


Abstract
Aim: The study of the anticancer effects of destruxin B (DB) is rare and its anticancer mechanism remains unknown. The aim of this study was to test the in vitro and in vivo anticancer effects of DB, on human HT-29 colorectal cancer (CRC). Materials and Methods: DB was isolated and characterized by high pressure liquid chromatography, electrospray ionization mass spectrometry and 1H-nuclear magnetic resonance spectroscopy. (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to assess the effects of DB on HT-29 cells in vitro. The anticancer effects of DB were investigated in a murine xenograft model of human colon cancer. Results: A significant inhibition of cell viability was observed with DB treatment in time- and dose-dependent manners. DB administered subcutaneously daily at 0.6-15 mg/kg was proven to be safe and effective in inhibiting the growth of CRC cells. Expression of Bax, cleaved poly (ADP-ribose) polymerase and active caspase-3 were observed with DB treatment and the increase in tumor volumes of treated groups were significantly (p<0.05) lower than those of the mock-treated group. Conclusion: DB has potential as a new therapeutic agent against human CRC.
Metarhizium is one of the best known genera of entomopathogenic fungi, commonly known as green muscardine fungus. Within this genus, M. anisopliae has been used for insect control in many countries including the USA, Brazil, Australia and the Philippines (1), and the most abundant metabolites of M. anisopliae are destruxins (2). These metabolites are usually secreted into the culture medium during growth. Destruxins are a class of cyclodepsipeptides composed of an alpha-hydroxy acid and five amino acid residues including proline, isoleucine, methylvaline, methyl-alanine, and beta-alanine (3). In recent years, the optimal conditions for culture and analysis of destruxins has been established (4). To date, there are 38 known destruxins or destruxin analogs chemically divided into five basic groups labeled A to E (5). Previous studies have shown that destruxins exhibited strong biological effects; in particular, destruxin E disturbs macromolecular syntheses (DNA, RNA and protein synthesis) (6), produces antiviral effects (7) and modifies the DNA content of leukemia cells (8, 9) in vitro. However, the study of anticancer effects of destruxins, especially destruxin B (DB), is rare.
Colorectal cancer (CRC) is one of the most common malignant diseases worldwide. According to the 2010 statistics of causes of death in Taiwan (10), CRC is the third leading cause of cancer deaths in Taiwan. Although curative resection remains the primary treatment for this disease, recent advances in the field of CRC chemotherapy have resulted in increased survival and improved quality of life in affected patients (11). Some studies (12-15) have noted that non-steroidal anti-inflammatory drugs, such as aspirin and fluorouracil (5-FU), can improve the prognosis of patients with CRC and serve as effective cancer chemopreventive or chemotherapeutic agents. However, the potential hazards of adverse effects associated with long-term use of such agents should be carefully considered.
In order to investigate the anticancer effects of DB, human HT-29 CRC cells were utilized. This is the first study examining the effects of DB as an anti-CRC agent in a murine xenograft model of human CRC. DB was explored potentially as a new therapeutic agent against human CRC.
Materials and Methods
Production of destruxins. A culture of M. anisopliae F061 var. anisopliae kindly provided by Dr. Suey-Sheng Kao, Taiwan Agricultural Chemicals and Toxic Research Institute (Taichung, Taiwan, ROC), was used in this study. The culture method used was as previously described (4). Briefly, the spore suspension culture at −80°C was thawed at room temperature and inoculated into a 500 ml Erlenmeyer flask with a baffle containing 200 ml of 3% (w/v) Czapek-Dox (CD) broth (Becton, Dickinson and Company, Spark, MD, USA) and 0.75% bacto-peptone (Becton, Dickinson and Company) as seed culture. The flask was cultivated in an incubator at 200 rpm, 28°C for 4 days. For the stirred-tank cultivation, the inoculum (10% of the working volume) was transferred from the flask of the 4-day old seed culture to the reactor, which contained 3l of the desired medium. Cultivations were conducted in a 5l stirred tank reactor at 28°C with the aeration rate regulated at 0.3 volume air/volume liquid/min (vvm). The culture medium (pH 9.0) was maintained by automatic addition of 2 N NaOH or 1 N HCl at an agitation rate of 150 rpm. After 14 days, the fermentation broth was harvested and purified as described below.
