


Abstract
Background: Several authors demonstrated that normal goblet cells intermingle with dysplastic cells in duodenal adenomas. Goblet cells in the normal duodenum were found to cross-react with the neurotransmitter synaptophysin, a synaptic glycoprotein present in neuroendocrine cells and in virtually all neurons of the central nervous system. In this work, we investigated whether goblet cells in duodenal adenomas express synaptophysin. Materials and Methods: Sections from 29 duodenal adenomas were immunostained for synaptophysin. Results: All intercalated normal goblet cells in duodenal adenomas expressed synaptophysin. The mean percentage of synaptophysin-expressing goblet cells in the basal third of the adenomas was 49%, in the middle third 38% and in the superficial third 5% (basal and middle thirds vs. superficial third, p<0.05). Synaptophysin-positive goblet cells were also stained with alcian blue (pH 2.5) and MUC2, that stain sialomucins. In the normal ileum, only occasional goblet cells were stained (faintly) with synaptophysin. On the other hand, all goblet cells in the normal cecum were synaptophysin negative. Conclusion: Normal goblet cells found in duodenal adenomas express synaptophysin. Consequently, duodenal adenomas with synaptophysin-expressing goblet cells should also be stained with alcian blue (pH 2.5) or with MUC2 to confirm that these are native normal goblet cells, thereby rejecting the possibility of a composite adenoma-microcarcinoid in the duodenum.
Duodenal adenomas are mucosal foci of mutated, dysplastic cells (1) displaying proliferative, biochemical and molecular aberrations (2). In one study comprising of 278 patients with duodenal adenomas, 60% had familial adenomatous polyposis (FAP) and the remaining 40% had sporadic adenomas (3). The cumulative incidence of duodenal adenomatosis in patients with FAP at the age of 70 years is 90% (4) and the estimated risk of duodenal cancer is between 100 and 330 times that for the general population. The lifetime risk of developing duodenal cancer in the remaining adenomatous area is approximately 4% (4).
In patients with FAP the malignant transformation of colorectal adenoma usually ensues rapidly (early in life), while the transformation of duodenal adenoma evolves more slowly (later in life) (5, 6). The causes for this age-related difference in malignant transformation of adenomas located in different organs, in the same patient remain elusive.
It is generally accepted that duodenal adenomas evolve following replacement of normal structures by dysplastic cells (1-7). However, not all cells in duodenal adenomas are dysplastic; Odze et al. (8) found goblet cell differentiation in as many as 18% of the cells in duodenal adenomas.
In a previous study of the normal duodenal mucosa, the neurotransmitter synaptophysin, which specifically stains neurons and synapses in the lamina propia mucosa and submucosa, was also found to stain normal goblet cells and Paneth cells, but not enterocytes (9). Despite the apparent cross-reaction of this immunostain, its distinct expression prompted the evaluation of synaptophysin as a tool for highlighting goblet cells in a cohort of duodenal adenomas.
Materials and Methods
Endoscopic biopsies from 44 patients were investigated: 29 had duodenal adenomas (20 had FAP and 9 sporadic adenomas) and 15 had histologically normal duodenal mucosa.
Sections were cut at 6 μm and stained with hematoxylin and eosin (H&E) and with anti-human synaptophysin antiserum (Leica Microsystems, Wetzlar, Germany). The antibody is ready-to-use, implying that the producer has optimized the dilution. The preparations were incubated for 30 min on a Leica Bond XT instrument (Leica Microsystems) and scrutinized under a conventional microscope using ×4 and ×10 objectives.
Five out of the 29 duodenal adenomas were also stained with chromogranin A, gastrin, glucagon, somatostatin, serotonin and neuron-specific enolase.
Adenomas were classified according to their histological configuration into tubular or villous and according to cellular severity into low-grade or high-grade dysplasia.
With the aid of an ocular scale, the thickness of the normal duodenal mucosa and of the adenoma was divided into a lower third, a middle third and an upper third.
Quantification of goblet cells. To quantify synaptophysin-labeled goblet cells, the following procedure was performed: a representative area containing a priori the highest number of synaptophysin-stained goblet cells in three different high power fields (HPF) in the normal mucosa and in adenomas were photographed at high power (×40). From the image (width 8 cm, resolution 300 pixels/inch) in Adobe Photoshop CS3 Extended, version 10.0.1) an A4 page (scale of 300%) was printed. On the three printed pages per case, the number of synaptophysin-stained goblet cells per total number of cells was calculated, both for the normal mucosa and for the adenoma. This method of quantification was considered to be more appropriate than the current applied method to semi-quantify the percentage of immunohistochemically stained cells into <10% cells stained (+); 10-50% cells stained (++), and >50% cells stained (+++) cells (10).
