


Abstract
Citric acid is an alpha-hydroxyacid (AHA) widely used in cosmetic dermatology and skincare products. However, there is concern regarding its safety for the skin. In this study, we investigated the cytotoxic effects of citric acid on the human keratinocyte cell line HaCaT. HaCaT cells were treated with citric acid at 2.5-12.5 mM for different time periods. Cell-cycle arrest and apoptosis were investigated by 4,6-diamidino-2-phenylindole dihydrochloride (DAPI) staining, flow cytometry, western blot and confocal microscopy. Citric acid not only inhibited proliferation of HaCaT cells in a dose-dependent manner, but also induced apoptosis and cell cycle-arrest at the G2/M phase (before 24 h) and S phase (after 24 h). Citric acid increased the level of Bcl-2-associated X protein (BAX) and reduced the levels of B-cell lymphoma-2 (BCL-2), B-cell lymphoma-extra large (BCL-XL) and activated caspase-9 and caspase-3, which subsequently induced apoptosis via caspase-dependent and caspase-independent pathways. Citric acid also activated death receptors and increased the levels of caspase-8, activated BH3 interacting-domain death agonist (BID) protein, Apoptosis-inducing factor (AIF), and Endonuclease G (EndoG). Therefore, citric acid induces apoptosis through the mitochondrial pathway in the human keratinocyte cell line HaCaT. The study results suggest that citric acid is cytotoxic to HaCaT cells via induction of apoptosis and cell-cycle arrest in vitro.
Citric acid is a type of α-hydroxy acid (AHA) which has been widely used in cosmetic products for years (1-4). AHAs attract customers with their potential rejuvenating ability to reduce wrinkles, spots, and other signs of photo-aging (5). Citric acid at a concentration of 20% increased the thickness of epidermis and increased glycosamino-glycans on sun-damaged skin (6, 7). Citric acid also induced collagen I and pro-collagen II proliferation (5). Behind these rejuvenating effects, the Food and Drug Administration voiced a particular concern about AHAs because of their ability to penetrate the skin barrier and cause skin irritations (5, 8). Being concerned about the safety of AHAs, especially in long-term application to the skin, we are interested in exploring the anti-proliferative and apoptotic effects of AHAs on human keratinocytes (9, 10). We evaluated the cytotoxic effects of glycolic acid (10-12) and lactic acid (13, 14) in previous studies, but the in vitro effects of citric acid remain unclear. In this study, we investigated the effects of citric acid in the human keratinocyte cell line (HaCaT).
Materials and Methods
Chemicals and reagents. Citric acid, propidium iodide (PI), RNase A, and Triton X-100 were obtained from Sigma Chemical Co. (St. Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM), penicillin-streptomycin, trypsin-EDTA, fetal bovine serum (FBS) and L-glutamine were obtained from GIBCO®/Invitrogen (Carlsbad, CA, USA). Dichloro-dihydro-fluorescein diacetate (DCFH-DA), 3,3-Dihexyloxacarbocyanine iodide (DiOC6) and 1-[Amino-5-(2,7-dichloro-6-acetomethoxy-3-oxo-3H-xanthen-9-yl)phenoxy]-2-(2’-amino-5’-methylphenoxy)ethane-N,N,N’,N’-tetraacetic acid, pentaacetoxy-methyl ester (Fluo-3/AM) were obtained from Molecular Probes/Invitrogen (Eugene, OR, USA). Caspase-3 activity assay kit was purchased from Roche Diagnostics (Mannheim, Germany). All chemicals used were reagent grade.

Morphological changes in citric acid-treated HaCaT cells were examined and photographed under phase-contrast microscopy at 24 h (A). The percentage of viable cells was measured by flow cytometric assay at 24 h (B). Data are presented as the mean±SD of three independent experiments. *p<0.05, ***p<0.001 significantly different compared with the control treatment.
Human immortalized keratinocyte cell line HaCaT. Cultures of HaCaT cells were kindly provided by Dr. Norbert E. Fusenig (Institute of Biochemistry, German Cancer Research Center) and were grown on DMEM supplemented with 1% L-glutamine, 25 mM HEPES, 10% FBS and 1% penicillin-streptomycin at 37°C in a humidified incubator with 5% CO2 atmosphere (10, 14).
Assessment of cell morphological changes and viability. HaCaT cells at the density of 2×105 cells/well were placed onto 12-well plates and incubated at 37°C for 24 h before being treated with different concentrations of citric acid for 24 h; 0.5% of DMSO (vehicle) was used as a control. At the end of incubation, cells were examined and photographed under phase-contrast microscopy for determination of morphological changes. Cells (1×105 cells per sample) were then centrifuged at 1000 ×g for 5 min, and cell pellets were dissolved with 0.5 ml of Phosphate buffered saline (PBS) containing 5 μg/ml PI and viable cells were determined by using a flow cytometer (Becton-Dickinson, San Jose, CA, USA) for determination of viable cells as previously described (10, 14, 15).
Assessment of cell-cycle distribution and apoptosis by flow cytometry. HaCaT cells treated with citric acid at 12.5 μM were incubated for different time periods up to 72 h before the cells were harvested by centrifugation. The cells were fixed gently by 70% ethanol at 4°C overnight and were then re-suspended in PBS containing 40 mg/ml PI, 0.1 mg/ml RNase and 0.1% Triton X-100 in a dark room. After incubation at 37°C for 30 min, the cell-cycle distribution and apoptosis were analyzed with a flow cytometer (15).
4,6-Diamidino-2-phenylindole dihydrochloride (DAPI) staining for apoptotic cells. Cells at a density of 2×105 cells/well were placed onto 6-well plates and treated with citric acid (0, 2.5, 5, 7.5, 10 and 12.5 mM) for 24 h before cells from each treatment were isolated for DAPI staining as described previously (10, 14). After staining, the cells were examined and photographed using a fluorescence microscope.