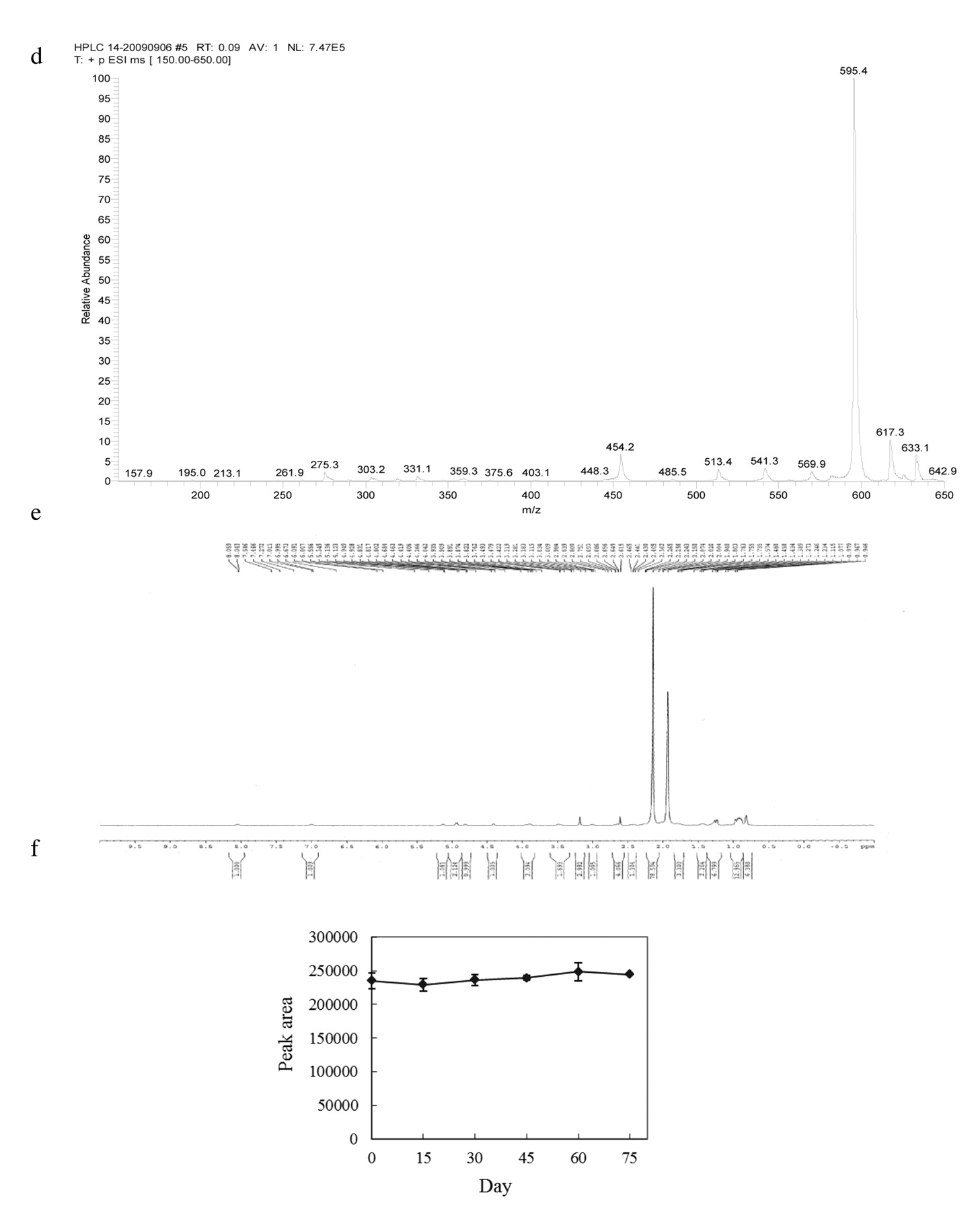
Purification of DB. The optimal conditions for destruxin purification were established according to a series of experiments of our previous studies (4, 16-19). The culture medium was harvested after incubation for 14 days and centrifuged at 700 ×g for 20 min. The supernatant was adjusted to pH 4.0 by 1 N HCl then extracted with ethyl acetate (sample: EA=5:2, v/v), and the organic phase was evaporated with a rotary vacuum evaporator (model N-1, Eyela, Tokyo, Japan) at 45°C. The concentrate was diluted with twice the volume of acetonitrile and filtered through a 0.22 μm chromatodisc unit before high pressure liquid chromatography (HPLC) analysis. The sample (800 μl) was injected into a preparative column (Cosmosil 15 C18-AR-II column, 28×250 mm, 15 μm). The eluent from the column was monitored at 215 nm with a L-7100 pump and a L-7400 UV detector (Hitachi, Tokyo, Japan). The mobile phase was: 80% methanol/H2O at a flow rate of 10 ml/min. Fractionated samples were characterized by analytic HPLC, electrospray ionization mass spectrometry (ESI-MASS) and 1H-nuclear magnetic resonance (NMR) spectroscopy. The chemical structure of DB (C30H51N5O7, molecular weight 593) is shown in Figure 1a.