Statistical analysis. The non-parametric Mann–Whitney test was used. Statistical significance was set at p<0.05.
Results
Histological evaluation. Out of 29 duodenal adenomas, 26 were tubular and the remaining three, villous. Twenty-eight exhibited low-grade dysplasia and the remaining one, high-grade dysplasia.
Immunostains showed that all normal goblet cells in the duodenum expressed synaptophysin.
Qualitative evaluation. Normal duodenal mucosa: Synaptophysin-labeled goblet cells were rounded and ballooned; they were found intercalated amid enterocytes (Figure 1). The mucin-producing glands in the submucosa (Brunner's glands) were synaptophysin negative.
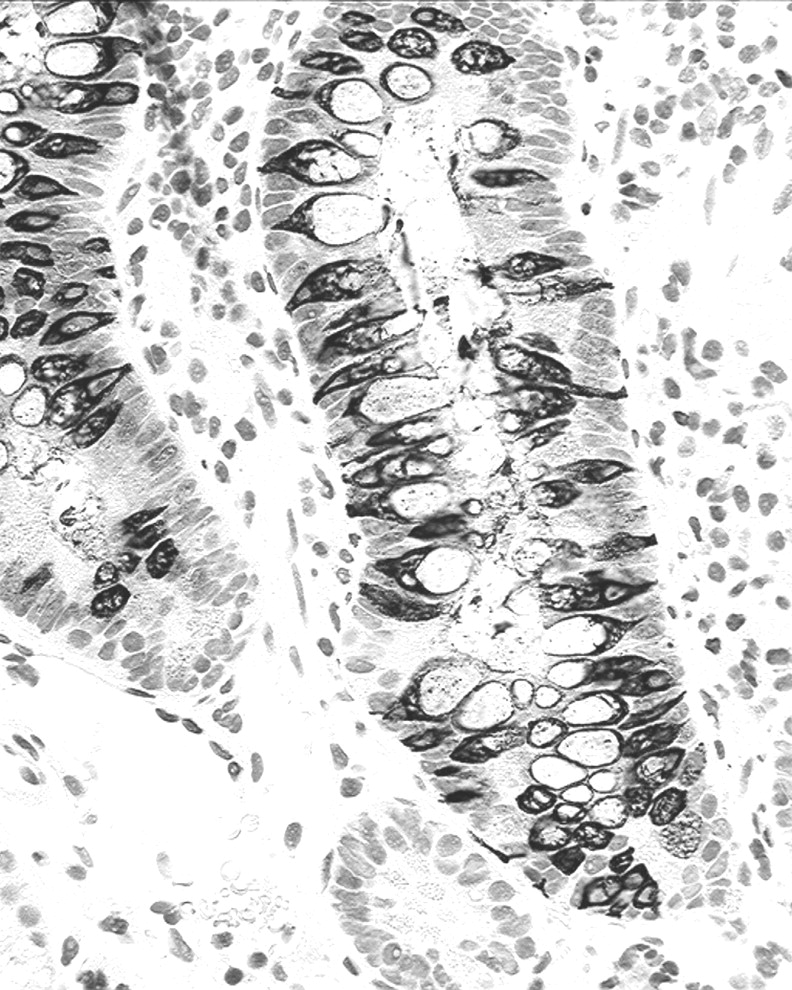
Duodenal adenomas: Synaptophysin-stained goblet cells were often elongated, bipolar, spindle-shaped, their base being attached to the basement membrane; they were found intercalated between dysplastic cells (Figure 2). Occasional rounded ballooned goblet cells were also seen.
Topographic distribution of synaptophysin-expressing cells. Normal mucosa: Synaptophysin stained goblet cells were recorded in the lower, in the middle third and to a lesser extent, in the upper third of the crypt-villous units.
Duodenal adenomas: Synaptophysin-stained goblet cells were found in the basal third in 29 adenomas, in the middle third in 26 adenomas (Figure 2), and in the upper third in 8 adenomas.
No essential quantitative differences were found between nine unselected FAP adenomas and the nine sporadic adenomas.