Detection of the changes of levels of reactive oxygen species (ROS), Ca2+, and mitochondrial membrane potential (MMP) in HaCaT cells by flow cytometry. The levels of ROS, Ca2+, and MMP of the HaCaT cells were determined by flow cytometry. HaCaT cells were treated with or without citric acid (12.5 mM) for different time periods (0, 1, 6, 12, 24, and 48 h). The cells were harvested and washed twice, resuspended in 10 μM DCFH-DA (Sigma-Aldrich) and incubated at 37°C for 30 min. The level of ROS was then analyzed by flow cytometry. DCFH-DA is an oxidation-sensitive fluorescent probe. In the presence of ROS, DCFH-DA is subsequently converted to DCF and emits a green fluorescent signal detected by flow cytometry (14, 15).
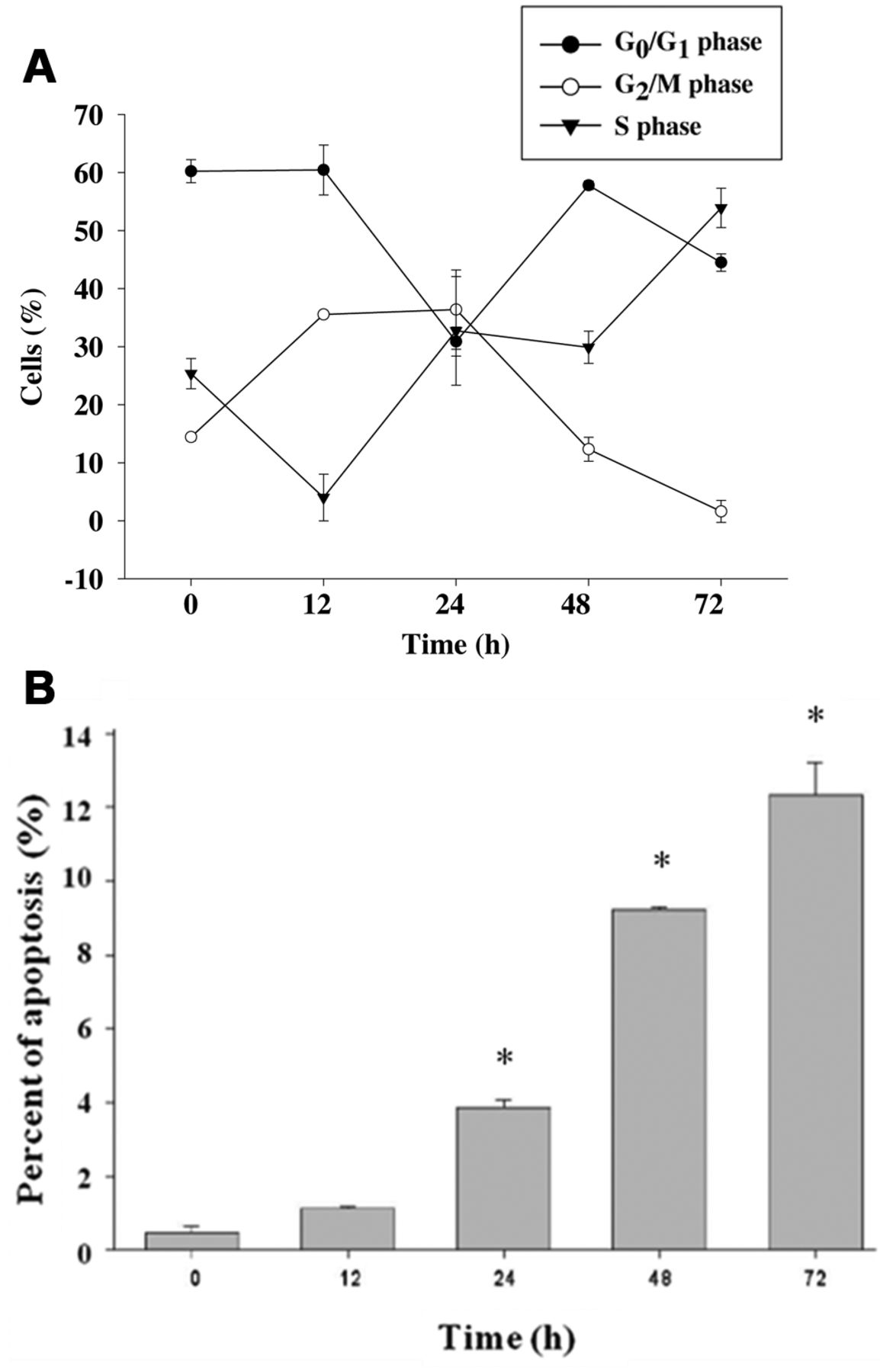
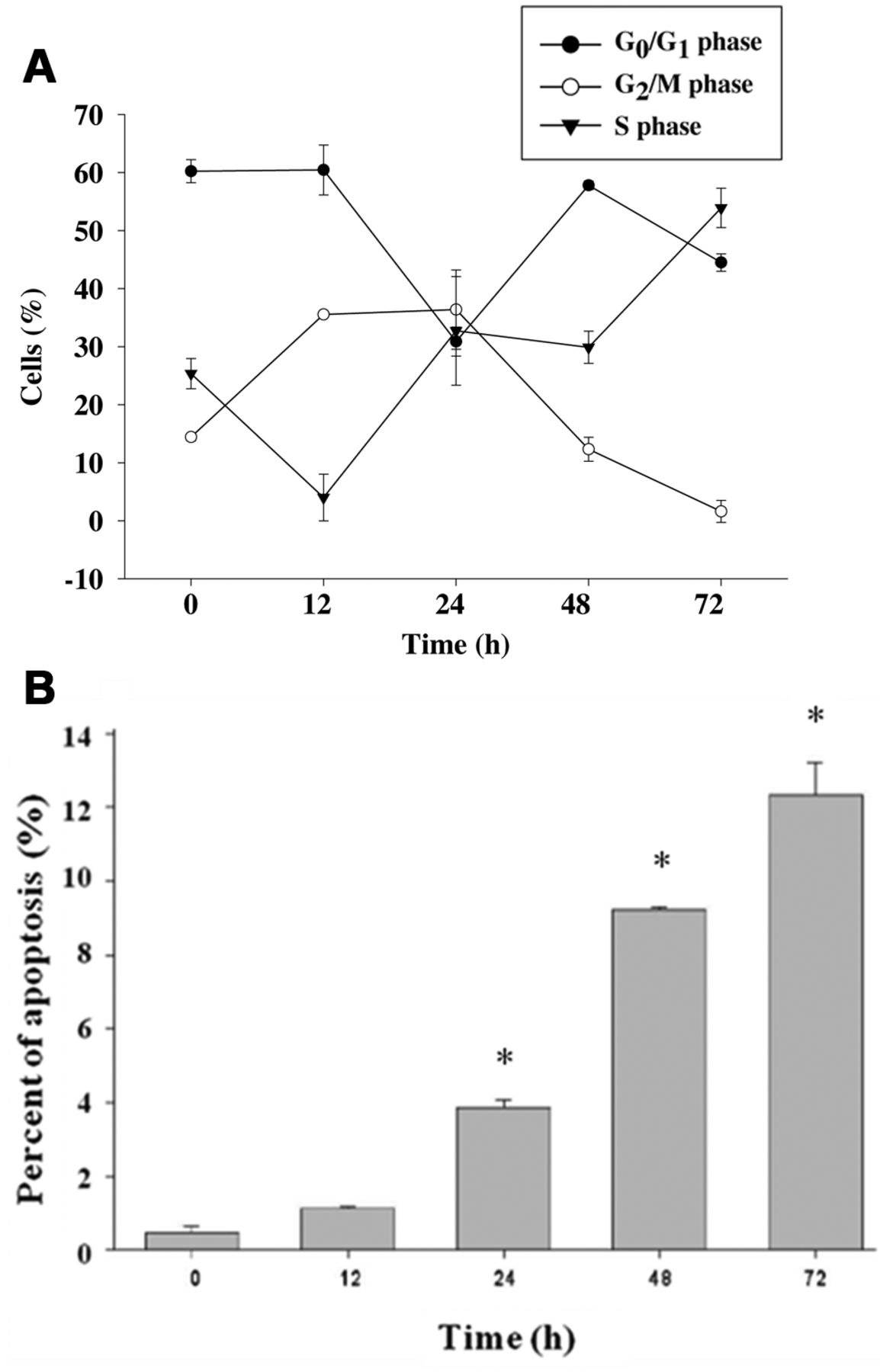
The effects of citric acid on cell-cycle distribution in HaCaT cells. Cells were treated with 12.5 mM of citric acid for 0, 12, 24, 48, and 72 h (A). The cell-cycle distribution and sub-G1 group (apoptosis phase) were determined using flow cytometric analysis (B) and were obtained from three independent experiments with similar results. Data represent the mean±SD of the results from three experiments (n=3). *p<0.05 compared with the control treatment.