a: Chemical structure of DB. b: Preparative reversed-phase HPLC chromatographic plot of the crude extract sample. Sample was injected into a preparative reversed-phase C18 HPLC column. The mobile phase was 80% MeOH/H2O, with a flow-rate of 10 ml/min, and the absorbance was recorded at 215 nm. The bar indicates where a single destruxin was collected from eluted fractions. c: HPLC analysis of purified DB from the preparative HPLC. The mobile phase was 50% acetonitrile/H2O, with a flow-rate of 1 ml/min, and the absorbance was recorded at 215 nm. The red line represents purified DB collected from preparative HPLC. The black line represents the DB standard. The retention time of DB is 5 min. d: Positive ESI-MASS spectograph of DB. e: 1H-NMR spectrum (500 MHz) of DB in acentonitrile. f: HPLC analysis of DB content after storage at 4°C for 75 days.
Cell line and cell culture. Human colon adenocarcinoma cell line HT-29 was routinely grown in RPMI-1640 medium supplemented with 10% heat-activated fetal bovine serum (FBS), 2 g/l sodium bicarbonate (SIGMA, St. Louis, MO, USA), and 1% penicillin-streptomycin antibiotics at 37°C in a humidified atmosphere with 5% CO2.
Cell viability assay. The effect of DB on cell viability was assessed by using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay in 4 to 6 replicates as previously described (20-25). HT-29 cells were grown in 5% FBS-supplemented culture medium in 96-well plates overnight. After 24 h, cells were incubated in medium containing different concentrations (0 to 25 μM) of DB for 3 days and then washed with phosphate-buffered saline. The cell viability was determined according to the amount of formazan crystals formed 3 h after MTT addition. The crystals were dissolved in dimethyl sulfoxide (DMSO) and the optical density was measured at 570 nm by a multi-well ELISA reader (BIO-RAD, Hercules, CA, USA).
Animal model of tumor xenografts. Athymic female nude mice (BALB/cAnN.Cg-Foxn1nu/CrlNarl), approximately 4-5 weeks old on arrival, were purchased from the National Laboratory Animal Center (Taipei, Taiwan, ROC) and maintained under specific pathogen-free conditions. The institutional guidelines for animal experiments (Animal Centre, Chung Shan Medical University) were followed for animal care and use (approval No. 807). HT-29 cells growing exponentially were harvested by brief incubation with 0.05% trypsin-EDTA solution. Before drug treatment, a xenograft was initiated by subcutaneous (s.c.) injection of 1×107 cells into the right flank of nude mice. Transplanted tumors were allowed to grow to a volume of more than 100 mm3. The mice were then randomly divided into three classes. Each class had six experimental groups, with 6 mice for each group. The mice receiving 14 days' treatment were designated as class I (mock-1, vehicle-1, 5-FU-1, high-1, medium-1, low-1); those receiving 28 days' treatment as class II (mock-2, vehicle-2, 5-FU-2, high-2, medium-2, low-2); and those receiving 42 days' treatment as class III (mock-3, vehicle-3, 5-FU-3, high-3, medium-3, low-3), respectively. Different doses (low, 0.6 mg/kg/day; medium, 3 mg/kg/day; and high, 15 mg/kg/day) of DB, 5-fluorouracil (5-FU, 5mg/kg, Pharmachemie B.V., GA, Harrlem, Netherlands) with DMSO vehicle as the positive control, phosphate-buffered saline mock, and DMSO vehicle was injected into the tumor daily. The tumor volume was recorded using Vevo 770™ high-resolution micro-ultrasound imaging system (VisualSonics Inc., Canada). Body weights were measured at 7-day intervals. Splenic weights of mice of each group were measured after sacrifice.
Evaluation of caspase-3 activity. Caspase colorimetric assay kit was used for the determination of caspase-3 activity as previously described (22, 24, 26). Briefly, tissue was resuspended in 300 μl of cell lysis buffer (Clontech Laboratories Inc., Mountain View, CA, USA) and incubated on ice for 30 min then centrifuged for 30 min in a microcentrifuge (12,000 ×g). The supernatant (50 μl) was mixed with 50 μl of 2× reaction buffer/dithiothreitol mixture and 1 μl of the caspase-3 inhibitor DEVD-fmk (1 mmol/l) and incubated on ice for 30 min. Five microliters of 1 mmol/l caspase-3 substrate DEVD-pNA was added and the mixture was incubated at 37°C for 1 h. After incubation, each sample was measured at 405 nm in a ELISA microtiter plate reader. The activity of caspase-3 in tissue samples was calculated using a standard curve and expressed as enzyme units.
Sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) and western blotting. SDS-PAGE and western blotting were performed as previously described (24, 25). Total proteins were solubilized and extracted from tissue with 300 μl lysis buffer. The lysates were used to estimate the protein content with Bradford protein assay. Equal amounts of protein (50 μg) from each sample were then subjected to electrophoresis on a 12% (v/v) SDS-polyacrylamide gel. After electrophoresis, proteins were electroblotted to a PolyScreen® PVDF hybridization transfer membrane (PerkinElmer, Boston, MA, USA). The membrane was blocked at room temperature with blast blocking buffer [1% blast blocking reagent (PerkinElmer, Boston, MA, USA) in Tris buffered saline (TBS) containing 0.05% (v/v) Tween (TBS-T)]. Then the membrane was washed three times with TBS-T and incubated overnight at 4°C with the primary antibody, mouse monoclonal anti-caspases-3 (1:1000, v/v), or rabbit monoclonal antibodies to poly (ADP-ribose) polymerase (PARP) (1:4000, v/v), Bcl-2-associated X protein (Bax) (1:8000, v/v) and β-actin (1:2000, v/v), followed by 1 h incubation with a 1:10000 (v/v) dilution of the appropriate horseradish peroxidase (HRP)-conjugated secondary antibody. Primary antibodies (β-actin, PARP, Bax) were purchased from Cell Signaling Technology Inc. (Danvers, MA, USA), whereas secondary antibodies (anti-rabbit IgG and anti-mouse IgG HRP-labeled) were purchased from PerkinElmer (Boston, MA, USA). After incubation, the membrane was washed with TBS-T three times, and the antigen-antibody complexes were visualized by the enhanced chemiluminescence system (PerkinElmer). The relative protein expression with the control, defined as 100%, were measured by Image J software (National Institutes of Health, USA).
Histopathology and immunohistochemistry. The tumor tissues were dehydrated, embedded in paraffin and cut into 4-μm-thick sections. For histopathology, the sections were rehydrated and stained with hematoxylin and eosin according to the standard protocol. The stained sections were analyzed for tumor grade by a pathologist who was unaware of the treatments. For immunohistochemical analysis, only non-invasive areas were selected for the evaluation of protein markers. Sections were treated with primary antibody against caspase-3 (1:250 diluted; Santa Cruz Biotechnology). Images were collected randomly at ×400 using a Zeiss AxioCam HRc camera fitted to a Zeiss Axioskope 2 Plus microscope (Carl Zeiss USA, Thornwood, NY, USA).
Statistical analysis. Statistical comparisons between the experimental groups were performed by one-way analysis of variance (ANOVA) with post-hoc Fisher's t-tests using Origin 8.0 professional statistical software (OriginLab, Northampton, MA, USA). All biological experiments were repeated at no less than three separate times. Each value represents the mean±standard deviation (SD), p<0.05 was considered significant.
Results
DB isolation, purification, and degradation analyses. To purify DB, the organic layer as crude extract from fermentation broth of M. anisopliae F061 var. anisopliae was concentrated by rotary vacuum evaporator and injected into a preparative HPLC column. The resulting chromatogram is shown in Figure 1b, the retention time of DB is 17-18 mins. The fraction of DB was collected and freeze-dried. Overall, 10 ml of crude extract was extracted from 3 l working volumes of fermentation broth. The yield was 260.8 mg/l. For confirmation of the purity, purified DB was dissolved in acetonitrile and injected into the preparative column. The result is shown in Figure 1c. The retention time of the sample purified from the preparative HPLC column is the same as the standard.
The identities of these purified destruxins were established on the basis of ESI-MS and 1H-NMR spectrometry. Figure 1d shows the normal positive ESI mass spectrum of preparative HPLC fractions of DB (molecular mass is about 593.7) produced from cultivation of M. anisopliae. There is a very strong peak at m/z 595.4, which corresponds to DB, as previously reported (16). Figure 1e represents the 1H-NMR spectrometry for purified DB: 1H-NMR (500 MHz, C2D3N) δ 5.136, 4.949, 4.928, 4.802, 4.684, 4.042, 3.935, 3.422, 3.034, 2.751, 2.751, 2.441, 2.362, 2.265, 2.150, 1.735, 1.458, 1.309, 1.234, 0.946. The 1H NMR result is identical to the one, previously reported by our group (16).
