. 266)].
- Received November 12, 2012.
- Revision received December 10, 2012.
- Accepted December 11, 2012.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.