Cells were treated with citric acid at 12.5 mM for different times (0, 0.5, 1, 6, 12, 24, and 48 h) before detection of the changes of Ca2+ concentration. The cells were harvested and washed twice, re-suspended in a buffer containing Indo-1-AM (3 mg/ml; Calbiochem, La Jolla, CA, USA) and incubated at 37°C for 30 min. The levels of Ca2+ was then analyzed by flow cytometry (16).
For detecting the changes of MMP in HaCaT cells, HaCaT cells were treated with or without citric acid (12.5 mM) for different time periods (0, 0.5, 1, 6, 12, 24, and 48 h). The cells were then harvested and washed twice, resuspended in 500 μl of DiOC6 (4 mol/l) and incubated at 37°C for 30 min, and subsequently analyzed by flow cytometry (17, 18).
Assays for caspase-3, -8 and -9 activities. Cells (total 2×105 cells) in 12-well plates were incubated with 12.5 mM of citric acid and 0.1% DMSO (vehicle-control) for 0, 24, and 48 h. Cells were harvested, and add 50 μl of 10 μM substrate solution (PhiPhiLux®-G1D2, CaspaLux®-8-L1D2 and CaspaLux®-9-M1D2 kits, OncoImmunin, Inc., Gaithersburg, MD, USA) were then added to the cell pellet. Cells were then incubated at 37°C for 60 min and were washed once by adding 500 μl of fresh PBS. The caspase-3, -8 and -9 activities were analyzed by flow cytometry, as previously described (18-20).

HaCaT cells were incubated with citric acid at different concentrations for 24 h. There was an increase in the number of higher intensity 4,6-diamidino-2-phenylindole dihydrochloride (DAPI)-staining cells and fragmented nuclei at higher concentrations of citric acid (scale bar=20 μm, fluorescence microscopy, ×200).