The DB degradation assay was performed by dissolving 1 mg DB into 1 ml acetonitrile which was then stored at 4°C. The analytical HPLC column was used to detect the DB content for 75 days. The peak area correlating to the content of DB suggested that no significant difference during the storage period was detected (Figure 1f). This storage condition was thus used through this study.
Cell viability assay. MTT assay was used to assess the effects of DB on HT-29 human colorectal cancer cells in vitro. 1.25, 2.50, 5.00, 10.00 and 20.00 μM DB inhibited cell viability by 28.7, 43.9, 73.5, 84.3 and 90.9%, respectively after 48 h treatment, compared with controls that were treated only with vehicle (indicated as 0 μM) (Figure 2). The results demonstrated that a significant inhibition in cell viability was observed upon treatments with various concentrations of DB in time- and dose-dependent manners. The half maximal inhibitory concentration (IC50) of DB at 24 h, 48 h, and 72 treatment on HT-29 cells was determined as 14.97, 2.00, and 0.67 μM, respectively.
In vivo cancer therapeutic effects of DB on body weight, spleen weight, and tumor volume. Tumor volume greater than 100 mm3 was successfully induced in 108 out of 120 BALB/cAnN.Cg-Foxn1nu/CrlNarl female nude mice. These 108 mice were then randomly divided into three classes as described in Materials and Methods. No mouse died during the experiment. There was no obvious metastasis, peptic ulcer, haemorhage, or pathological change of the liver and kidney observed for any of the DB-treated mice (Figure 3). To assess the effects of treatments on toxicity, body weight and food, water consumption were also monitored throughout the study. DB at doses of 0.6, 3, or 15 mg/kg daily was not found to be associated with any apparent toxicity in terms of food and fluid consumption (data not shown). The effects of treatment on body weight and splenic weights are shown in Table I. After 6 weeks' treatment, there was no statistically significant difference observed in body weight and splenic weights, although the splenic weights of some groups, including mice of all classes receiving high dose DB and medium DB in class III, were slightly higher than those of the mock groups.