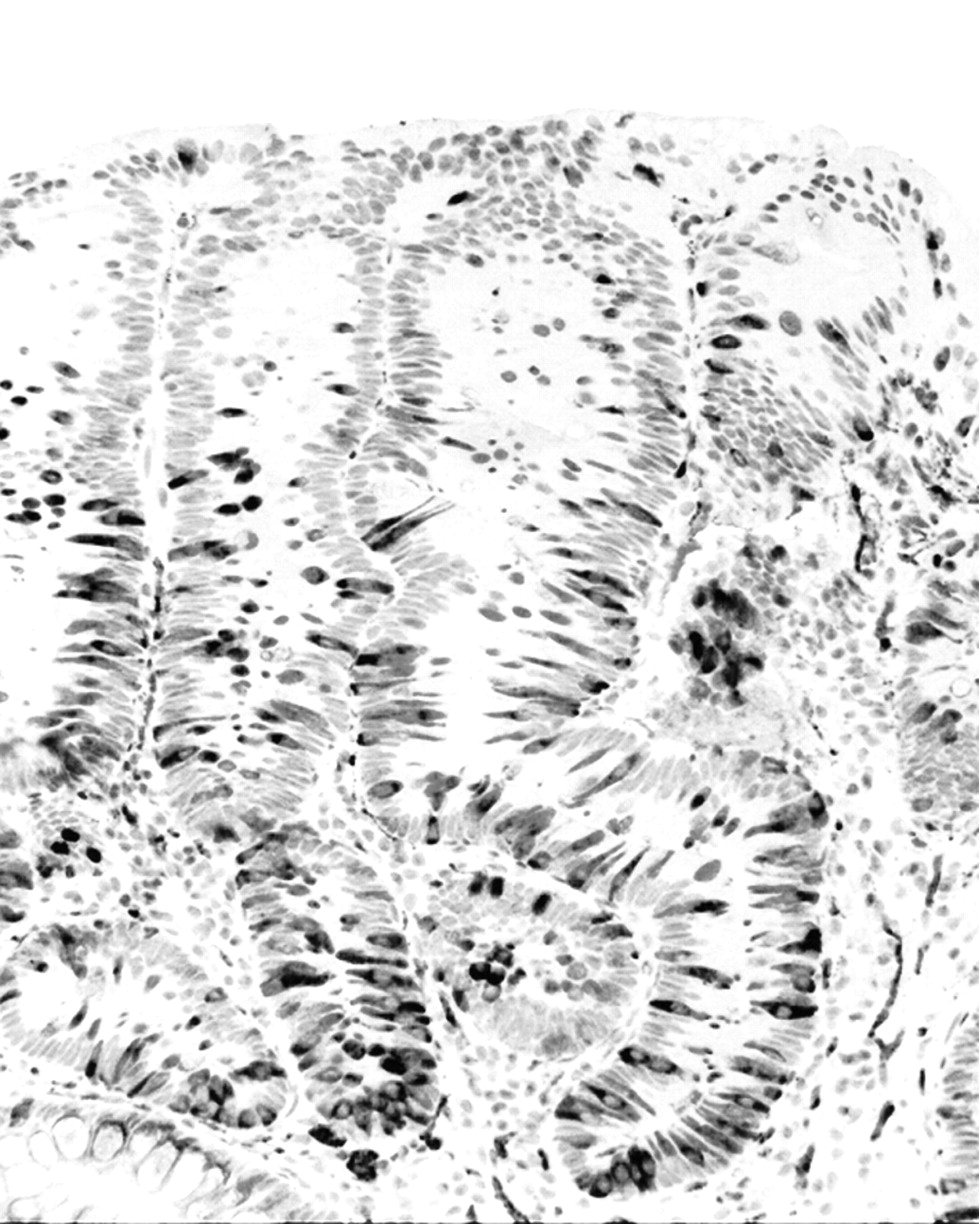
The absence of synaptophysin-expressing cells in the remaining adenomatous areas concurred with the presence of tightly packed adenomatous cells. The deformed, synaptophysin-positive goblet cells were also stained with alcian blue (pH 2.5) stain and/or MUC2 (Figure 3).
Quantitative assessment. Normal duodenal mucosa: Synaptophysin-stained goblet cells were recorded in the lower and middle thirds, and to a lesser extent, in the upper third of the crypt-villous units. The mean percentage of synaptophysin-expressing goblet cells was 10% (range 6%-18%).