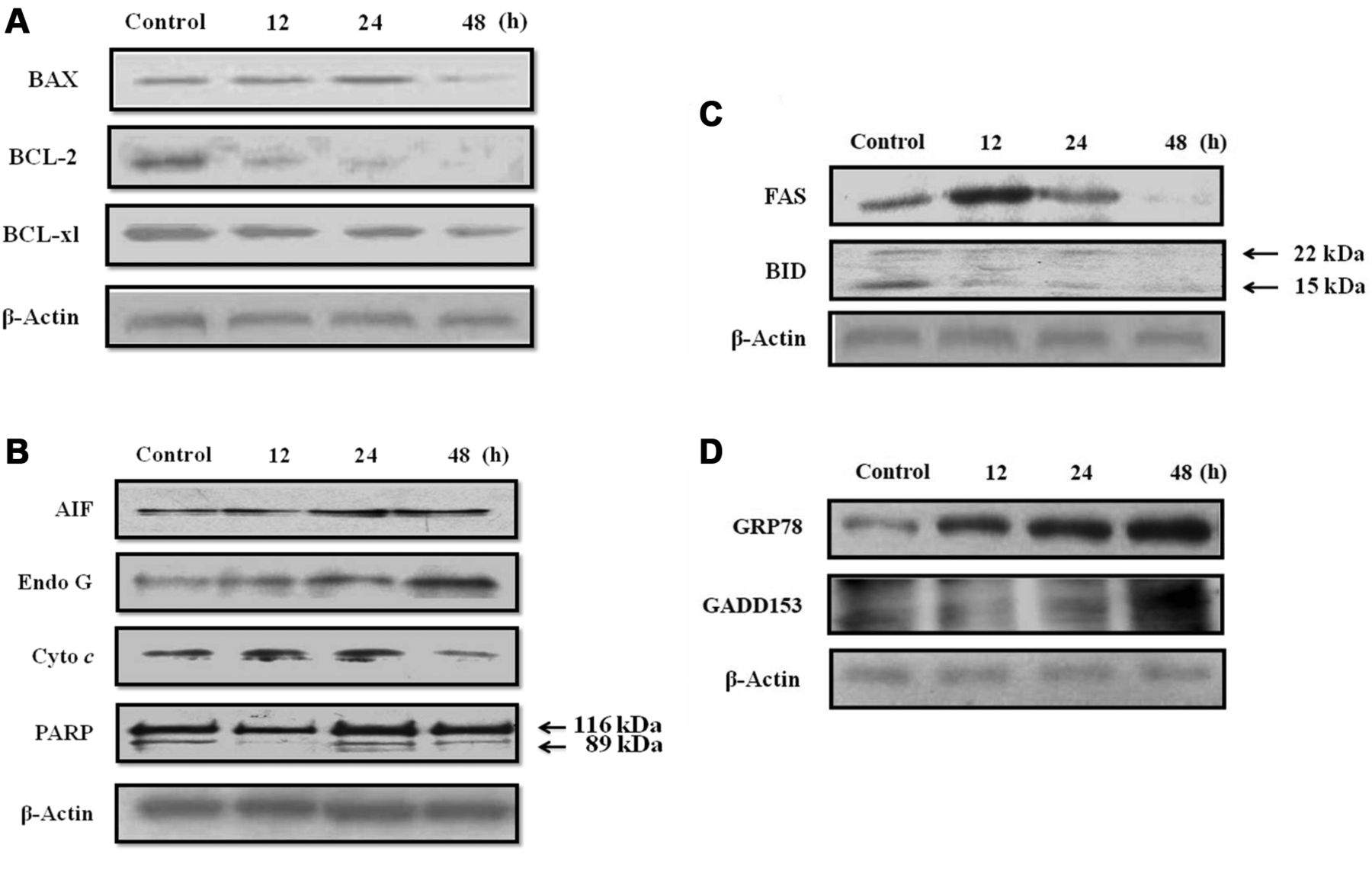
Apoptosis-associated proteins determined by western blotting. HaCaT cells at a density of 1×106 cells/ml cells in 6-well plates were treated with 12.5 mM of citric acid for 0, 12, 24 and 48 h. Cells were harvested from each treatment by centrifugation for total proteins determination as Bio-Rad kit for western blotting (Bio-Rad Laboratories, Inc., Berkeley, CA, USA) The protein levels of Bcl-2-associated X protein (BAX), B-cell lymphoma 2 (BCL-2), B-cell lymphoma-extra large (BCL-XL), apoptosis-inducing factor (AIF), endonuclease G (EndoG), cytochrome c, Poly ADP ribose polymerase (PARP), TNF receptor superfamily, member 6 (FAS), BH3 interacting-domain death agonist (BID), 78 kDa glucose-regulated protein (GRP78), and DNA-damage-inducible transcript 3 (GADD153) were examined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting, as described previously (21-23).
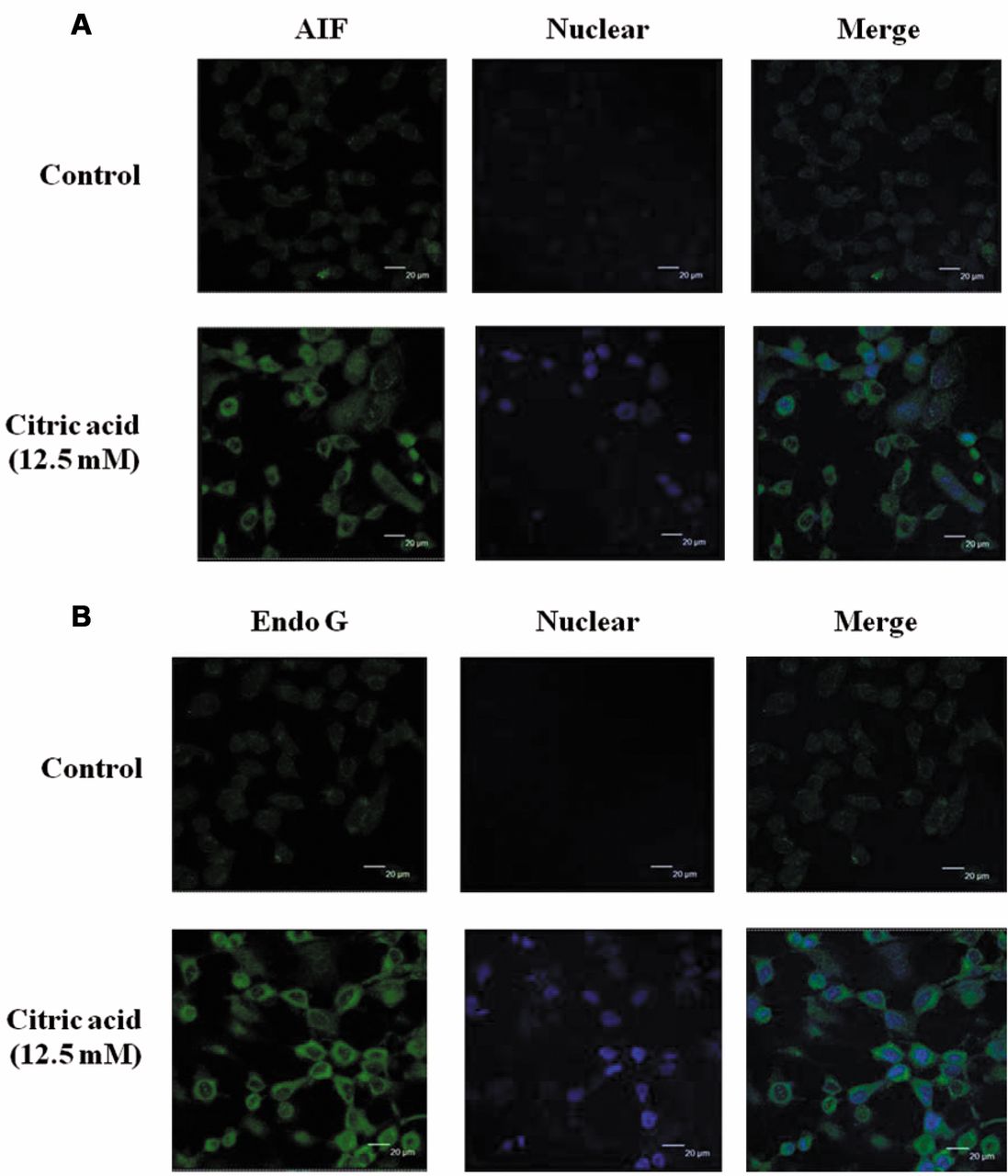
Confocal microscopy to detect AIF, EndoG and cytochrome c in HaCaT cells exposed to citric acid. HaCaT cells were gently fixed in buffer (400 mM sucrose, 2 mM EGTA, 4% paraformaldehyde in PBS) for 15 min, and then the cells were permeabilized with 0.1% Triton-X in PBS for 5 min. After rinsing cells with PBS, blocking solution [1% (BSA) in PBS] was added for 1 h, and cells subsequently stained with a mouse monoclonal antibody against EndoG (at 1:200 dilution), goat polyclonal antibody against AIF (at 1:200 dilution), or rabbit polyclonal antibody against cytochrome c (at 1:200 dilution) at 4°C overnight. After rinsing with PBS, cells were stained with a Fluorescein isothiocyanate (FITC)-conjugated fluorescent secondary antibody (at 1:200 dilution; all antibodies from Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 2 h at room temperature. The cells were rinsed with PBS containing 1 μg/ml PI to act as a nuclear counter stain, and then washed with ice-cold PBS. After mounted with a mounting medium, the cells were examined and photo-micrographed under a Leica TCS SP2 Confocal Spectral Microscope, as described previously (15, 23, 24).
Statistical analysis. The results are presented as the mean±S.E.M, and the difference between the citric acid-treated and control groups was analyzed by Student's t-test, a probability of *p<0.05, ***p<0.001 being considered significant.