Effects of DB on the cell viability of HT-29 human colon carcinoma cells. The cells were treated with the indicated concentrations for different time periods and cell viability assessed by MTT assay. Each value is expressed as the mean±SD (n=3).
Tumor volume was measured by Vevo 770™. The tumor volume of individual mice in each treatment group was normalized with respect to the initial tumor volume prior to treatment. Figure 4 shows the normalized tumor volume for control and treated groups. As shown in Figure 4, DB attenuated the tumor growth in time- and dose-dependent manners. Treatment with DB for 6 weeks at low dose (0.6 mg/kg), medium dose (3 mg/kg), and high dose (15 mg/kg) daily indicated a reduction of 23.9%, 33.4% and 55.8% in mean tumor size compared with the mock group, respectively. The dose of 3 mg/kg of DB modestly reduced the tumor size and was comparable with the effect of the clinical drug 5-FU (5 mg/kg: 28.5%).
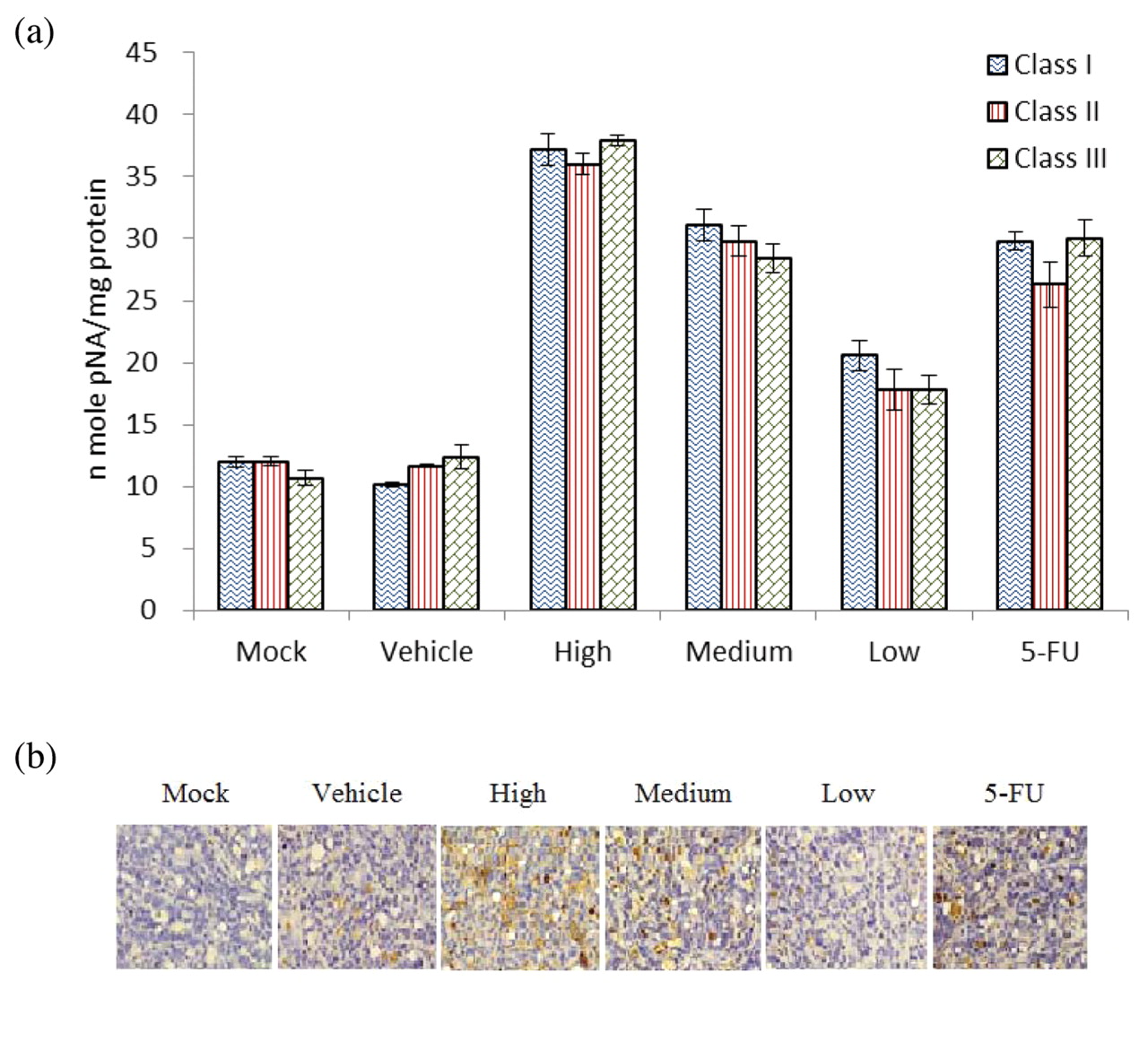
Caspase-3 activity. Caspase-3 activity was measured as being representative of ongoing levels of apoptosis in tumor samples since the proapoptotic enzyme caspase-3 is activated at a point of convergence for induction of the intrinsic and extrinsic apoptosis pathways. To assess the effects of DB treatment on molecular targets in vivo, caspase colorimetric assay was used to demonstrate the caspase-3 activity of each group (Figure 5a). All the DB-treated groups demonstrated significantly (p<0.05) higher caspase-3 activity than the mock and vehicle control groups in classes I-III. As the caspase-3 activity was not significantly different between the same treated groups in classes I-III, the tumor samples of class I were used for further apoptosis assays. Immunohistochemical analysis revealed that the HT29 xenograft, treated with DB showed a marked increase in caspase-3 staining (Figure 5b). These data suggest that apoptotic cell death was significantly induced with DB treatment in the xenograft HT29 cells in a dose-dependent manner.