Duodenal adenomas: The mean percentage of synaptophysin-stained goblet cells in the basal third in 29 adenomas was 49% (range 29%-89%), in the middle third, it was 38% (range 20%-68%), and in the superficial third, 5% (range 1%-10%). The difference between synaptophysin-labelled goblet cells in the basal/middle thirds and the superficial third was significant (p<0.05). Unfortunately, the number of villous adenomas was too small to permit a statistical analysis.
Discussion
All adenomas analyzed in this survey, contained among dysplastic (adenomatous) cells, intermingled with normal goblet cells. Similarly, all ten duodenal adenomas analyzed by Odze et al. (8) contained interspersed goblet cells. Hence, if all duodenal adenomas contain goblet cells, what is the function of the latter cell phenotype? Does the mucin present in goblet cells protect dysplastic cells from the disparate pH in the microenvironment conveyed by the three main gastrointestinal secretions: gastric, biliary and pancreatic? These secretions are in fact harmful for normal naked epithelial cells without mucous protection. Hence, it is not totally inconceivable that mucins in goblet cells might be required to protect dysplastic cells from the disparate pH in the duodenal microenvironment, thus securing continuous replication of dysplastic cells. The faint synaptophysin expression in occasional goblet cells in the normal ileum and the lack of expression in the normal cecum (9), suggest that the mucin contained in synaptophysin-positive cells was sialomucin. This possibility was endorsed by staining duodenal adenomas with alcian blue (pH 2.5) and MUC2 stains.
Quantitative results indicate that 49% of the dysplastic glands located in the basal third and 38% in middle third of duodenal adenomas, contained intercalated synaptophysin-positive goblet cells. In the superficial third, only 5% of the adenomas contained synaptophysin-positive goblet cells. The absence of synaptophysin-expressing cells in the remaining adenomatous areas concurred with the presence of tightly packed dysplastic cells, suggesting that these areas with dysplastic cells-only might mirror a biological step towards a more self-governing dysplastic cell growth, without the chaperon protection of participating goblet cells.
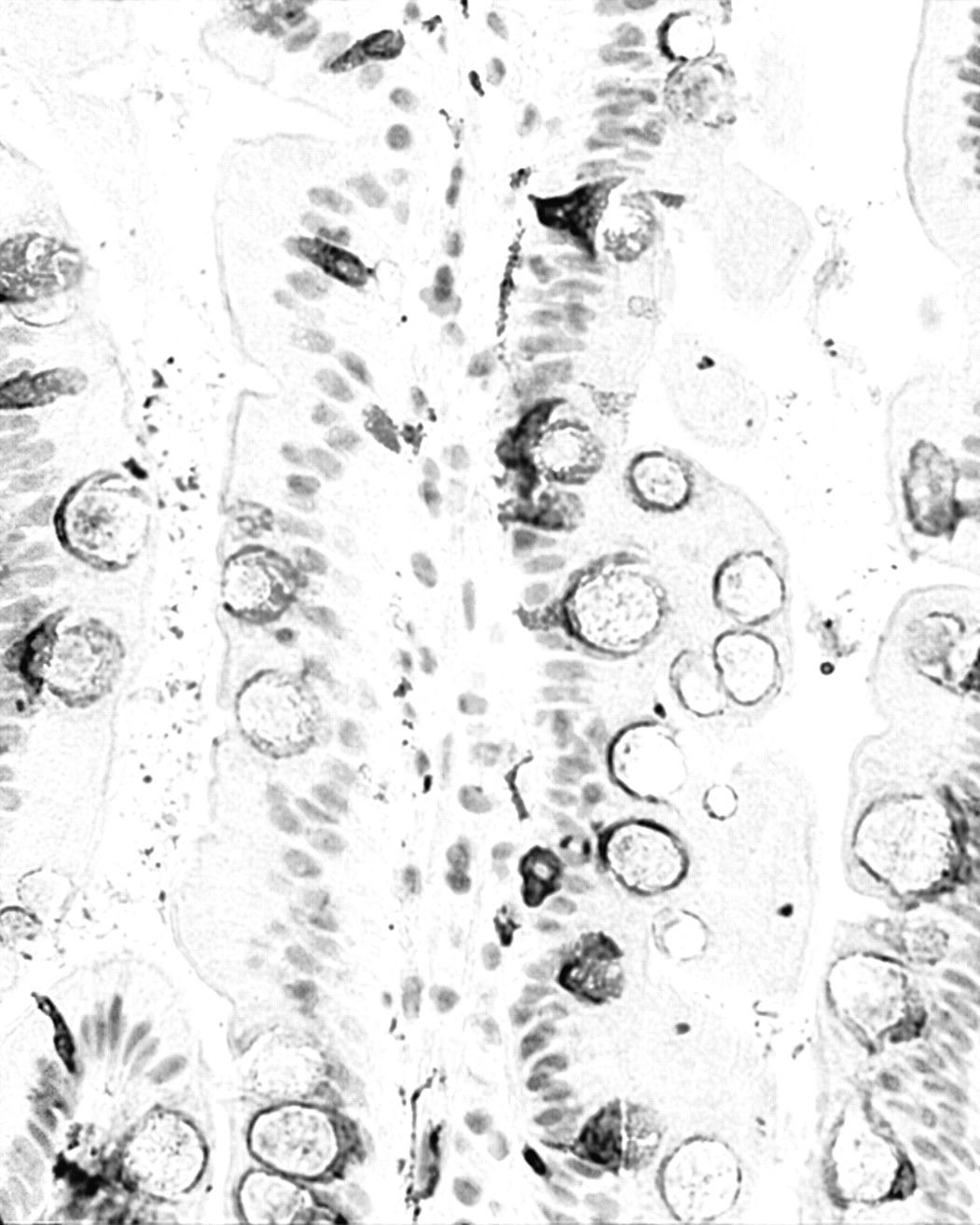
Normal duodenal mucosa highlighting synaptophysin-expressing goblet cells (synaptophysin immunostain, ×40).
Quantitative results also showed that the number of synaptophysin-expressing goblet cells per HPF was significantly higher in the lower thirds of the majority of dysplastic glands than in the normal duodenal crypts. Since dysplastic cells are rapidly proliferating cells and goblet cells are post-mitotic cells (11), it might be assumed that the number of goblet cells per HPF would proportionally decrease in all adenomatous