Citric acid affected the mitochondrial membrane potential (MMP), reactive oxygen species (ROS), Ca2+ accumulation in HaCaT cells. Cells were treated with 12.5 mM of citric acid for different time periods. All samples were analyzed by flow cytometric assay as described in Materials and Methods. Data are presented as the mean±SD of three independent experiments. *p<0.05, ***p<0.001, significantly different compared to control treatment.
Results
Citric acid inhibited cell viability and morphological changes in HaCaT cells. After HaCaT cells were treated with various concentrations of citric acid for 24 h, the cell morphological changes also occurred from citric acid-treated cells and the results are shown in Figure 1A. The cell viability significantly decreased in a dose-dependent manner (Figure 1B). The half-maximal inhibitory concentration (IC50) value of citric acid was approximately 12.5 mM on 24-h exposure. Thus, this concentration of citric acid was applied for all subsequent experiments.
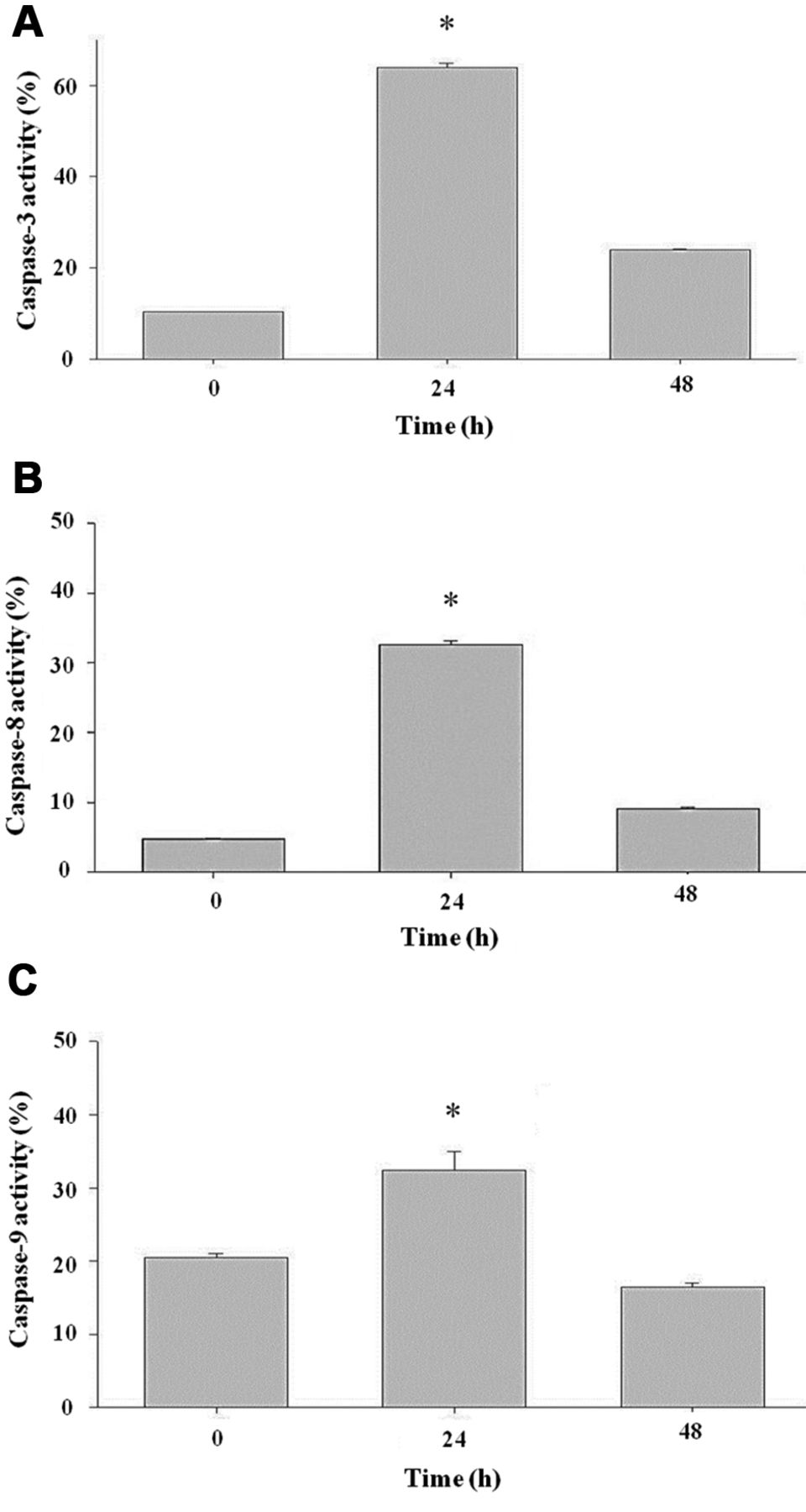
Citric acid induced caspase-3, -8 and -9 activities. Cells were treated with 12.5 mM of citric acid for different time periods. All samples were analyzed by flow cytometric assay as described in Materials and Methods. Data are presented as the mean±SD of three independent experiments. *p<0.05, ***p<0.001, significantly different compared with the control treatment.
Citric acid induced cell-cycle arrest and apoptosis of HaCaT cells. Flow cytometry disclosed that treatment with citric acid at 12.5 mM increased the proportion of cells at G2/M phases before 24 h and increased the proportion of cells at S phase after 24 h (Figure 2A). Citric acid also induced a distinct sub-G1 peak, which represents the population of apoptotic cells. Higher concentrations of citric acid increased the proportion of apoptosis, as shown in Figure 2B.

Western blots demonstrated the increase of FAS, BAX, BID, AIF, EndoG, cytochrome c, PARP, GADD153, GRP78 and caspase-3, -8, -9, and decrease of BCL-2 and BCL-Xl in HaCaT cells exposed to citric acid at 12.5 mM for different periods.