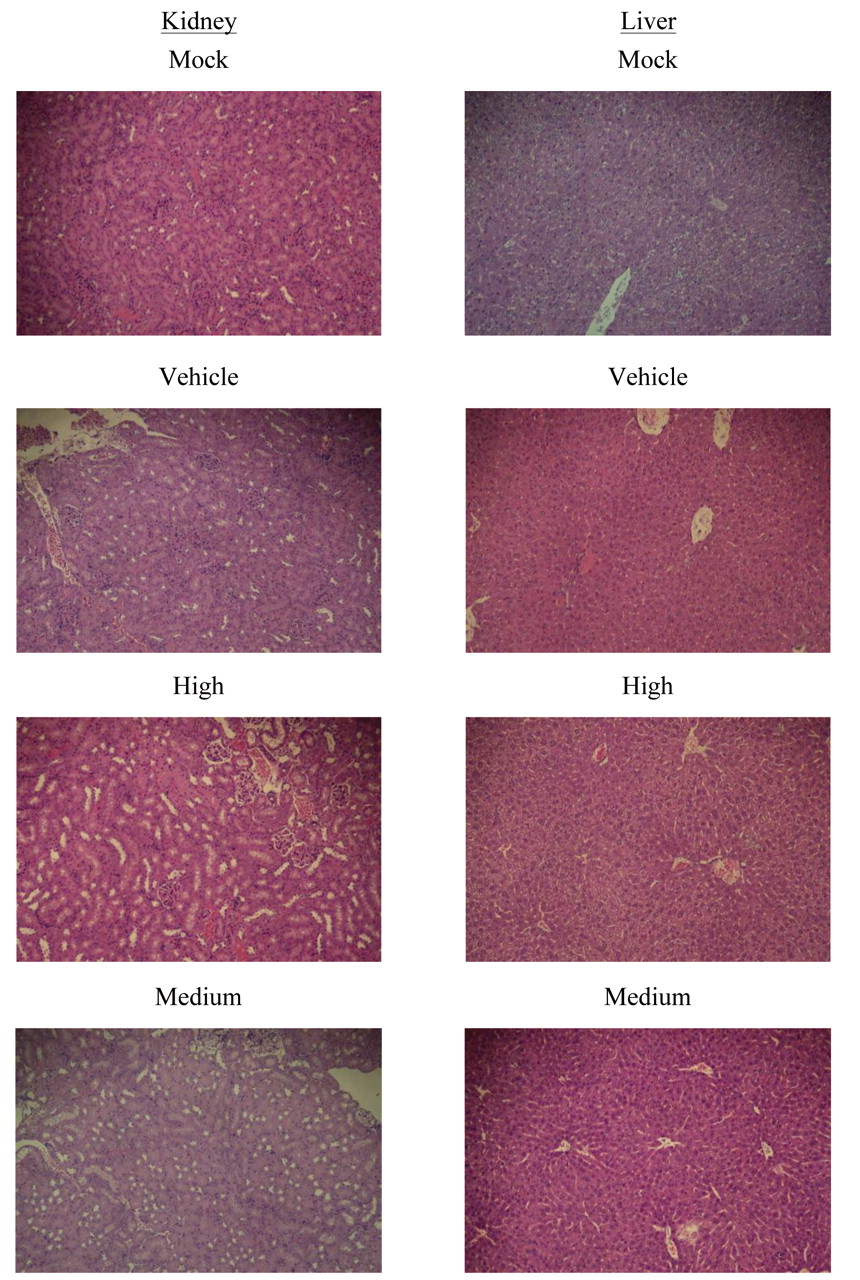

The pathological examination of kidney and liver for Class III (original magnification, ×100). 5-FU: 5-fluorouracil; Low: 0.6 mg/kg/day; Medium: 3 mg/kg/day; High: 15 mg/kg/day, Mock: phosphate-buffered saline; Vehicle: dimethyl sulfoxide.
Expression of Bax and cleavage of PARP. To verify the in vivo apoptotic effects of DB, the tumor samples from class I were used for the analysis. Equal amounts of tumor tissue proteins were subjected to SDS-PAGE followed by western blotting analysis for the expression of Bax and cleavage of PARP with β-actin expression as the internal control. Western blotting analyses showed that DB treatment increases Bax protein expression and induces the activation of PARP (Figure 6a). Significant PARP cleavage was also observed in the 5-FU-treated groups. Tumor tissues from DB-treated mice exhibited higher Bax and PARP protein expressions than that of the mock group (Figure 6b). These data suggest that in vivo, Bax-mediated apoptosis of colon cancer tissue can be induced by DB.
Discussion
In the present study, we provide the first in vitro and in vivo evidence for the effects of DB on human HT-29 CRC. The in vitro results demonstrated that a significant inhibition in cell viability was observed upon treatments with different concentrations of DB in time- and dose-dependent manners. Drug-related death, body weight loss, splenomegaly and pathological examination are used as parameters for the evaluation of toxicity in vivo. Toxicity of DB at the doses of 0.6 to 15 mg/kg/day, administered up to six weeks was not observed, as evidenced by no significant changes in body weight, splenomegaly (Table I), and pathological examination (Supplementary Figure 1).

Normalized tumor volume of HT-29 xenografts. Values shown are from baseline (day 0) and at weeks 2, 4, and 6 post-treatment initiation. 5-FU: 5-fluorouracil; Low: 0.6 mg/kg/day; Medium: 3 mg/kg/day; High: 15 mg/kg/day, Mock: phosphate-buffered saline; Vehicle: dimethyl sulfoxide.