areas. Surprisingly, the opposite was found in some dysplastic areas. The explanation for this remains unclear. Notwithstanding this, Turner and Odze (12) showed that goblet cells in duodenal adenomas are able to synthesize DNA. Similarly, we found DNA synthesis in goblet cells in duodenal adenomas. These results would argue against the common notion that goblet cells are differentiated, mature, post-mitotic cells without the capacity to proliferate (11), at least in duodenal adenomas. The finding that the ratio of goblet cells to dysplastic cells was higher in adenomas than in the normal counterpart substantiates the possibility that, in addition to DNA synthesis, goblet cells might also divide in adenomas (12). This possibility was explored by examining mitotic figures with the mitosis marker anti-phospho histone 3 (Ser10; Milipore, Temecula, CA, USA), but it was difficult to ascertain whether some of the cells in mitosis were in fact goblet cells.
None of the five duodenal adenomas challenged with other immunohistochemical stains (see Materials and Methods) expressed chromogranin A, gastrin, glucagon, somatostatin, serotonin or neuron-specific enolase, indicating that intermingled goblet cells in these five adenomas did not secrete neuroendocrine products, and yet these cells expressed synaptophysin.

Duodenal adenoma. Note synaptophysin-expressing goblet cells in the lower and middle thirds of the adenoma (synaptophysin immunostain, ×40).
Duodenal adenoma showing MUC2-expressing goblet cells (MUC2 immunostain ×40).
Recently, Lin et al. (12) reported two cases of composite adenoma-microcarcinoid in the duodenum. In one of the two cases staining for chromogranin A was negative, whereas that for synaptophysin, was positive. In the other case, no immunostaining was carried out.
Despite synaptophysin obviously cross-reacting with goblet cells, its strong expression permitted monitoring the abrogation of goblet cells, particularly in the superficial compartments in duodenal adenomas. Snaptophysin-expressing goblet cells were often elongated and bipolar, with the foot attached to the basement membrane, deformed by the juxtaposed proliferating dysplastic cells. These cytological characteristics may strengthen the suspicion of a neuroendocrine component. In light of this, duodenal adenomas exhibiting deformed, synaptophysin-positive cells should also be stained with alcian blue (pH 2.5) and/or MUC2 to confirm that these are indeed native goblet cells that secrete sialomucins, rejecting thereby the possibility of a composite adenoma-microcarcinoid in the duodenum (13).
Acknowledgements
Thanks are due to Dr Daren Buhrkuhl, Department of Pathology, Karolinska University Hospital, for revising the manuscript.
- Received April 5, 2012.
- Revision received May 8, 2012.
- Accepted May 9, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.