Apoptotic cells examined by DAPI staining. Citric acid-treated cells were variable in cell size, with fewer and less fluorescent cells than that of the control samples (Figure 3). Moreover, after treatment with citric acid, cells displayed chromosomal condensation and formation of apoptotic bodies (Figure 3). These effects occurred in a dose-dependent manner.
Citric acid reduced the MMP, increased levels of ROS and caused Ca2+ accumulation in HaCaT cells. To investigate whether citric acid induces cell death through the dysfunction of mitochondria, ROS, and Ca2+ accumulation, cells were harvested after exposure to citric acid and were assayed by flow cytometry. The results, shown in Figure 4A-C, indicate that citric acid reduced the MMP (Figure 4A), promoted the production of ROS (Figure 4B) and led to Ca2+ accumulation (Figure 4C) in a time-dependent manner. In addition, caspase-3, capase-8 and -9 activities were promoted by citric acid (Figure 5) in HaCaT cells.
Effect of Citric acid on the expression of apoptosis-related proteins in HaCaT cells. HaCaT cells were treated with citric acid at 12.5 mM for different time periods. Figure 6 shows that citric acid promoted the expression of apoptosis-related proteins as FAS, BAX, cytochrome c, PARP, AIF, EndoG, GRP78, and GADD153, but reduced the levels of BCL-2, BCL-xL, and BID.
Confocal microscopy showing the release of AIF, EndoG and cytochrome c in HaCaT cells exposed to citric acid. To investigate the mechanism underlying apoptosis induced by citric acid, we tested the effects of citric acid on AIF and EndoG localizations in HaCaT cells. Thus, our hypothesis was that citric acid-induced apoptosis of HaCaT cells might be mediated through mitochondria-dependent pathways releasing AIF and EndoG. The results from confocal laser microscopy indicated that citric acid promoted the release of AIF (Figure 7A) and EndoG (Figure 7B) to nuclei in HaCaT cells.

Confocal microscopy showed a marked increase in the intra-cytosolic release of AIF and EndoG; in addition, we found the translocation of AIF and EndoG from the cytoplasm to the nucleus in HaCaT cells treated with citric acid at 12.5 mM for 24 h. AIF, EndoG, and cytochrome c were stained by FITC fluorescence green, whereas nucleus stained by DAPI fluoresce blue (scale bar=20 μm).