Caspase-3 activity of HT-29 xenograft treated with DB. a: Caspase-3 activity was determined by colorimetric assay kit. Class I, class II, and class III: mice received 14, 28 and 42 days' treatment, respectively. b: Xenograft specimens removed from controls and mice treated with DB were immunohistochemically analyzed for the expression of caspase-3 (magnification, ×200). 5-FU: 5-fluorouracil; Low: 0.6 mg/kg/day; Medium: 3 mg/kg/day; High: 15 mg/kg/day, Mock: phosphate-buffered saline; Vehicle: dimethyl sulfoxide.
The spleen is one of the key organs in the immune system. The spleen can be enlarged in response to medical conditions such as cancer, or to drug administration. Although splenomegaly was not observed in this study, a previous report has suggested that DB, as a hexacyclodepsipeptide, showed insecticidal activities by weakening the immune defense of insect hosts (27). Moreover, an undecapeptide, cyclosporine, from the mosquito pathogenic fungus Tolypocladium inflatum, reduces interleukin-2 expression in mammalian cells by inhibiting calcineurin and has been used recently in human therapies such as organ transplantation for suppression of the host immune response (28). Whether DB modulates the immune response deserves further investigation.
For the use of DB, an appropriate vehicle is needed. The body weight of the vehicle-treated group was also not significantly changed after 6 weeks' treatment. In the present study, DMSO was used as the vehicle and the safety, toxicity, and biocompatibility are herein demonstrated as being acceptable. Recently, a chitosan-egg phosphatidylcholine (chitosan-ePC) implantable formulation has been developed for localized and sustained delivery of paclitaxel (PTX) (29). The formulation is demonstrated to be a safer and better tolerated method for PTX administration, with significant increase in maximum tolerated dose and enhanced antitumor efficacy, suggesting improved therapeutic index with possible clinical implications in the treatment of ovarian tumors (29). Other vehicles including chitosan-ePC may be worthy of testing for better therapeutic effects.
Body weight (a) and splenic weight (b) of human xenograft in nude mice treated with DB or 5-FU.
Caspases are cysteine proteases that play a critical role in the execution of apoptosis. Several chemotherapeutic and chemopreventive agents have been shown to cause apoptotic cell death through activation of caspases (30). In this study, the colorimetric assay and immunohistochemical analysis results showed that caspase-3 is significantly activated with DB treatment (Figure 4). Moreover, the results of western blotting have demonstrated that DB treatment elevated the level of Bax, which promotes cell death and cleaves pro-caspase-3 to active caspse-3, triggering PARP cleavage (Figure 5). Caspase-3 activity was significantly increased with DB treatment in all classes. Furthermore, the increase of caspase-3 and PARP cleavage was also observed in the group treated with the clinical drug 5-FU and this is consistent with a previous study (31). In this study, expression of Bax, cleaved PARP and active caspase-3 were observed with DB treatment (Figure 5) and the tumor volume increases of treated groups were significantly less than those of the mock group (Figure 3). These results suggest that DB reduces tumor cell proliferation through caspase-mediated apoptosis, not only in vitro but also in vivo.
In conclusion, our data indicate that DB administered daily s.c. at 0.6-15 mg/kg was safe and effective in inhibiting the growth of human CRC cells in a cancer xenograft mouse model. In the present study, however, the xenograft tumors treated with DB continued to grow slowly, suggesting that complete inhibition of xenografted HT-29 cells was not achieved by using 15 mg/kg DB treatment alone. Higher doses of DB or in combination with other potent drugs as polytherapy may be needed to achieve complete inhibitory effects.

Effect of DB on apoptosis of HT-29 xenograft tumor. a: The expression of Bax and PARP were analyzed by western blotting, with β-actin expression as the internal control. b: The protein expression relative to the one of the control, defined as 100%, was measured by Image J software (National Institutes of Health, USA) (n=3). 5-FU: 5-fluorouracil; Low: 0.6 mg/kg/day; Medium: 3 mg/kg/day; High: 15 mg/kg/day, Mock: phosphate-buffered saline; Vehicle: dimethyl sulfoxide.
Acknowledgements
The Vevo 770™ real-time high-frequency micro-ultrasound imaging system was utilized at the Instrument Center of Chung Shan Medical University, which is supported by National Science Council, Ministry of Education and Chung Shan Medical University. This study was funded by a grant from the Chung Shan Medical University (98-CCH-CSMU-07) and the National Science Council, Taiwan (NSC91-2113-M-324-008).
Footnotes
-
↵* These authors contributed equally to this study.
- Received December 16, 2011.
- Revision received January 26, 2012.
- Accepted January 30, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}