Discussion
Although several reports have outlined the safety concern regarding AHAs, the exact molecular mechanisms of the action of citric acid on human epidermal keratinocytes remain unclear (7). According to the recommendation of the Department of Health, R.O.C., the concentration of AHAs must be less than 10%, at pH3.5 for consumers (2). However, chemical peeling agents used in hospitals and local practitioner's clinics were found to contain higher concentrations of AHAs (20-70%) (25). Regarding cell toxicity of citric acid to skin, we herein investigated citric acid effect on human epidermal keratinocytes (HaCaT cells). The results indicated that citric acid induced cell death and morphological changes through cell-cycle arrest and induction of apoptotic characteristics, which were observed using DAPI staining and flow cytometric assays. HaCaT cells treated with citric acid exhibited chromatin condensation and apoptotic bodies, suggesting typical morphological apoptotic features. To confirm the apoptotic action of citric acid, cell-cycle analyses were performed. Citric acid also significantly increased the sub-G1 peak of HaCaT cells in a time-dependent manner, indicating the occurrence of apoptosis in HaCaT cells.
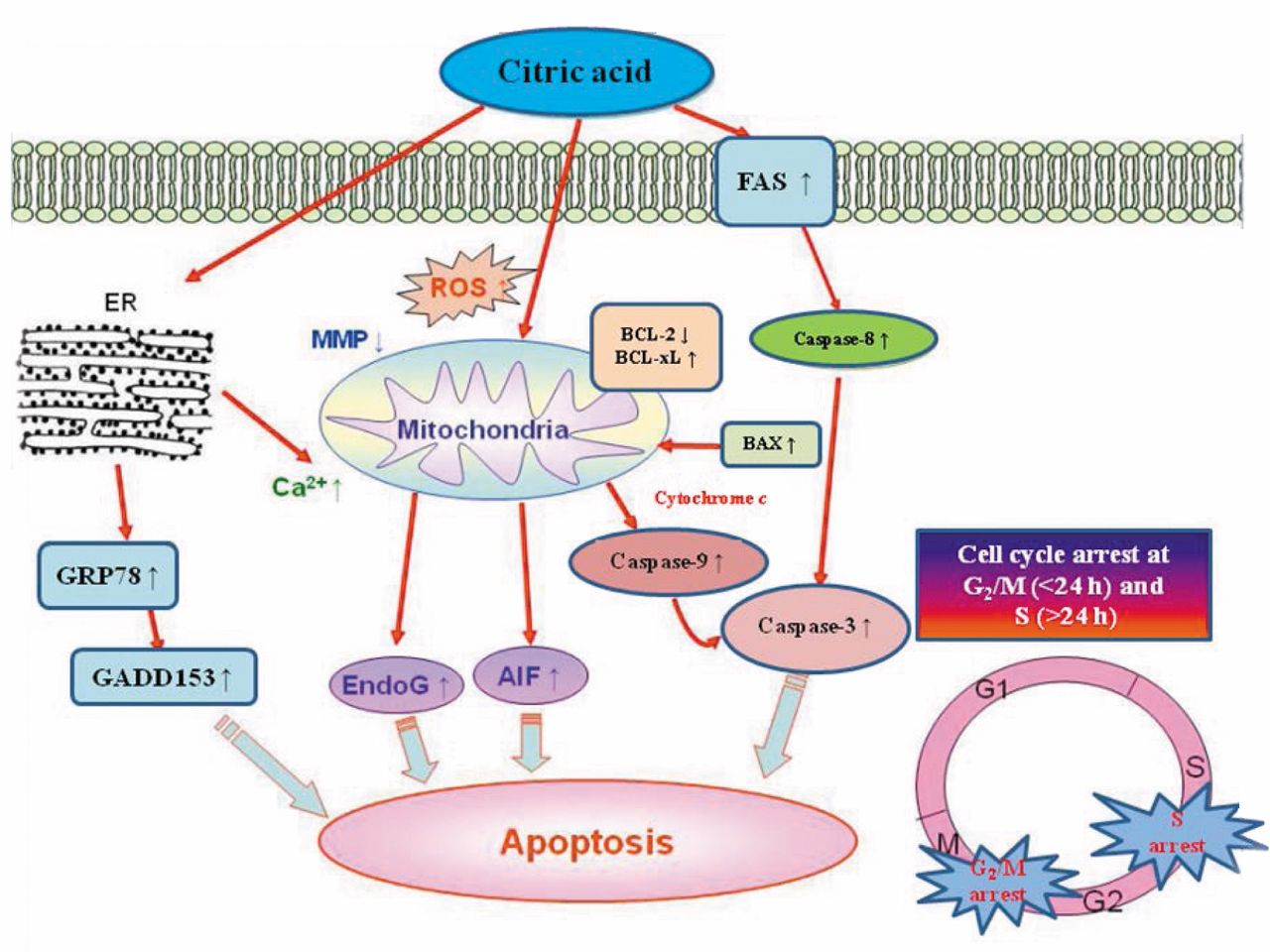
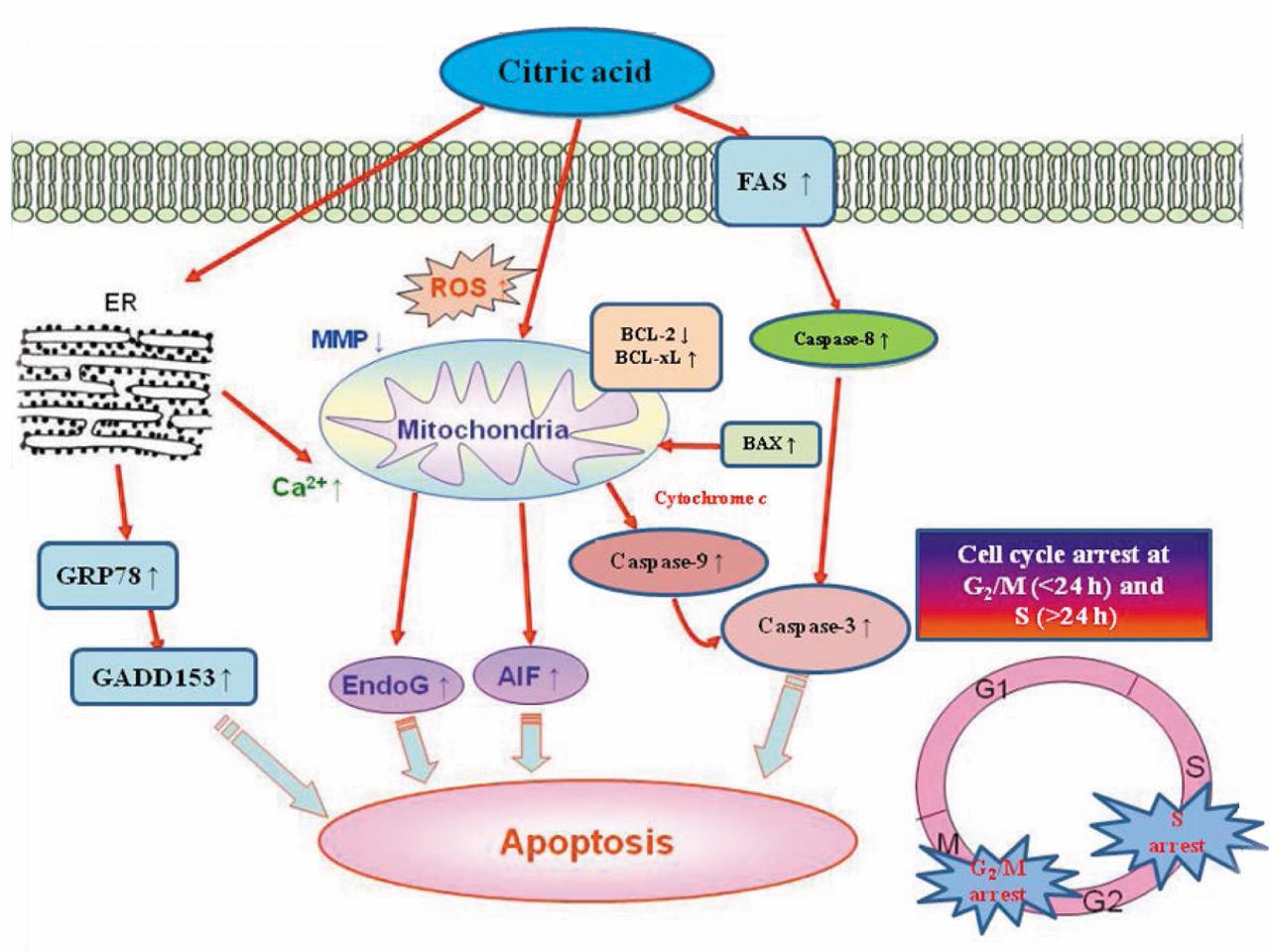
Molecular pathways involved in the effects of citric acid on HaCaT cells. Citric acid inhibits the proliferation of HaCaT cells via induction of cell-cycle arrest and apoptosis.
Apoptosis can be divided into intrinsic and extrinsic pathways through caspase-dependent and -independent pathways (21, 26), based on the activation of caspase family members. It was reported that caspase-8, -9, and -10 are the upstream initiator caspases and caspase-3, -6, and -7 are the downstream effectors caspases (27). Our results shown in Figure 5 indicate that citric acid induced activity of caspase-3, -8 and -9, and Figure 6C indicates that citric acid induced FAS protein expression. These data imply that both intrinsic and extrinsic pathways are involved in citric acid-induced apoptosis. We also used FITC-conjugated immunofluorescent staining before using a confocal laser microscope to examine apoptosis-associated AIF and EndoG proteins. Their translocation from cytoplasm into the nucleus indicated that citric acid promoted the expression of AIF (Figure 7), and EndoG (Figure 7) as well as stimulating intracellular Ca2+ release (Figure 4C), which also indicated that citric acid-induced apoptosis of HaCaT cells occured partly through caspase-independent pathways.
In recent years, numerous studies have demonstrated that AIF induces caspase-independent apoptosis under various conditions (14, 28, 29). Furthermore, Ca2+ has been shown to be involved in AIF cleavage and release from mitochondria (14, 30, 31). In the present study, our findings also indicated that citric acid promoted Ca2+ accumulation (Figure 4C) and AIF release (Figure 6B) from mitochondria, indicating that citric acid induces apoptosis of HaCaT cells via caspase-independent pathways. Results from western blotting (Figure 6C) indicated that citric acid-induced apoptosis might occur through FAS receptor to activate caspase-8, or through mitochondria pathways. The results also showed that citric acid promoted the expression of pro-apoptotic proteins BID, BAX and PARP, and reduced that of anti-apoptotic proteins BCL-2 and BCL-XL (Figure 6A). It is well-known that the ratio of BAX/BCL-2 is related to the level of MMP (14, 32). Our results showed that citric acid reduced the MMP, leading to cytochrome c release and activation of caspase-9 followed by the activation of caspase-3, causing apoptosis.
In summary, after treatment with citric acid, immortalized human epidermal keratinocytes (HaCaT) exhibit apoptotic features of DNA damage, apoptotic bodies, and an increase of sub-G1 cells, resulting from activation of caspase-8, -9, and -3 and the induction of AIF and Endo G release from mitochondria. These findings are summarized in Figure 7.
Conclusion
We demonstrated the apoptotic and cytotoxic effects of citric acid on a human keratinocyte cell line, HaCaT, in vitro. Our investigation highlights to consumers and physicians the potential risks of citric acid and other AHAs which may be contained in cosmetic products.
Acknowledgements
This work was supported by grant NSC97-2314-B-040-026, NSC102-2314-B-303-002 from the National Science Council, Taiwan, Republic of China, the grant TCRD102-42 from Buddhist Tzu Chi General Hospital, and the grant CSH-2011-D-002 from Chung Shan Medical University Hospital.
Footnotes
-
↵* These Authors contributed equally to this work.
- Received August 19, 2013.
- Revision received September 18, 2013.
- Accepted September 19, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}